高原2種草本植物的光合作用和葉綠素熒光參數日動態
2015-12-16 07:44:50高麗楠陳舒慧孫佳媛
四川師范大學學報(自然科學版) 2015年4期
關鍵詞:植物
高麗楠, 張 宏, 陳舒慧, 賀 瓊, 孫佳媛
(1.成都大學旅游文化產業學院,四川成都610106;2.四川師范大學資源生態學研究中心,四川成都610066)
青藏高原屬于氣候變化的敏感區和生態脆弱帶,在全球氣候變化背景下呈現出暖化趨勢[1].青藏高原的沼澤化草甸面積4.9×104km2,占整個青藏高原草地面積的1.96%,是青藏高原分布面積較廣的草地生態系統之一[2].作為青藏高原較為典型的植被類型,高寒沼澤草甸是適應高原隆起和長期低溫環境形成的特殊產物,也是牧民的主要牧草來源.但其生態系統極其脆弱,對人類干擾和由溫室氣體增加引起的全球氣候變化極為敏感,一旦遭受破壞很難在短期內得到恢復,并可能導致草地退化和沙漠化發生,生態環境日趨惡化.在全球變化情景下,高寒沼澤草甸土壤層的碳釋放和植被層的碳吸收對全球變化更為敏感,因而在區域生態系統碳平衡過程中起著極為重要的作用[3].然而,光合作用是植物生長發育的基礎和生產力高低的決定性因素,光合作用過程中所吸收的CO2是陸地生態系統碳循環的入口,并對減緩全球溫室效應具有重要作用[4],同時也是一個對外界環境變化非常敏感的生理過程.因此,深入研究高寒沼澤草甸的光合特性是當前急需開展的工作.近年來,高寒草甸植物光合作用的研究相對較少[5-6],并且尚未見到系統探討青藏高原高寒沼澤草甸光合特性的報道.
川西北高寒沼澤草甸位于青藏高原東緣,是我國主要牧區之一.川西北高寒沼澤草甸不僅是四川省重要的水源地,而且是黃河上游重要的水源涵養區和集水區.目前川西北草地退化面積已達60%,出現草地生產力降低、水土流失嚴重等多種生態系統退化癥狀,生態系統失衡的問題十分突出[7],其中,四川嵩草和木里苔草是川西北高寒沼澤草甸植物群落的優勢種,其良好生長對維持青藏高原的生態平衡起著重要作用,但對其生理生態學方面的研究較少.所以,本文重點討論青藏高原東緣高寒沼澤草甸2種優勢草本的光合作用和葉綠素熒光參數的日動態,以探討高寒環境對植物光合作用的影響及高山植物光合特性,也為該地區植被恢復與重建提供理論依據.
1 材料與方法
1.1 實驗樣地 實驗樣地位于四川省阿壩州紅原縣(101°51'-103°23'E,31°51'-33°19'N,平均海拔3 600 m),地處青藏高原東部邊緣.氣候屬大陸性高原氣候,日溫差大、霜凍期長,年平均氣溫1.1℃,年平均相對濕度60% ~70%;日照時間長,太陽輻射強,年均日照時間2 158.7 h,太陽輻射年總量為6 194 MJ/m2[8].草甸和灌叢是當地的主要植被類型,草甸分為帶有高寒性質的亞高山灌叢草甸、亞高山草甸和沼澤草甸類型[9].區內土壤類型以亞高山草甸土、沼澤土和高山草甸土為主,其中亞高山草甸土占全縣土地總面積的55.4%[10].
測定點位于紅原縣城以北10 km的平坦寬谷中,選取了一塊約400 m2的沼澤草甸樣地(102°32'E,33°03'N;海拔:3 514 m 左右),該草地為季節性積水,積水面積為30% ~40%,雨季積水2~5 cm.群落優勢種主要有:四川嵩草(Kobresia setchwanensis Hand.-Mazz)、西藏嵩草(Kobresia tibetica Maxim)、木里苔草(Carex muliiensis Hand.-Mazz)、烏拉苔草(Carex meyeriana Kunth)等,主要物種見表1.樣地群落總蓋度在90%以上[11],土壤類型為泥炭沼澤土,成土母質以河湖沉積物為主.

表1 樣地的主要物種組成Table 1 The composition of plant species at sampling sites
1.2 測定方法及數據處理
1.2.1 光合作用測定 野外實驗選在草本生長旺季的7月8-16日進行.選擇晴朗無云的天氣,7:00-19:00,每2 h測定一次.分別隨機選取8株生長良好的木里苔草和四川嵩草,選取植株的中上部向陽的成熟的葉片重復測定2次,共16個數據取平均值.由于無法在同一時間同時測定2種草本,所以采取每株植物輪流測定的方法,即第1次以木里苔草→四川嵩草的順序來測定,第2次就以相反的順序來測定,第3次的測定順序又與第1次的相同,以此類推,以消除2種草本測定時間上的誤差.
用LI-6400光合測定儀測定葉片的光合速率(Pn)和蒸騰速率(Tr),并同步記錄光合參數:有效輻射強度(PAR)、大氣 CO2濃度(ca,)、氣溫(Tair)、大氣相對濕度(RH)、葉片氣孔導度(Gs)、胞間CO2濃度(ci)和葉片溫度(Tleaf).
最后根據以上參數,計算出葉片水分利用效率(WUE=Pn/Tr)[12]和氣孔限制值 (Ls=1-ci/ca)[13].木里苔草的葉面積使用AM300葉面積儀進行測定;四川嵩草的葉面積采用幾何計算方法進行測定,先使用游標卡尺測量針葉的平均直徑和長度以獲取總表面積,再乘以0.5得到半球表面積[14].
1.2.2 葉綠素熒光參數測定 與光合作用同步測定,測定方式同理于光合測定.利用 PAM-2100(Walz,Germany)便攜式葉綠素熒光儀測定葉綠素熒光參數,葉片暗適應30 min后,打開弱測量光測定初始熒光(Fo),再打開一次飽和脈沖光(PFD為8 000 μmol·m-2·s-1,頻率為 20 kHz,0.8 s,1 個脈沖)測定最大熒光(Fm).然后在自然光下,光下穩態熒光(Ft)穩定后,再打一次飽和脈沖光測定光下最大熒光(F'm).得出光系統II(PSII)的反應中心
原初光能轉化效率為
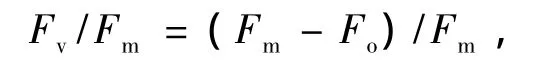
實際光化學反應效率為
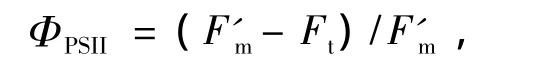
和電子傳遞速率[15]為
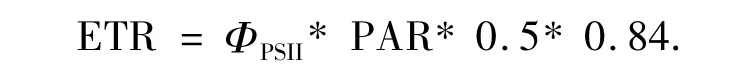
電子傳遞速率響應曲線的測量,7:00-19:00,每2 h測定一次.測量光為波長650 nm的紅光,強度為0.1 μmol·m-2·s-1.光化光和飽和脈沖光均為鹵素燈發出的白光,其中飽和脈沖光的強度大于5 000 μmol·m-2·s-1,脈沖時間 0.8 s.為避免外界光對植物生理狀態的影響,作電子傳遞速率響應曲線時用小黑布罩住植物[16].熒光測量由電腦控制,步驟為:打開測量光并打開光化光,適應10 s后打開飽和脈沖光,升高光化光強度(升高1),適應10 s后再打開飽和脈沖,如此重復5次.
將測定的電子傳遞速率響應曲線通過擬合[17],得到最大電子傳遞速率(ETRmax)、光能利用效率(α)、半飽和光強(Ek).擬合方程式為
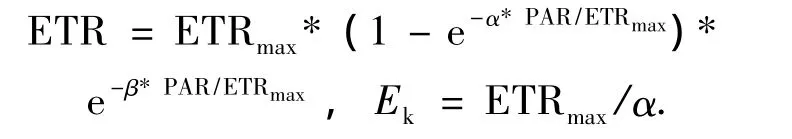
用SPSS12和Originpro 7.5軟件對所得實驗數據進行統計、分析和圖形處理.
2 結果與分析
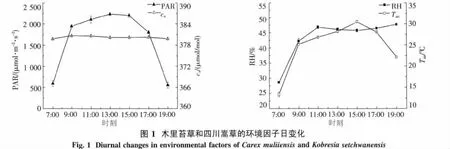
2.1 環境因子日變化規律 圖1表明,光合有效輻射(PAR)的日變化呈單峰曲線,最高峰出現在13:00左右,此時PAR值達到2 237.43 μmol·m-2·s-1,日平均 PAR 為 1 634.43 μmol·m-2·s-1.大氣CO2濃度(ca)基本保持不變,日均值為380.2 μmol/mol.相對濕度(RH)則在11:00達到峰值,此時RH為46.87%,其后隨著光合有效輻射、氣溫的升高而逐漸下降,15:00降至最低,之后隨著光合有效輻射和氣溫的下降又略有升高.氣溫(Tair)主要受PAR的影響,7:00左右為最低值(13.33℃),此后隨著光合有效輻射的增加而迅速上升,到15:00達到峰值(30.50℃),其峰值比PAR略滯后,日平均氣溫為24.88℃(圖1).
2.2 光合速率及其參數日變化特征
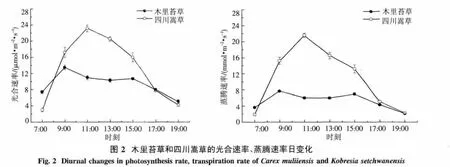
2.2.1 光合速率和蒸騰速率日變化特征 木里苔草的Pn日變化表現為典型的雙峰曲線,有明顯的“午休”現象,第一個峰值出現在9:00左右,為13.5 μmol·m-2·s-1,第二個峰值出現在15:00左右,為 10.7 μmol·m-2·s-1,在 13:00 左右出現低谷(圖2).由此可以看出,木里苔草Pn的第二個峰值低于第一個峰值,且中午Pn下降的速率均大于下午Pn回升的速率,即Pn中午降低快而恢復慢.四川嵩草的Pn日變化表現為典型的單峰曲線,在11:00達到峰值,為 23.2 μmol·m-2·s-1,而后又迅速下降.木里苔草與四川嵩草的Pn日平均值之間存在極顯著差異(P<0.01).2種草本所處的環境條件相同,而Pn日動態曲線表現出不同規律,四川嵩草Pn上升和下降速率均大于木里苔草,說明植物光合速率主要由本身的生理生態特性所決定.Tr日變化趨勢和Pn日變化相似,木里苔草Tr表現為雙峰曲線,而四川嵩草依然為單峰曲線,嵩草Pn上升和下降速率均大于木里苔草.木里苔草Tr分別在9:00及15:00左右出現峰值;而四川嵩草Tr出現在11:00左右,木里苔草與四川嵩草的Tr日平均值之間存在極顯著差異(P<0.01).
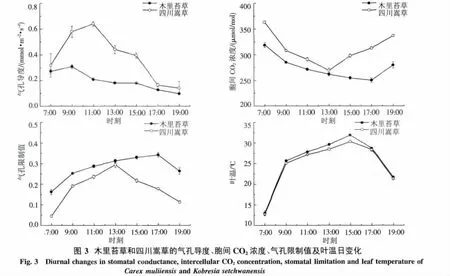
2.2.2 氣孔導度和胞間CO2濃度日變化特征 四川嵩草Gs日變化表現為單峰曲線,從早晨開始隨著PAR和Tair的增加,Gs逐漸增加,到11:00左右出現峰值,之后由于高溫低濕的環境導致Gs迅速降低.但木里苔草Gs日變化也表現為微弱的雙峰態勢,9:00左右出現最高值,9:00-13:00呈遞減趨勢,Gs較低且日變化波動較小.四川嵩草的Gs日平均值極顯著高于木里苔草(P<0.01).
四川嵩草的胞間CO2濃度(ci)在一天中呈早晚高、中午低的近“V”型變化趨勢.7:00為363.0 μmol/mol,隨著Pn的加強,葉片固定較多的CO2致使ci迅速下降;13:00左右達到低谷,之后一直呈上升趨勢,到19:00達到336.9 μmol/mol.但木里苔草的ci的日變化規律表現出7:00最高,之后逐漸降低,在17:00達到全天的最低值.四川嵩草的ci日平均值極顯著高于木里苔草(P<0.01).
2.2.3 氣孔限制值及葉溫日變化 四川嵩草和木里苔草的Ls日變化都呈現出單峰型曲線,2個峰值分別出現在13:00左右和17:00左右.木里苔草Ls在17:00之前保持緩慢上升,之后逐漸下降.而四川嵩草Ls在13:00之前急劇增大,之后迅速降低(圖3).從Ls的日平均值的大小看,與ci表現出相反的趨勢,木里苔草Ls顯著高于四川嵩草(P<0.01).
木里苔草和四川嵩草的葉面溫度日變化規律與氣溫日變化完全一致,日均葉溫分別為25.5℃和24.8℃.木里苔草(r=0.998,P<0.01)和四川嵩草(r=0.891,P<0.01)的葉面溫度都與大氣溫度呈極顯著正相關,所以葉片溫度主要受氣溫影響.
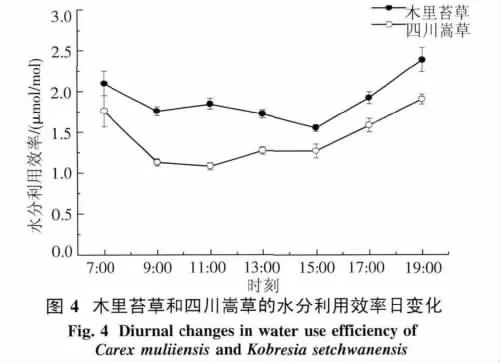
2.2.4 水分利用效率日變化 木里苔草WUE在7:00最大,為 2.1 μmol/mol,之后隨著光合有效輻射和葉溫的增加而降低,9:00-15:00,木里苔草WUE 在1.6~1.8 μmol/mol范圍內波動,15:00之后又迅速上升.四川嵩草WUE日變化呈早晚高,中午低的趨勢,7:00表現為最大,11:00降至最低,11:00-15:00都較為平穩的波動,15:00又迅速上升(圖4).木里苔草的WUE日平均值極顯著高于四川嵩草(P<0.01).
2.3 光合速率與生理生態因子的相關性分析
2.3.1 木里苔草凈光合速率與生理生態因子的關系 木里苔草凈光合速率與生理生態因子通徑分析和相關分析結果(表2和表3)表明:從相關系數的大小看來,生理生態因子對木里苔草的Pn影響的大小順序為:Tr>PAR>Gs>Tleaf>Tair>ca>ci>RH,其中,Tr、PAR、Gs、Tleaf、Tair、ca與 Pn呈極顯著正相關,而ci與Pn呈顯著的負相關.各生理生態因子對凈光合速率直接通徑系數的大小依次為:Tr>RH>Gs>PAR>ca>Tair>Tleaf>ci;Tr正向的直接作用遠大于其對Tair和Tleaf的負向間接作用,故表現為其與Pn呈顯著的正相關性.生理因子的決策系數順序為:R2(Tr)>R2(Gs)>R2(PAR)>R2(ca)>R2(RH)>R2(ci)>R2(Tair)>R2(Tleaf),其中,R2(RH)、R2(ci)、R2(Tair)和 R2(Tleaf)均小于 0.所以,影響木里苔草Pn日變化規律的決定生理生態因子為Tr、Gs、PAR 和 ca,其中 Tr為主要決定因子;限制因子為 RH、ci、Tair和 Tleaf,其中,Tleaf為主要限制因子.
2.3.2 四川嵩草凈光合速率與生理生態因子的關系 四川嵩草凈光合速率與生理因子通徑分析和相關分析結果(表3 和表 4)表明:PAR、ca、RH、Tair、Gs、Tr、Tleaf與 Pn呈極顯著正相關,而 ci與 Pn呈顯著的負相關.各生理因子對凈光合速率直接通徑系數的大小依次為:Tr>Gs>PAR>RH>ca>Tair>Tleaf> ci;盡管 ci通過 ca、RH、Tair和 Tleaf對 Pn的間接作用均為正值,但ci直接作用(-0.431)遠高于這些間接作用,所以沒有影響ci與Pn呈極顯著負相關;Tleaf對Pn的直接作用不大(-0.138),但被Tr正間接作用(0.483)所掩蓋而使其與Pn正相關.生理生態因子的決策系數順序為:R2(Tr)>R2(ci)>R2(Gs)>R2(PAR)>R2(RH)>R2(ca)>R2(Tair)>R2(Tleaf),其中,R2(ca)、R2(RH)、R2(Tair)和R2(Tleaf)小于0.所以,影響木里苔草Pn日變化規律的決定生理生態因子為Tr、Gs、PAR和ci,其中,Tr為主要決定因子;Tleaf為主要限制因子.
2.4 葉綠素熒光參數
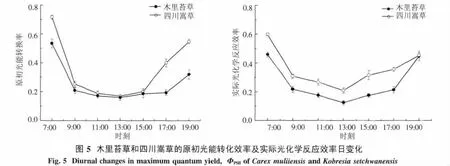
2.4.1 原初光能轉化效率和實際光化學反應效率由圖5看出,2種草本植物Fv/Fm和ΦPSII的日動態趨勢相似,7:00最高,并隨著光合有效輻射的增強而逐漸降低,在13:00降至最低,表明此時發生了光抑制現象;下午隨著光強的減弱又逐漸升高,說明其光抑制到下午又逐漸得到恢復,19時2種草本Fv/Fm并未恢復到7:00的水平.木里苔草13時Fv/Fm相對于其7:00降低了70.3%,而四川嵩草13時Fv/Fm相對于其7:00降低了76.5%.早晚時,四川嵩草與木里苔草Fv/Fm之間存在極顯著差異(P<0.01),表明四川嵩草葉片PSII的潛在活性和原初光能轉換效率均高于木里苔草;而且四川嵩草ΦPSII日平均值極顯著高于木里苔草(P<0.01).
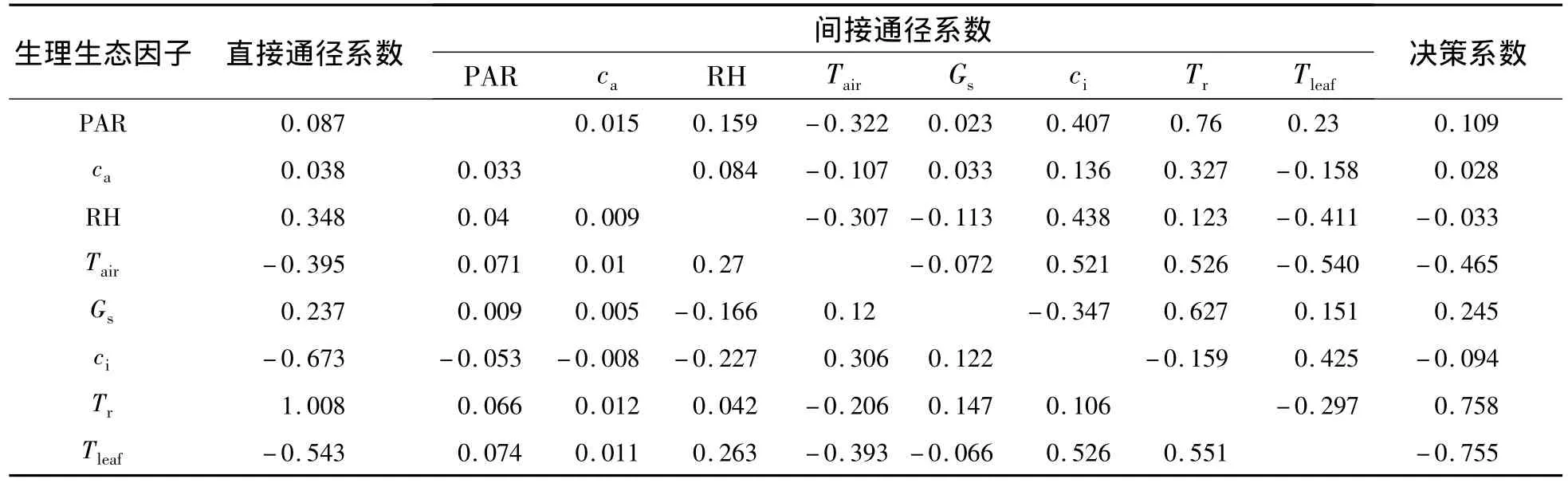
表2 木里苔草的凈光合速率與生理生態因子通徑系數Table 2 Path coefficients of Pnand eco-physiological factors in Carex muliiensis
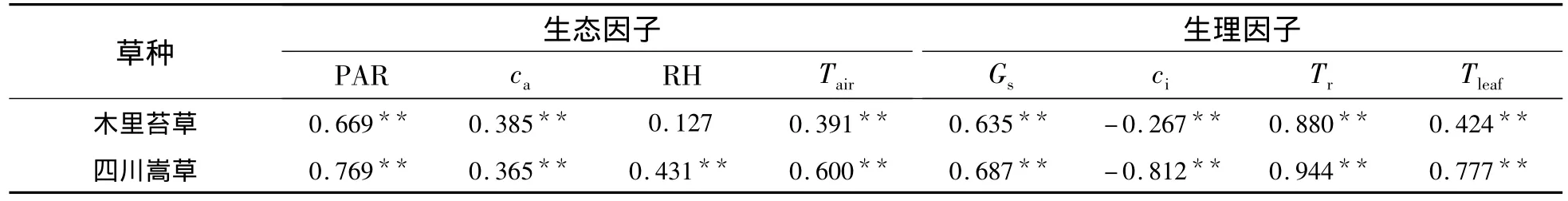
表3 凈光合速率與生理生態因子相關系數Table 3 Correlation coefficients of Pnand eco-physiological factors
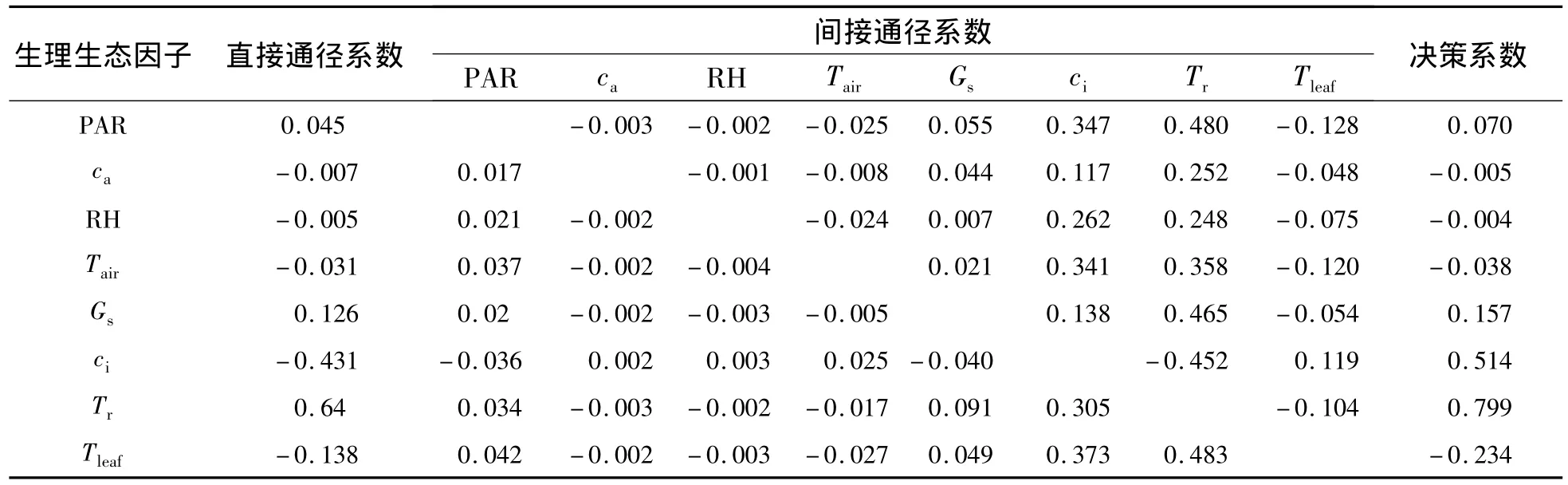
表4 四川嵩草的凈光合速率與生理生態因子通徑系數Table 4 Path coefficients of Pnand eco-physiological factors in Kobresia setchwanensis
2.4.2 快速光響應曲線 四川嵩草的ETRmax與Pn日變化相似,都呈單峰曲線,在11:00達到峰值,之后ETRmax逐漸降低.木里苔草ETRmax日變化呈微弱雙峰曲線,第一個峰值出現在9:00左右,之后降低,在13:00左右出現低谷,隨后略有回升,第二個峰值出現在15:00左右.另外,四川嵩草和木里苔草的Ek日變化都呈單峰曲線,分別在11:00和9:00達到峰值(圖6).四川嵩草的ETRmax和 Ek日平均值都極顯著高于木里苔草(P<0.01).
四川嵩草的α一天中呈早晚高、中午低的近“V”型變化趨勢,11:00降至最低,之后逐漸上升.但木里苔草α日變化呈“W”型變化趨勢,7:00-11:00較高且波動較小,之后降低,13:00左右出現低谷,隨后再上升,15:00出現峰值,之后呈現再下降及上升的趨勢.
3 討論
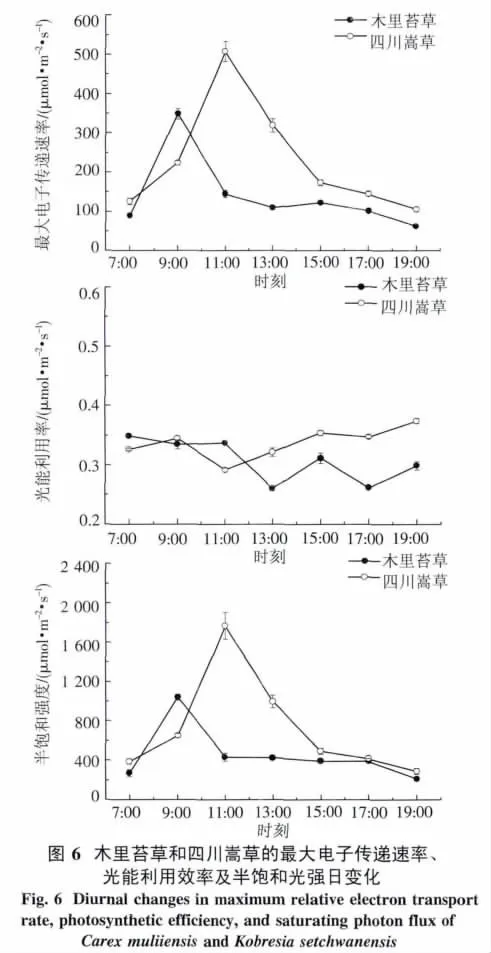
3.1 木里苔草與四川嵩草的光合特性 本試驗四川嵩草Pn、Tr和Gs的日變化都呈單峰型曲線,最高峰均出現在11:00,由于11:00以前,土壤水分較充足,高原PAR強且增加較快,植物Pn迅速提高,植株通過增大氣孔使外界CO2迅速向細胞內擴散以滿足光合作用的需要,使得Gs增大,蒸騰速率加快;11:00以后隨著光照強度的增強,過強的植物蒸騰和土壤蒸發量大[18],使植物體劇烈失水,而四川嵩草為淺根植物[19],植物根系得不到充足的供應,產生一定的信息物質(如脫落酸等),使葉片的氣孔開張度減少蒸騰失水[20-21],Pn也隨之下降,始終處于較低水平,所以四川嵩草Pn、Tr和Gs的日變化均呈單峰曲線,這與三江平原濕地毛苔草(Carex lasiocarpa Ehrh)研究結果相一致[22].但是,木里苔草Pn和Tr的日變化均呈典型的“雙峰”曲線.日出之后,隨著 PAR、Tair、Gs的逐漸增加,9:00左右達到最大值,之后強光照使葉片蒸騰失水過多,當在水分供應不充足時植物體多采用關閉氣孔而減少水分散失,氣孔的關閉造成ci降低,迫使光合速率下降[23],出現了光合“午休”現象.午后,由于光照減弱,使氣孔導度略有回升,Pn在15:00達到第二次高峰,這也受益于它們具有發達的根系[24],與四川嵩草相比具有較高的WUE,可以充分吸收土壤水分補充蒸騰失水來滿足生長需要,所以植物葉子的水分是對氣孔的開閉有著強有力的控制作用.
木里苔草和四川嵩草Pn的日變化都呈現出光合“午休”現象,這與青藏高原東北隅矮嵩草(Kobresia humilis)[25]、老芒麥(Elymus sibiricus)[26]以及青海高原海北高寒草甸的垂穗披堿草(Elym usnutans)的研究結果相一致.光合作用的“午休”是2種草本植物在高寒沼澤草甸形成的對付環境脅迫的一種表現,這避免了植物的過度失水和保護了光合機構,并且加強了熱耗散過程的運作[27].光合作用除了受到環境因素的影響外,其光合和蒸騰的大小其實還受到氣孔導度的影響[28].植物葉片Pn中午降低的植物自身因素不外乎2個:即氣孔因素和非氣孔因素.前者是由于中午光照、溫度和濕度等環境因子的變化引起植物氣孔的部分關閉,CO2進入葉片受阻而使光合下降,而后者是由于葉肉細胞自身羧化酶活性的下降而引起Pn的降低[29].根據G.D.Farquhar等[30]的觀點,只有當 Pn和 ci變化方向相同,兩者同時減小,且Ls增大時,才可以認為Pn的下降主要是由Gs引起的,否則Pn的下降要歸因于葉肉細胞羧化能力的降低.木里苔草在9:00-13:00期間Pn降低、ci下降、Ls增大,因此可以推斷出引起光合“午休”的主要原因是氣孔因素;午后15:00-17:00期間,Pn的下降主要也受氣孔因素限制.而四川嵩草在11:00-13:00階段,Pn與ci都下降,Ls上升,說明Pn的下降受氣孔因素限制;13:00-19:00階段,Pn的下降伴隨著ci上升,Ls下降,說明Pn的下降主要受非氣孔因素限制.研究表明,植物水分虧缺時光合速率的降低并不是由于水分供應不足直接引起的,而是由水分虧缺造成的氣孔或非氣孔因素限制,輕度水分虧缺以氣孔限制為主,而較重水分脅迫下則以非氣孔限制為主[31].說明氣孔關閉對水分虧缺較敏感,木里苔草主要受到輕微水分脅迫,較低的Tr和較高的WUE顯示出在受水分脅迫的高溫低濕環境下的適應性,所以午后由于光照減弱,Pn再升高達到第二次高峰,日變化呈典型的“雙峰”曲線.同理,四川嵩草主要受到較重水分脅迫,因受強烈PAR影響,植物蒸騰和土壤蒸發量大,而四川嵩草植物又多為淺根系植物,故為減少水分蒸發將氣孔開度逐漸減小[32],Pn也隨之下降,并難以恢復到較高水平,日變化呈典型的“單峰”曲線.所以,水分供應狀況對2種植物的光合作用非常重要,而植物蒸騰是植物調節體內水分平衡的主要環節,植物通過調節蒸騰作用使光合作用最大化,減少水分散失并降低葉面溫度、免受強光的灼傷[33],本研究結果也表明,2種草本植物光合作用Pn的主要決定因子是Tr.
與四川嵩草相比,木里苔草的WUE較高,說明生產相同量的物質消耗水分數量較少.木里苔草是沼澤濕地莎草科植物,多生長于沼澤草甸及低濕草地中,它兼具有水生和陸生植物共有的某些特征,適應性廣,也有發達的根莖[34],長期適應沼澤草甸的環境中生長,可能使其具有較高的WUE.有研究表明,莎草科植物普遍具有較高的WUE[35],在生長過程中高效地使用了水分,進而保證了它們在沼澤濕地中作為優勢種的地位[36].另外,本研究發現,2種草本植物WUE日變化均呈現早晚高,中午低的趨勢,這是因為研究區早晨氣溫低,植物能以較低的水分代價得到更多的光合產物所致.隨著溫度升高,葉片氣孔導度和蒸騰速率增加,這是對高溫和強輻射的適應性策略,但卻顯著降低了光合速率和水分利用效率,這與內蒙古錫林浩特典型草原的羊草(Leymus Chinensis)具有相似的特征[37].
2種草本植物Pn、Gs和PAR的日變化趨勢基本一致(圖1~3),Pn與 PAR和 Gs呈正相關(表3).Pn與PAR的相關性大于與Gs的相關性(表3),而 Gs受光的誘導[38-39],可以認為生態因子PAR是影響2種草本植物光合作用的重要因子,這與青海高寒草甸的麻花艽(Gentiana straminea Maxim)研究結果相一致[40].低光強下隨光強的升高Gs[39]、光合作用暗反應的酶活性升高[38],光合傳遞的電子增多(圖6),光合作用加快.Pn與ci呈負相關,表明光合碳固定隨光強升高而增加的速度大于Gs隨光強增加的速度,導致ci隨光強的升高而降低(圖3),即強光下ci降低是由于光合作用利用CO2增加的結果.
3.2 木里苔草與四川嵩草的葉綠素熒光特性Fv/Fm反映了PSII反應中心潛在光能轉化效率,即PSII反應中心處于開放狀態時的量子產量.正常生長的植物的Fv/Fm值通常約為0.8~0.85之間[41],當植物受到水分脅迫、極端溫度和鹽脅迫等逆境脅迫后Fv/Fm值會不同程度下降,因此,Fv/Fm是逆境生理研究的重要指示性參數.本研究結果顯示,清晨7:00,木里苔草和四川嵩草的Fv/Fm都低于上述范圍,所以認為清晨環境對二者的生長構成抑制或脅迫,發生了長期的光抑制.這是因為,高原地區清晨的低溫使植物的光合機構對光抑制更加敏感,低溫下較低的光強也會引起光抑制[42].而且,四川嵩草7時Fv/Fm顯著高于木里苔草,清晨時木里苔草光抑制程度更大,可能導致木里苔草的Pn上升速率較四川嵩草緩慢.四川嵩草Fv/Fm值高于木里苔草,也表明四川嵩草的光能轉化效率也更高,適應環境變化的潛力更強,高的光化學轉化效率有利于其維持較高的能量消耗和適應氣溫的急劇變化,因此四川嵩草的ETRmax較高.本研究結果表明,2種草本植物的Fv/Fm隨日照時間的延長會不斷降低,表明這2種植物遭受光抑制,光照越強光抑制現象越嚴重.13:00,四川嵩草Fv/Fm相對于其7:00降低了76.5%(圖5),與木里苔草相比光抑制程度更大,植物水分脅迫較重,從而導致其光合速率的迅速降低.另外,19時2種草本Fv/Fm未恢復到7:00的水平,表明PSII反應中心可能發生了不可逆失活,或光合機構受到了破壞或損傷[43].這可能與青藏高原獨特環境有關,低溫和強光并存的環境條件往往造成植物較嚴重的光抑制[44].進一步說,在遭受低溫的情況下,大多數植物的光抑制恢復較慢[45],是由于修復 PSII的 D-1蛋白質速率降低[46].本研究的通徑分析也表明,2個草本植物的Pn,Tleaf為主要限制因子(表3和表4).
ΦPSII反映的是光系統II反應中心部分關閉情況下的實際光化學效率,與碳同化反應的強度密切相關.在中午強光、較高氣溫、低濕脅迫下,ΦPSII明顯下降,午后又都有所恢復,說明強光脅迫使2種草本植物開放的光系統II反應中心部分關閉,光能轉換效率下降,從而降低了電子傳遞能力,導致用于CO2同化的光能減少.其次,從7:00-19:00,四川嵩草的ΦPSII大于木里苔草,說明PSII的實際光能捕獲效率較高,能夠把所捕獲的光能更多地用于光化學反應[47],因此四川嵩草的光合能力ETRmax較高.
PSII的相對光合電子傳遞速率(ETR)是反映實際光強下的表觀電子傳遞速率,用于度量光化學反映導致碳固定的電子傳遞情況.將測定的電子傳遞速率響應曲線通過擬合,得到最大電子傳遞速率(ETRmax)、光能利用效率(α)和半飽和光強(Ek)的日變化.四川嵩草Ek顯著大于木里苔草,說明其耐受強光.另外,2種草本植物ETRmax和Ek的變化趨勢一致.這是因為Ek反映了植物耐受強光的能力[48],而耐受強光能力強的植物具有較高的最大電子傳遞速率ETRmax,所以四川嵩草有更大的光適應范圍和較強的耐受強光能力,四川嵩草的ETRmax和Pn較木里苔草晚1 h到達峰值.由圖6看出,從7:00-19:00,2個草本植物的Ek一直低于PAR,可見木里苔草一天中有12 h處在光飽和狀態.本研究表明,2種草本植物光能利用效率α日變化趨勢表現為清晨和傍晚較高,正午較低.這是因為,α反映了葉片捕光能力的高低,在中午光強太高,為了避免吸收的光能太多造成損傷,植物可以通過降低捕光能力α來保護自身,說明有調控光合作用和高效利用強光的能力.PAR最強13:00,四川嵩草α高于木里苔草,這說明上午光合有效輻射較強時,四川嵩草α較高,對強光耐受能力強.
4 結語
綜上所述,川西北高寒沼澤草甸2種植物的光合生理特征之間有著較大的差異,這些差異在一定程度上是對外界變化生境適應的結果.木里苔草Pn和Tr的日變化呈雙峰曲線,而四川嵩草則呈單峰曲線,兩者都呈現光合“午休”現象,木里苔草的光合“午休”主要受氣孔因素限制,但四川嵩草主要受非氣孔因素限制.沼澤濕地植物木里苔草,它兼具有水生和陸生植物共有的某些特征,也有發達的根莖,長期適應沼澤草甸的環境中生長,較低的Tr和較高的WUE顯示出在受水分脅迫的高溫低濕環境下的適應性;但高寒草甸四川嵩草具有較高實際光化學反應效率(ΦPSII)和半飽和光強(Ek),表明四川嵩草對強光耐受能力強,高原沼澤草甸不同植物即使在相同環境條件下具有不同適應對策.2種草本植物光合作用Pn的主要決定因子是Tr,高寒沼澤草甸2種植物光合對環境的適應取決于水分狀況,水分是植物生長的主要因子.過多的土壤水分在生長季節通過植物蒸騰的散失,從而使土壤含水量進一步減少,這在某種程度上加劇高寒沼澤濕地的退化.所以要保護和恢復濕地資源,合理利用濕地的植物資源,以及在已經退化的濕地中補充高WUE的土著濕地物種,都是在現有條件下保護和恢復濕地的策略.
致謝 成都大學校青年基金項目(2013XJZ23)對本文給予了資助,謹致謝意.
[1]Wu Q B,Li X,Li W J.The prediction of permafrost change along the Qinghai-Tibet highway,China[J].Permafrost and Periglacial Processes,2000,11(4):371-376.
[2]王俊峰,吳青柏.青藏高原沼澤草甸區植被退化對淺層寒凍土壤環境的影響[J].蘭州大學學報:自然科學版,2011,47(6):39-45.
[3]徐洪靈,張宏,張偉.川西北高寒草甸土壤呼吸速率日變化及溫度影響因子比較[J].四川師范大學學報:自然科學版,2012,35(3):405-411.
[4]張新時,周廣勝,高瓊,等.中國全球變化與陸地生態系統關系研究[J].地學前緣,1997,4(1/2):137-144.
[5]盧存福,卉桂英.矮嵩草光合作用與環境因素關系的比較研究[J].植物生態學報,1995,19(1):72-78.
[6]Cui X Y,Niu H S,Wu J,et al.Response of chlorophyll fluorescence to dynamic light in three alpine species differing in plant architecture[J].Environmental and Experimental Botany,2006,58:149-157.
[7]張宏,張偉,徐洪靈.川西北高寒草甸生長季土壤氮素動態[J].四川師范大學學報:自然科學版,2011,34(4):583-588.
[8]四川省紅原縣志編撰委員會.紅原縣志[M].成都:四川人民出版社,1996.
[9]劉長秀,張宏,澤柏.灌叢對川西北高寒草甸土壤資源的影響[J].山地學報,2006,24(3):357-365.
[10]田應兵,熊明標,宋光煜.若爾蓋高原濕地土壤的恢復演替及其與養分變化[J].生態學雜志,2005,24(1):21-25.
[11]張偉,張宏.青藏高原東緣紅原地區三種不同草甸土壤活性碳特征[J].山地學報,2008,26(2):205-211.
[12]張正斌,山侖.作物水分利用效率和蒸發蒸騰估算模型的研究進展[J].干旱地區農業研究,1997,15(1):73-78.
[13]許大全.光合作用效率[M].上海:上海科學技術出版社,2002.
[14]Wykoff W R.Measuring and modeling surface area of ponderosa pine needles[J].Canadian J Forest Research,2002,32(1):1-8.
[15]尹華軍,賴挺,程新穎,等.增溫對川西亞高山針葉林內不同光環境下紅樺和岷江冷杉幼苗生長和生理的影響[J].2008,32(5):1072-1083.
[16]李暢,蘇家樂,劉曉青,等.一品紅不同品種葉片葉綠素熒光特性比較[J].園藝學報,2009,36(10):1519-1524.
[17]Platt T,Gallegos C L,Harrison W G.Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton[J].J Marine Research,1980,38:687-701.
[18]李英年,趙亮,徐世曉,等.祁連山海北高寒濕地植物群落結構及生態特征[J].冰川凍土,2006,28(1):76-84.
[19]周興民.中國嵩草草甸[M].北京:科學出版社,2001.
[20]Zhang J W,Davies J.Increased synthesis of ABA in partially dehydrated root tips and ABA transport from roots to leaves[J].J Experimental Botany,1987,38:2015-2023.
[21]徐惠風,劉興土,金研銘,等.沼澤植物澤瀉氣孔導度日變化的研究[J].生態科學,2003,22(3):218-221.
[22]欒金花.干旱脅迫下三江平原濕地毛苔草光合作用日變化特性研究[J].濕地科學,2008,6(2):221-228.
[23]李文瑞,馮金朝,江天然,等.沙冬青幾種光合特性的季節性變化的研究[J].植物學報,1999,41(2):190-193.
[24]趙佐成.四川省紅原縣水生植物群落調查[J].武漢植物學研究,1996,14(3):213-222.
[25]師生波,韓發,卉桂英.高寒草甸矮嵩草群落光合作用的“午休”現象[J].植物生理學報,1997,23(4):405-409.
[26]王巖春,干友民,邱英,等.高寒地區“川草2號”老芒麥夏季光合生理生態特性的初步研究[J].草業與畜牧,2008(1):1-3.
[27]陳曉亞,湯章程.植物生理與分子生物學[M].北京:高等教育出版社,2007.
[28]陳模舜,柯世省,倪瓊瓊.田間珊瑚樹凈光合速率及生態因子的日變化[J].四川師范大學學報:自然科學版,2004,27(3):298-302.
[29]杜占池.羊草和大針茅光合作用午間降低與生態因子關系的研究[J].自然資源學報,1990,5(2):177-186.
[30]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Ann Rev Plant Physiol,1982,33:317-345.
[31]薛崧,汪沛洪,許大全,等.水分脅迫對冬小麥CO2同化作用的影響[J].植物生理學報,1992,18(1):1-7.
[32]師生波,賁桂英,韓發.矮嵩草草甸植物群落的光合特性研究[J].植物生態學報,1996,20(3):225-234.
[33]Sparks J P,Black R A.Regulation of water loss in populations of populus trichocarpa:the role of stomatal control in preventing xylem cavitation[J].Tree Physiol,1999,19(7):453-459.
[34]趙佐成,魏泰昌,邱發英,等.若爾蓋轄曼-阿西高原沼澤自然保護區沼澤植物特征[J].武漢植物學研究,1987,5(8):235-246.
[35]李宏林,徐當會,杜國禎.青藏高原高寒沼澤濕地在退化梯度上植物群落組成的改變對濕地水分狀況的影響[J].植物生態學報,2012,36(5):403-410.
[36]Wang Y S,Chu C J,Maestre F T,et al.On the relevance of facilitation in alpine meadow communities:an experimental assessment with multiple species differing in their ecological optimum[J].Acta Oecologica,2008,33:108-113.
[37]許振柱,周廣勝,李暉.羊草葉片氣體交換參數對溫度和土壤水分的響應[J].植物生態學報,2004,28(3):300-304.
[38]翁曉燕,蔣德安,陸慶.影響水稻光合日變化的酶和相關因素的分析[J].生物數學學報,1999,14(4):495-500.
[39]趙平,曾小平,彭少麟,等.海南紅豆(Ormosia pinnata)夏季葉片氣體交換、氣孔導度和水分利用效率的日變化[J].熱帶亞熱帶植物學報,2000,8(1):35-42.
[40]吳建國.UV-B輻射增強對麻花艽葉片光合作用及相關生理參數的影響[J].環境科學研究,2010,23(11):1395-1404.
[41]Genty B,Briantais J M,Baker N R.The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J].Biochimica et Biophysica Acta:General Subjects,1989,990(1):87-92.
[42] Huner N P A,Oquist G,Hurry V M.Photosynthesis,photoinhibition and low temperature acclimation in cold tolerant plants[J].Photosynth Res,1993,37:19-39.
[43]Kolek J,Kozinka V.Physiology of the Plant Root System[M].Dordrecht:Kluwer Academic Publishers,1991.
[44]郭連旺,沈允鋼,許大全,等.幾種高寒草甸常見植物的光合特性及其光合作用的光抑制[C]//高寒草甸生態系統:第四集.北京:科學出版社,1995:65-74.
[45]Greer D H,Laing W A.Photoinhibition of photosynthesis in intact kiwifruit Actinidia deliciosa leaves,recovery and its dependence on temperature[J].Planta,1988,174:15-165.
[46]Franklin L A.The effects of temperature acclimation on the photoinhibitory responses of ulva rotundata blid[J].Planta,1994,192:324-331.
[47]張杰,楊傳平,鄒學忠,等.蒙古櫟硝酸還原酶活性、葉綠素及可溶性蛋白含量與生長性狀的關系[J].東北林業大學學報,2005,33(3):20-21.
[48]Ralph P J,Gademann R,Dennison W C.In situ seagrass photosynthesis measured using a submersible,pulse-amplitude modulated fluorometer[J].Marine Biology,1998,132:367-373.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
