基于用戶畫像模型的農(nóng)業(yè)保險精準營銷研究
2021-09-10 22:43:42周紫薇
商業(yè)2.0-市場與監(jiān)管 2021年6期
摘要:近年來,“三農(nóng)”問題的關(guān)注度持續(xù)攀升,農(nóng)業(yè)保險的發(fā)展也關(guān)乎“三農(nóng)”問題的解決,調(diào)查農(nóng)戶種植現(xiàn)狀以及投保需求,依據(jù)收集的數(shù)據(jù)構(gòu)建農(nóng)戶畫像模型,實現(xiàn)農(nóng)險精準營銷具有重要意義。
關(guān)鍵詞:用戶畫像;精準營銷;農(nóng)業(yè)保險
近些年來,隨著社會不斷進步和科技不停地發(fā)展,農(nóng)業(yè)生產(chǎn)技術(shù)和生產(chǎn)水平得到了顯著地提高,但是現(xiàn)階段自然環(huán)境的不斷惡化,農(nóng)業(yè)生產(chǎn)仍面臨著巨大的挑戰(zhàn)。此外,我國的農(nóng)業(yè)生產(chǎn)規(guī)模大且生產(chǎn)類型多樣化,這使得農(nóng)戶在種植業(yè)中的投入不斷增多,導(dǎo)致了農(nóng)戶所面臨的農(nóng)業(yè)風(fēng)險在日漸增加。而農(nóng)業(yè)保險通過預(yù)防賠付農(nóng)業(yè)風(fēng)險損失,保障農(nóng)業(yè)生產(chǎn)者基本收入的同時,也一定程度上增強農(nóng)業(yè)生產(chǎn)者的抵抗風(fēng)險能力。因此需要積極研究農(nóng)業(yè)生產(chǎn)者農(nóng)險需求,推動農(nóng)業(yè)保險的創(chuàng)新式發(fā)展。
自2012年《農(nóng)業(yè)保險條例》提出農(nóng)業(yè)保險以來,我國農(nóng)業(yè)保險逐漸得到發(fā)展,農(nóng)險類別開始分化。目前,若按照不同的農(nóng)業(yè)種類,我國農(nóng)業(yè)保險可分為種植業(yè)保險和養(yǎng)殖業(yè)保險,如若按遇險類別則可劃分為自然災(zāi)害損失險、病蟲害損失險、疾病死亡險等類別。同時伴隨著國家持續(xù)加大對“三農(nóng)”的投入,我國農(nóng)險消費能力逐漸擴大發(fā)展起來。在高投入低回報及不確定因素較多的高風(fēng)險性背景下,農(nóng)業(yè)保險為農(nóng)戶撐起一把“保護傘”,對保障農(nóng)業(yè)生產(chǎn)、促進農(nóng)民增收起到積極作用。
隨著互聯(lián)網(wǎng)的迅速發(fā)展與普及,各類信息數(shù)據(jù)呈爆發(fā)式噴涌。我國作為農(nóng)業(yè)大國,有著龐大的農(nóng)業(yè)數(shù)據(jù)基礎(chǔ),伴隨著大數(shù)據(jù)在農(nóng)業(yè)領(lǐng)域的應(yīng)用,農(nóng)業(yè)數(shù)據(jù)類型變得更加豐富多樣化,此時借助于充足而廣泛的農(nóng)業(yè)數(shù)據(jù),對農(nóng)業(yè)保險與農(nóng)業(yè)生產(chǎn)者進行研究,實現(xiàn)農(nóng)戶的農(nóng)險個性化服務(wù),促進農(nóng)業(yè)生產(chǎn),意義尤為重大。
本項目在文獻研究的基礎(chǔ)上,以安徽省保險業(yè)和農(nóng)業(yè)為研究對象,通過對保險公司和農(nóng)險客戶進行實地調(diào)研,收集農(nóng)戶種植現(xiàn)狀、特點和利益訴求等信息,利用大數(shù)據(jù)分析處理,建立用戶畫像模型,分析用戶畫像特征,了解不同的農(nóng)戶特點及對農(nóng)險的需求,根據(jù)不同類型的農(nóng)戶特性,結(jié)合安徽本地種植養(yǎng)殖品種、農(nóng)業(yè)風(fēng)險的特色和重點需求,對農(nóng)業(yè)保險產(chǎn)品進行個性化的設(shè)計,使農(nóng)險產(chǎn)品盡可能的貼合農(nóng)戶、農(nóng)產(chǎn)品特點,滿足農(nóng)戶需求,更加精準高效應(yīng)對農(nóng)業(yè)風(fēng)險,進而實現(xiàn)精準營銷。用創(chuàng)新推動農(nóng)業(yè)保險的個性化,用個性化更好的發(fā)揮保險的保障作用,以促進農(nóng)業(yè)保險及農(nóng)村經(jīng)濟的健康快速發(fā)展。
1.研究現(xiàn)狀
1.1用戶畫像相關(guān)研究
用戶畫像即目標用戶的角色,它是一種專門用于描繪和了解目標客戶、聯(lián)系他們的用戶訴求和確定產(chǎn)品設(shè)計的有效手段。基于充分客戶信息前提下,將客戶的每一個具體化的信息都抽象成標簽,通過這些標簽把客戶信息進行了具體化,進而為客戶提供優(yōu)質(zhì)的個性化服務(wù)。近些年,伴隨著移動互聯(lián)網(wǎng)的普及以及大數(shù)據(jù)的興起,國內(nèi)外眾多專家學(xué)者參與到對用戶畫像的研究中,使得用戶畫像在各個領(lǐng)域都開始普及。在國內(nèi)學(xué)者中,郝勝宇和陳靜仁(2016)進行了關(guān)于企業(yè)用戶動態(tài)圖像技術(shù)的研究,致力于企業(yè)的精準化市場營銷;郭光明(2017)通過收集社交媒體數(shù)據(jù),構(gòu)建網(wǎng)絡(luò)社交用戶畫像,研究用戶信用屬性;劉穎(2020)以大數(shù)據(jù)為基礎(chǔ),將用戶信息進行分類,對數(shù)字圖書館進行用戶畫像模型建構(gòu),以實現(xiàn)圖書信息精準推送;而國外研究者中,Sumitkumar,Debajyoti和Sheetal(2016)則通過自動信息檢索,利用用戶畫像技術(shù)為大學(xué)推薦系統(tǒng)的用戶提供個性化指導(dǎo);An、Kwak和Jansen(2017)通過構(gòu)建用戶畫像,對YOUTUBE社交媒體上用戶進行興趣特征細分。
1.2農(nóng)業(yè)保險相關(guān)研究
農(nóng)業(yè)保險是為從事農(nóng)業(yè)生產(chǎn)過程的農(nóng)業(yè)生產(chǎn)者減輕因意外災(zāi)害帶來經(jīng)濟損失的保險。關(guān)于農(nóng)業(yè)生產(chǎn)者和農(nóng)業(yè)保險的研究中,國內(nèi)外不同學(xué)者從不同角度出發(fā)進行研究并得出不同結(jié)論。在國內(nèi),王阿星、張峭(2008)依據(jù)logistic模型對內(nèi)蒙古鄂爾多斯市的農(nóng)業(yè)生產(chǎn)者的農(nóng)業(yè)保險需求的影響因素進行研究,得出農(nóng)業(yè)生產(chǎn)者的農(nóng)業(yè)收入占比、受災(zāi)類型、受教育程度等顯著影響其農(nóng)業(yè)保險需求;程靜、劉飛和陶建平(2018)分析湖北省農(nóng)戶農(nóng)險決策時,研究發(fā)現(xiàn)影響農(nóng)戶農(nóng)業(yè)保險需求的最主要因素是其獲得資金援助機會、家庭勞動力比例以及土壤灌溉條件等;學(xué)者干潔(2019)采取Logistic計量經(jīng)濟模型對浙江紹興市農(nóng)戶的農(nóng)業(yè)保險態(tài)度及認知進行了分析與評價,研究得出農(nóng)業(yè)生產(chǎn)者對農(nóng)業(yè)保險的了解程度、戶主遭遇農(nóng)業(yè)災(zāi)害次數(shù)以及其風(fēng)險偏好均能影響農(nóng)業(yè)生產(chǎn)者對農(nóng)業(yè)保險的態(tài)度和認知;在國外學(xué)者中,Bruce(2004)研究發(fā)現(xiàn)農(nóng)戶的年齡以及風(fēng)險管理能力會一定程度上影響其購買保險的決策;Jason、Vincent (2012)通過建立模型對農(nóng)險需求量進行研究時,發(fā)現(xiàn)農(nóng)作物價格的提高能提升市場上農(nóng)業(yè)保險的需求量;Ruth Vargas Hill(2013)對農(nóng)戶購買農(nóng)險意愿進行實證分析時,得出受教育水平、財富的多少均會顯著影響農(nóng)戶農(nóng)險意愿;Anderson(2015)通過研究發(fā)現(xiàn)農(nóng)業(yè)生產(chǎn)者的財富水平并非和其農(nóng)險需求呈正相關(guān),當(dāng)其財富值達到一定程度時,其農(nóng)險需求可能會隨之降低。
綜上所述,結(jié)合以往學(xué)者的研究成果可知,用戶畫像是通過對用戶信息進行加工,從而將用戶進行特征分類以實現(xiàn)精準營銷。而對于農(nóng)業(yè)生產(chǎn)者購買農(nóng)業(yè)保險的決策,若從不同角度分析則其與多種特征因素相關(guān)聯(lián)。因此,將兩者結(jié)合,以安徽省為范圍,一方面對購買農(nóng)業(yè)保險的農(nóng)業(yè)生產(chǎn)者進行相關(guān)信息收集,將信息分類并標簽化。另一方面通過構(gòu)建用戶畫像模型,將這些農(nóng)業(yè)生產(chǎn)者進行特征細化,從而實現(xiàn)農(nóng)業(yè)保險的精準營銷。
1.3農(nóng)業(yè)保險用戶畫像模型構(gòu)建
1.3.1數(shù)據(jù)采集獲取數(shù)據(jù)與數(shù)據(jù)處理
為保證構(gòu)建的用戶畫像精準鮮明,數(shù)據(jù)采集應(yīng)該具備真實客觀性。因此將從兩個方面展開,一方面通過行業(yè)調(diào)研將安徽省保險公司銷售部門的農(nóng)險購買顧客的歷史記錄等相關(guān)信息數(shù)據(jù)進行整理收集;另一方面前往安徽省各農(nóng)村地區(qū)進行實地調(diào)研,通過用戶問卷、農(nóng)戶訪談等形式收集農(nóng)業(yè)種植養(yǎng)殖以及農(nóng)險相關(guān)信息數(shù)據(jù)。同時為確保數(shù)據(jù)的全面性和準確性,應(yīng)對新增農(nóng)業(yè)生產(chǎn)者的基礎(chǔ)信息數(shù)據(jù)、種植特征數(shù)據(jù),風(fēng)險數(shù)據(jù)等各類動態(tài)數(shù)據(jù)和靜態(tài)數(shù)據(jù)進行更新和補充。其次對各個渠道收集整理的數(shù)據(jù)清洗,剔除異常數(shù)據(jù),保證數(shù)據(jù)的高質(zhì)量。用戶畫像收集的數(shù)據(jù)范圍越廣,用戶特性越鮮明,就能很好的實現(xiàn)農(nóng)險的精準營銷。
1.3.2設(shè)定用戶畫像標簽體系
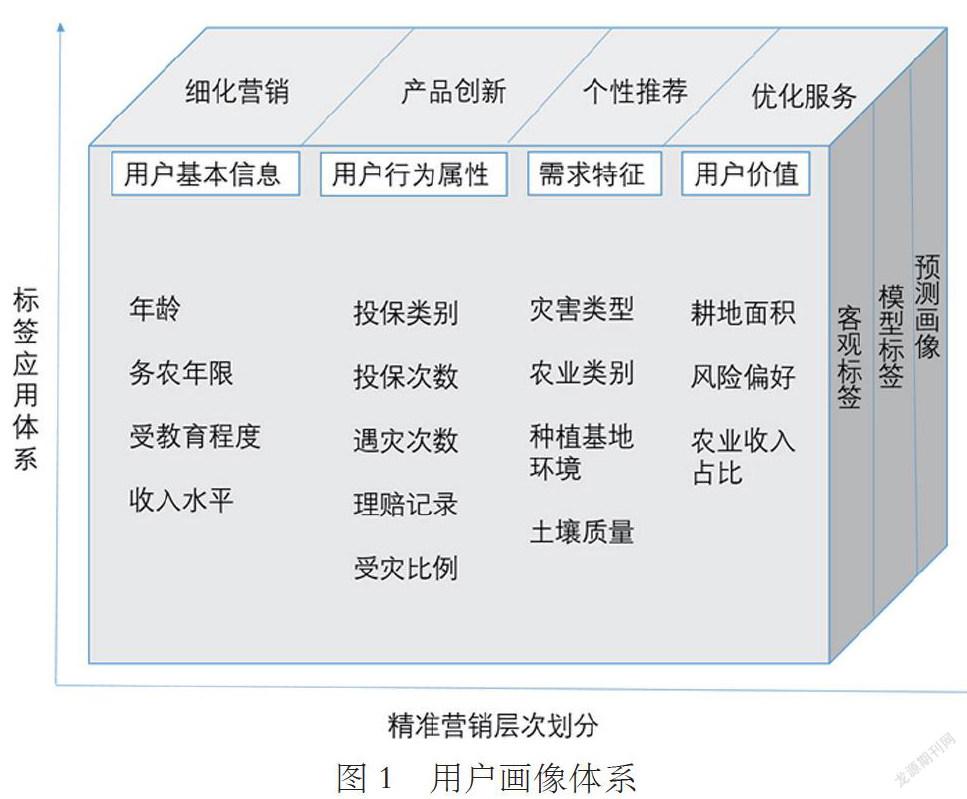
通過收集整理包括年齡、務(wù)農(nóng)年限、受教育程度、收入水平、投保類別、投保次數(shù)、受災(zāi)比例、遇災(zāi)次數(shù)、理賠記錄、受災(zāi)比例、災(zāi)害類型、農(nóng)業(yè)類別、種植基地環(huán)境、土壤質(zhì)量、耕地面積、風(fēng)險偏好、農(nóng)業(yè)收入占比等相關(guān)信息與數(shù)據(jù),并將抽取的數(shù)據(jù)進行標簽化,利用各類標簽的特性將用戶標簽化。同時可將標簽劃分為基本屬性、行為屬性、需求屬性、用戶價值。基于這些標簽,形成完整的用戶形象,完成用戶畫像模型構(gòu)建。利用用戶畫像標簽特性,將滿足用戶標簽的產(chǎn)品對用戶實行精準營銷,確保農(nóng)險銷售基于農(nóng)戶需求為前提。
1.3.3用戶畫像模型構(gòu)建
可構(gòu)建如下圖 1用戶畫像體系,將收集的已標簽化的數(shù)據(jù)進行處理,從而標準化各類用戶特質(zhì),了解用戶需求與愛好,構(gòu)建用戶畫像模型,進行業(yè)務(wù)層次劃分,進而實現(xiàn)精準營銷。
1.4精準營銷策略
1.4.1實施精準推廣,吸引潛在客戶
提升數(shù)據(jù)挖掘技術(shù),聘請專業(yè)數(shù)據(jù)分析人員。采用大數(shù)據(jù)、云計算形式,通過收集、挖掘數(shù)據(jù),全面追蹤用戶投保記錄。針對處理過的數(shù)據(jù),精細劃分,精確指標和標簽,分析潛在用戶偏好,線上線下交叉實施精準推廣方案,從而提供差異化產(chǎn)品服務(wù),吸引更多潛在的農(nóng)業(yè)生產(chǎn)者。
1.4.2精準營銷服務(wù),實現(xiàn)用戶乘數(shù)
基于用戶數(shù)據(jù)構(gòu)建用戶畫像,根據(jù)用戶價值進行客戶細分,從而挖掘用戶深層次需求,發(fā)現(xiàn)新需求,滿足其新需求,提升農(nóng)業(yè)生產(chǎn)者滿意度,創(chuàng)造長尾用戶。維持用戶與企業(yè)的連續(xù)溝通產(chǎn)生商業(yè)價值,創(chuàng)造用戶終身價值,實現(xiàn)用戶乘數(shù)。
1.4.3終端實時監(jiān)測,精準防治災(zāi)害
加大終端監(jiān)測技術(shù)投資,拓寬實時監(jiān)測范圍,確保監(jiān)測的準確全面性。提高對農(nóng)作物種植面積、自然災(zāi)害強度、天氣情況、氣溫條件的因素監(jiān)控。通過實時的監(jiān)測結(jié)果,把握時效,科學(xué)有效的提供防治措施,精準幫助農(nóng)業(yè)生產(chǎn)者積極規(guī)避各類風(fēng)險。從而在提高農(nóng)業(yè)生產(chǎn)者對農(nóng)險產(chǎn)品的認可度的同時,幫助其減少農(nóng)業(yè)生產(chǎn)過程中的風(fēng)險損失。
2.結(jié)語
農(nóng)業(yè)保險還存在很大的創(chuàng)新空間,未來會繼續(xù)做大做強。雖然目前我國的農(nóng)業(yè)保險種類多、覆蓋面廣,但仍存在深度和密度不夠、農(nóng)業(yè)保險參與農(nóng)村治理的作用體現(xiàn)不明顯、農(nóng)民投保積極性不夠高、產(chǎn)品創(chuàng)新性還有待繼續(xù)加強等問題。本文通過對農(nóng)險購買者信息特征化,輔助用戶畫像模型將農(nóng)險用戶進行個性化分類,致力于實現(xiàn)農(nóng)業(yè)保險的精準營銷,推動農(nóng)業(yè)保險的健康快速發(fā)展,實現(xiàn)農(nóng)業(yè)的高質(zhì)量發(fā)展。
參考文獻:
[1]郝勝宇,陳靜仁.大數(shù)據(jù)時代用戶畫像助力企業(yè)實現(xiàn)精準化營銷[J].中國集體經(jīng)濟,2016(04):61-62.
[2]郭光明.基于社交大數(shù)據(jù)的用戶信用畫像方法研究[D].中國科學(xué)技術(shù)大學(xué),2017.
[3]程靜,劉飛,陶建平.風(fēng)險認知、風(fēng)險管理與農(nóng)險需求——基于行為金融視角的實證研究[J].南京農(nóng)業(yè)大學(xué)學(xué)報(社會科學(xué)版),2018,18(03):133-141+156.
[4]干潔.紹興市水稻種植農(nóng)戶參保行為及其影響因素研究[D].浙江農(nóng)林大學(xué),2019.
[5]劉穎.大數(shù)據(jù)技術(shù)的圖書館用戶畫像模型建構(gòu)策略[J].現(xiàn)代電子技術(shù),2020,43(09):55-57+61.
[6]王阿星,張峭.內(nèi)蒙古鄂爾多斯市農(nóng)業(yè)保險需求實證分析[J].農(nóng)業(yè)經(jīng)濟問題,2008(S1):101-106.
[7]User20Profiling20for20University20Recommender20System20Using20Automatic20Information20Retrieval[J].Sumitkumar Kanoje,Debajyoti Mukhopadhyay,Sheetal Girase. Procedia Computer Science.2016.
[8]Automatic20Generation20of20Personas20Using20YouTube20Social20Media20Data.An J,Kwak H,Jansen B J. Proceedings of the50th International Conference on System Sciences.2017.
[9]Factors Influencing Farmers Crop Insurance Decisions.Bruce J Sherrick. American Journal of Agricultural Economics.2004.
[10]Agricultural Insurance in Developed Countries: Where Have We Been and Where Are We Going?[J].Vincent H.Smith,Joseph W.Glauber.Applied Economic Perspectives and Policy.2012 (3).
[11]Adoption of weather‐index insurance: learning from willingness to pay among a panel of households in rural Ethiopia[J].Ruth Vargas Hill,John Hoddinott,Neha Kumar.Agricultural Economics.2013 (4-5).
[12]Low prices and crop insurance[J]. Anderson,Kim. Southwest Farm Press. 2015.
基金項目:本文屬安徽財經(jīng)大學(xué)國家級創(chuàng)新訓(xùn)練項目《基于用戶畫像模型的農(nóng)業(yè)保險精準營銷研究——以安徽省為例》(編號:201910378033)研究成果。
作者簡介:周紫薇(1999-),女,漢族,安徽銅陵人,單位:安徽財經(jīng)大學(xué)金融學(xué)院,學(xué)歷:金融工程專業(yè)。
