黑曲霉木聚糖酶的同源表達(dá)及其高產(chǎn)木聚糖酶發(fā)酵條件的優(yōu)化*
2015-12-25 01:59:08倪大偉李瑞云白愛枝張文濤王軍
食品與發(fā)酵工業(yè) 2015年4期
倪大偉,李瑞云,白愛枝,張文濤,王軍
1(內(nèi)蒙古自治區(qū)離子束生物工程重點實驗室,內(nèi)蒙古呼和浩特,010021)
2(內(nèi)蒙古大學(xué)生命科學(xué)學(xué)院,內(nèi)蒙古呼和浩特,010021)3(內(nèi)蒙古大學(xué) 物理科學(xué)與技術(shù)學(xué)院,內(nèi)蒙古呼和浩特,010021)
黑曲霉(Aspergillus niger)具有多種活性較強的酶系,是重要的酶制劑生產(chǎn)菌種,也是木聚糖酶的生產(chǎn)菌種之一。隨著真菌基因組學(xué)和基因微陣列及蛋白質(zhì)組學(xué)等實驗技術(shù)的發(fā)展,利用絲狀真菌作為宿主表達(dá)同源和異源蛋白也得到快速發(fā)展和應(yīng)用[1],黑曲霉作為絲狀真菌的代表性菌種之一,其表達(dá)系統(tǒng)也越來越受到科技工作者的關(guān)注[2-3]。黑曲霉是國際公認(rèn)的安全生產(chǎn)菌株[4],并且其本身具有強大的蛋白質(zhì)胞外分泌能力,有完善的翻譯后加工功能及成熟的發(fā)酵工藝等優(yōu)點,是外源和內(nèi)源蛋白表達(dá)的優(yōu)良宿主。
木聚糖酶是一種重要的工業(yè)酶制劑,在食品、飲料、造紙和飼料等方面有廣泛的應(yīng)用。目前市場上大部分高效木聚糖酶產(chǎn)品的技術(shù)核心都屬于國外公司,因此通過基因工程技術(shù),構(gòu)建高表達(dá)的木聚糖酶工程菌,對于降低木聚糖酶的價格促進其應(yīng)用水平,是國內(nèi)木聚糖酶工業(yè)所面臨的最緊迫的問題。黑曲霉產(chǎn)木聚糖酶基因xynB的同源表達(dá)研究的甚少,2005年Anthony等[5]利用 RT-PCR 從 A.niger BRFM281 mRNA擴增的帶有6個his標(biāo)簽的xynB基因克隆到有g(shù)pd強啟動子的表達(dá)盒,將該表達(dá)盒在蛋白酶陰性突變株Aspergillus niger D15#26中進行表達(dá),獲得的1株基因工程菌的木聚糖酶產(chǎn)量達(dá)到900 mg/L,酶活力為625 U/mL;國內(nèi)的鄭瑞娟[6]從A.niger TS1中利用同樣方法克隆得到xynB基因,通過構(gòu)建與葡萄糖淀粉酶的融合表達(dá)質(zhì)粒,在尿嘧啶缺陷型A.niger M54中表達(dá),獲得的重組菌的酶活力最高達(dá)507 IU/mL,分別是原出發(fā)菌和宿主菌的6.7倍和3.89倍。Record等[7]利用 Aspergillus nidulans gpd基因的啟動子、5’非編碼區(qū)和終止子將A.niger來源的阿魏酸脂酶faeA基因進行同源表達(dá),表達(dá)量較原始菌株提高24.5 倍,達(dá) 1 g/L;Anthony Levasseur[8]采用同樣的策略表達(dá)A.niger BRFM131的阿魏酸脂酶faeB基因,產(chǎn)量較原菌株提高了16倍;同源表達(dá)較之異源表達(dá),可有效避免異源表達(dá)中難以解決的問題,如分泌過程中蛋白的錯誤折疊和被內(nèi)源蛋白酶降解的可能以及物種密碼子的偏好性、基因結(jié)構(gòu)的復(fù)雜性等,多種蛋白酶同源表達(dá)的成功,表明該策略具有廣泛的應(yīng)用價值。本實驗室從A.niger A327中克隆了木聚糖酶基因xynB[9],采用同源表達(dá)策略將其表達(dá)在黑曲霉菌株A.niger A327中,本文對木聚糖酶活力最高的轉(zhuǎn)化子A.niger A70菌株的搖瓶發(fā)酵條件進行了優(yōu)化。
1 材料與方法
1.1 菌株與載體
出發(fā)菌株黑曲霉A.niger A327,由內(nèi)蒙古自治區(qū)離子束生物工程重點實驗室保藏;質(zhì)粒 VHb和p3SR2由中國科學(xué)院微生物所唐國敏教授惠贈。
1.2 主要儀器與試劑
TU-1800PC紫外-可見分光光度計,北京譜析通用儀器設(shè)備有限責(zé)任公司;DYY-12型電泳儀,北京六一;ZHWY-1102/1121型普通搖床,上海智誠,主要試劑:木聚糖(birchwood xylan),美國sigma公司;玉米芯麩皮購于呼和浩特市郊,玉米芯經(jīng)粉碎機粉碎后備用;其他化學(xué)試劑均為國產(chǎn)分析純。
1.3 表達(dá)載體的構(gòu)建及其表達(dá)
從A.niger A327中克隆編碼xynB結(jié)構(gòu)基因及其5’調(diào)控區(qū)序列片段,總長1611bp,將其與目的載體pBS-T 連接[5],并轉(zhuǎn)化 E.coli DH5α,挑取單克隆,鑒定陽性克隆子,分別將從質(zhì)粒VHb和質(zhì)粒p3SR2酶切得到的來源于A.nidulans的終止子TtrpC和選擇標(biāo)記基因amdS連接到已連入xynB基因的T載體中,得到表達(dá)載體pBS-XTP(圖12,并利用原生質(zhì)體轉(zhuǎn)化技術(shù)將表達(dá)載體轉(zhuǎn)入菌株A327中,通過搖瓶發(fā)酵篩選產(chǎn)木聚糖酶活力最高的轉(zhuǎn)化子。
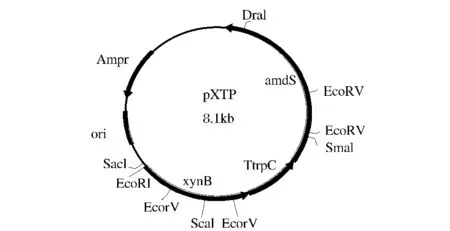
圖1Fig.1
1.4 木聚糖酶活力的測定方法[10]
取適當(dāng)稀釋的粗酶液0.1 mL,加到1 mL、1%的樺木木聚糖懸浮液中(pH5.0,0.2 mol/L醋酸緩沖液配制)于50℃水浴反應(yīng)10 min,用 DNS(3,5-二硝基水楊酸)法,測定產(chǎn)生的還原糖。以木糖為標(biāo)準(zhǔn),在本實驗條件下,每分鐘產(chǎn)生1 μmol還原糖所需的酶量定義為1個酶活力單位U/mL。
1.5 培養(yǎng)基和培養(yǎng)方法
分離培養(yǎng)基(g/L):麩皮5,蛋白胨1,瓊脂20,Manders營養(yǎng)鹽液[11]定容 1 000 mL,pH 自然。PDA培養(yǎng)基(g/L):馬鈴薯200(去皮、切塊煮30 min,紗布過濾),葡萄糖20,瓊脂15,蒸餾水定容至1 000 mL,pH自然。基礎(chǔ)產(chǎn)酶培養(yǎng)基(g/L):玉米芯75,麩皮24,Manders營養(yǎng)鹽液 1 000 mL,pH5.5 ~6.0。以上培養(yǎng)基在沒有添加糖蜜的情況下均在121℃滅菌30 min,添加糖蜜的培養(yǎng)基在115℃滅菌30 min;固體培養(yǎng),置于30℃下培養(yǎng)4~5 d;液體培養(yǎng),30℃下,搖瓶培養(yǎng)發(fā)酵4 d,搖床轉(zhuǎn)速為200 r/min。
1.6 基因工程菌產(chǎn)木聚糖酶條件優(yōu)化
1.6.1 培養(yǎng)基的單因素優(yōu)化
在初始發(fā)酵培養(yǎng)基的基礎(chǔ)上,保持其他條件不變,分別添加不同濃度的玉米芯(40,50,60,70,80,90 g/L),不同濃度的麩皮(15,20,25,30 g/L),不同濃度的糖蜜(0,2,4,6,8,10 g/L)以及濃度均為 6 g/L的不同氮源((NH4)2SO4、NaNO3、NH4NO3、蛋白胨、尿素),進行搖瓶發(fā)酵,研究各因子對木聚糖酶酶活的影響。
1.6.2 發(fā)酵條件優(yōu)化
在基礎(chǔ)產(chǎn)酶培養(yǎng)基的條件下,分別設(shè)置不同溫度(25℃(室溫)、28℃(黑曲霉A327產(chǎn)酶最適生長溫度)、31 ℃、34 ℃),接種量(1.0、1.5、2.0、2.5 mL,孢子懸液濃度 108個/mL),裝液量(30、45、60 mL)和搖床轉(zhuǎn)速(180、200、220轉(zhuǎn)/min)對基因工程菌A70的發(fā)酵條件進行優(yōu)化。
1.6.3 培養(yǎng)基的正交試驗
設(shè)計了4因素3水平的L9(34)正交試驗(見表1),進一步對營養(yǎng)條件進行優(yōu)化。

表1 正交設(shè)計方案 單位:g/LTable1 Orthogonal design project
1.6.4 最適發(fā)酵條件驗證
利用上述單因素和正交試驗獲得的最適發(fā)酵條件和培養(yǎng)基組成進行最適發(fā)酵條件的驗證實驗。
以上發(fā)酵試驗均設(shè)置3組平行試驗,取平均水平,以發(fā)酵96 h上清液的木聚糖酶活力作為考核指標(biāo)考察各因素對基因工程菌A70產(chǎn)木聚糖酶的影響。
2 結(jié)果與分析
2.1 xynB基因在A.niger A327中的同源表達(dá)
將出發(fā)菌株A327菌株和篩選的部分轉(zhuǎn)化子(包括產(chǎn)酶水平最高的A70轉(zhuǎn)化子)分別在100 mL搖瓶中,28℃,200 r/min搖床發(fā)酵培養(yǎng),將發(fā)酵上清液離心得到粗酶提取液,沸水煮沸5 min,取相同體積的粗酶液進行SDS-PAGE(結(jié)果見圖2)。可見菌株A37,A55,A70,A74,A75,A77,A78 均不同程度地表達(dá)了xynB基因,木聚糖酶的產(chǎn)量不同程度的有所提高。目的產(chǎn)物分子量約為21 kDa,從圖中可以看出A63菌株產(chǎn)木聚糖酶的量較對照還要低,而A70菌株分泌木聚糖酶的量最多,這與發(fā)酵后酶活測定的結(jié)果一致(見表2)。表達(dá)程度的高低與xynB基因插入的拷貝數(shù)有關(guān),在一定范圍內(nèi)插入拷貝數(shù)越多,表達(dá)量越高,當(dāng)拷貝數(shù)進一步提高,或許由于調(diào)控蛋白的滴定效應(yīng),表達(dá)不再上升[12],這一基因劑量效應(yīng)在外源蛋白的表達(dá)中同樣成立。從圖中可以看出有的菌株在表達(dá)xynB基因的同時,使其他蛋白的分泌有所提高,這可能與xynB基因所插入的位置有關(guān),正好使其他蛋白的表達(dá)受到調(diào)控,提高了產(chǎn)量。

圖2 轉(zhuǎn)化株產(chǎn)酶的SDS-PAGE分析Fig.2 The SDS-PAGE detection of xylanase transformants

表2 轉(zhuǎn)化株發(fā)酵的木聚糖酶活力(IU/mL)Table 2 Xylanase activity of transformants fermentation
2.2 高效表達(dá)菌株A70的遺傳穩(wěn)定性
基因工程菌的遺傳不穩(wěn)定性主要表現(xiàn)在重組質(zhì)粒的不穩(wěn)定性,這種不穩(wěn)定性具有下列兩種表現(xiàn)形式:其一是結(jié)構(gòu)不穩(wěn)定性,重組 DNA分子上某一區(qū)域發(fā)生缺失、重排、修飾,導(dǎo)致其表觀生物學(xué)功能的喪失;其二是分配不穩(wěn)定性,整個重組 DNA分子從受體細(xì)胞中逃逸。將A70菌株進行斜面到斜面的傳代培養(yǎng),傳代6次后,在基礎(chǔ)產(chǎn)酶培養(yǎng)基和培養(yǎng)條件下,將經(jīng)不同傳代次數(shù)的A70菌株同時進行發(fā)酵實驗,結(jié)果(見表3)顯示,該菌株的遺傳穩(wěn)定性較好。

表3 Strain A70菌株的傳代酶活測定Table 3 The enzyme activity from continual inoculation of A70
2.3 發(fā)酵時間對菌株A70產(chǎn)酶的影響
隨著發(fā)酵時間的延長,微生物的代謝產(chǎn)物隨之增加,但分泌的代謝產(chǎn)物量通常受到營養(yǎng)條件、培養(yǎng)條件等多種因素的影響。實驗采取在相同的培養(yǎng)條件下,利用基礎(chǔ)產(chǎn)酶培養(yǎng)基對出發(fā)菌株A327和基因工程菌A70的發(fā)酵特性進行比較。結(jié)果如圖3和圖4所示,可見發(fā)酵96 h基因工程菌A70達(dá)到產(chǎn)酶高峰期,較出發(fā)菌株A327的產(chǎn)酶周期延長24 h,這可能是由于其基因序列中插入了表達(dá)載體,使得菌體的生長變慢或者使菌體的代謝調(diào)控機制受到影響,而使產(chǎn)酶周期延長,工程菌A70產(chǎn)酶水平為360 IU/mL,較出發(fā)菌株A327提高約16倍。

圖3 黑曲霉菌株A327的產(chǎn)酶曲線Fig.3 The cure of Strain A327 on xylanase production
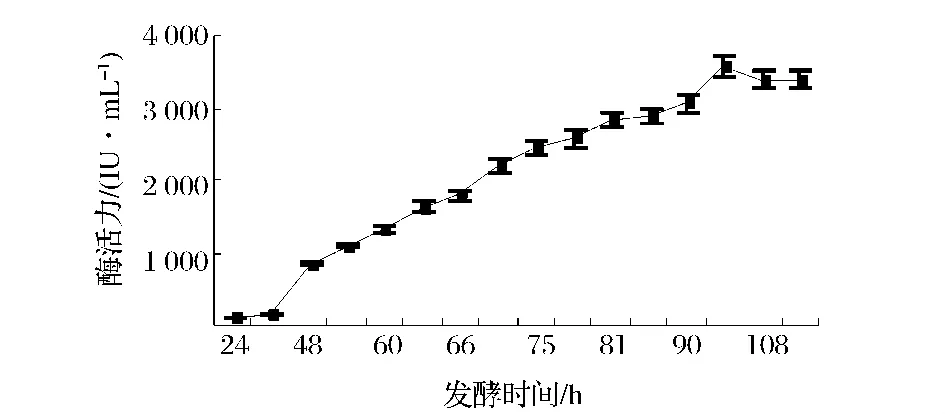
圖4 黑曲霉菌株A70的產(chǎn)酶曲線Fig.4 The curve of A70 on xylanase production
2.4 高表達(dá)菌株發(fā)酵條件優(yōu)化
在基礎(chǔ)產(chǎn)酶培養(yǎng)基條件下,根據(jù)實驗的目的添加或改變相應(yīng)的條件,對產(chǎn)酶水平最高的基因工程菌A70的發(fā)酵條件進行優(yōu)化。
2.4.1 碳源濃度對產(chǎn)酶的影響
在前期的研究中發(fā)現(xiàn)出發(fā)菌株A327以玉米芯為碳源時產(chǎn)木聚糖酶活力最高[13],因此以玉米芯為主要碳源考察其初始發(fā)酵濃度對基因工程菌株A70產(chǎn)木聚糖酶的影響,結(jié)果見圖5,90 g/L的起始玉米芯濃度產(chǎn)酶的活力最高,同時也發(fā)現(xiàn)該濃度下發(fā)酵液因失水較多而變得粘稠,其次是80,40 g/L稍次之。

圖5 玉米芯的量對產(chǎn)酶的影響Fig.5 Effect of corncob concentration on xylanase production
2.4.2 麩皮質(zhì)量濃度對產(chǎn)酶的影響
本試驗在2.3.1的基礎(chǔ)上,在其它條件不變的情況下,分別以玉米芯質(zhì)量濃度為40、80和90 g/L,添加不同比例的麩皮進行發(fā)酵研究,發(fā)現(xiàn)當(dāng)玉米芯濃度和麩皮濃度為80 g/L和15 g/L時菌株A70產(chǎn)木聚糖酶的活力最高(結(jié)果見表4)。
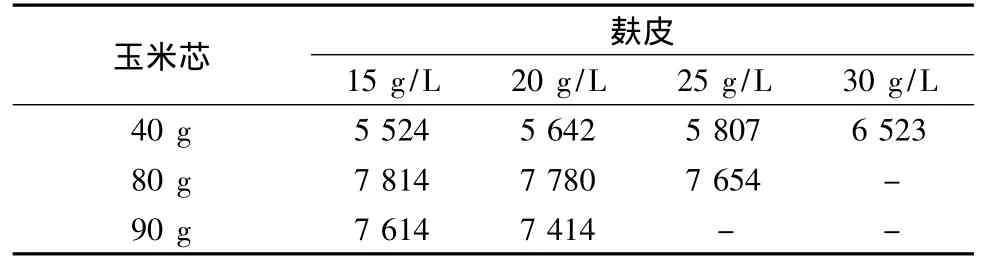
表4 麩皮質(zhì)量濃度對產(chǎn)木聚糖酶活力的影響 單位:IU/mLTable 4 Effect of bran concentration on xylanase production
2.4.3 附加碳源濃度對產(chǎn)酶的影響
通常微生物發(fā)酵所需的碳源不止一種,而且不同種類的碳源對微生物代謝產(chǎn)物的分泌都有較大的影響。在其它條件不變的情況下,以玉米芯濃度為80 g/L,麩皮濃度為15 g/L,添加不同的速效碳源淀粉、葡萄糖、蔗糖、甜菜糖蜜、麥芽糖,發(fā)現(xiàn)菌株對甜菜糖蜜的利用效果最好。糖蜜是制糖工業(yè)的副產(chǎn)品,其組成因制糖原料、加工條件的不同而有差異,因其含有大量的蔗糖,因此是很好的微生物發(fā)酵培養(yǎng)基組分。對菌株A70來說,其利用甜菜糖蜜發(fā)酵產(chǎn)酶效果較單純的蔗糖好的原因可能是糖蜜中還含有較多的礦物質(zhì)成分以及其他營養(yǎng)素,對菌株的生長更有利。通過添加不同濃度的糖蜜考察其對菌株A70產(chǎn)木聚糖酶的影響,發(fā)現(xiàn)當(dāng)添加的糖蜜濃度為12 g/L時,菌株產(chǎn)木聚糖酶的活力最高(見圖6)。
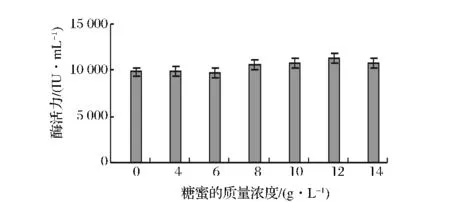
圖6 糖蜜質(zhì)量濃度對產(chǎn)酶的影響Fig.6 Effect of molasses concentration on xylanase production
2.4.4 不同氮源對產(chǎn)酶的影響
在上述實驗結(jié)果的基礎(chǔ)上,培養(yǎng)基以80 g/L玉米芯,15 g/L麩皮,Mandels鹽液中去除(NH4)2SO4和 NaNO3后,分別加入(NH4)2SO4、NaNO3、NH4NO3、蛋白胨、尿素,質(zhì)量濃度為6 g/L,對照為原Mandlels鹽液不變,來研究氮源對產(chǎn)木聚糖酶的影響,結(jié)果見圖7,可見菌株A70對氮源的利用以尿素的產(chǎn)酶效果最好,NaNO3次之;但是實驗發(fā)現(xiàn)以尿素為氮源時,到發(fā)酵終點培養(yǎng)基的失水較嚴(yán)重,相比之下NaNO3的發(fā)酵效果更好。

圖7 不同氮源對產(chǎn)酶的影響Fig.7 Effect of different nitriogen source on xylanase production
2.5 發(fā)酵條件對產(chǎn)酶的影響
以基礎(chǔ)產(chǎn)酶培養(yǎng)基對菌株A70的發(fā)酵溫度、接種量及溶氧條件進行了研究,試驗表明,產(chǎn)酶的最適溫度為28~30℃(見表5),最適接種量為1.5 mL/瓶(見表6),最適裝液量為30 mL/瓶,當(dāng)搖床轉(zhuǎn)速逐漸從180 r/min增加到220 r/min時,菌株的產(chǎn)酶水平隨之提高,說明菌株對溶氧的需求較高(見表7),這也符合霉菌生長的特點。
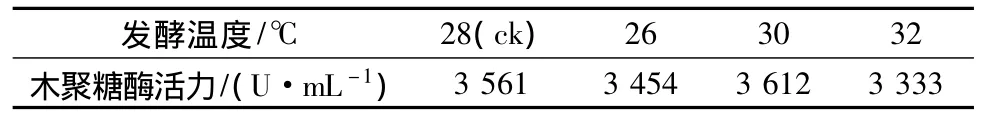
表5 不同發(fā)酵溫度對酶活的影響Table 5 The effect of different fermentation temperature

表6 接種量對菌株發(fā)酵產(chǎn)酶的影響Table 6 The effect of inoculation quantity on xylanase production

表7 溶氧對發(fā)酵的影響Table 7 The effects of ventilation in fermentation
2.6 發(fā)酵產(chǎn)酶培養(yǎng)基的正交試驗
上述實驗僅考慮了各個因素單獨的作用,未考慮各個因素均發(fā)生變化時,高產(chǎn)菌株發(fā)酵水平的變化。為了獲得同源表達(dá)基因工程菌黑曲霉A70最佳的產(chǎn)酶營養(yǎng)條件,選用L9(34)表設(shè)計了4因素3水平的正交試驗(見表1)進一步對營養(yǎng)條件進行優(yōu)化。正交試驗結(jié)果見表8。
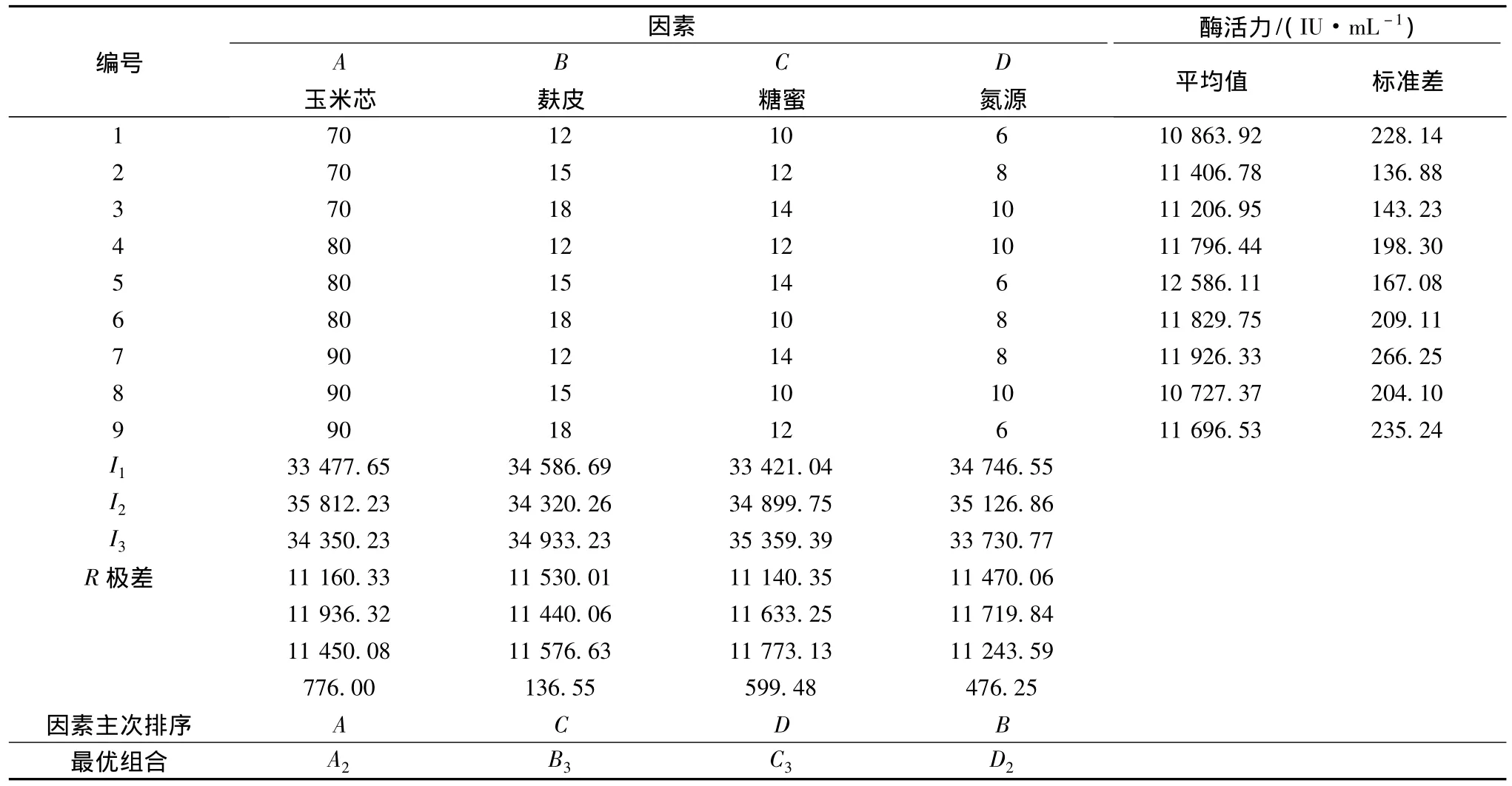
表8 正交實驗結(jié)果Table 2 Results of orthogonal test design
通過直觀分析正交試驗的結(jié)果,可以看出影響基因工程菌A70產(chǎn)酶的各因素主次順序為A>C>D>B,最優(yōu)組合為玉米芯80 g/L,麩皮18 g/L,糖蜜14 g/L,NaNO38 g/L,在該組合下菌株產(chǎn)酶的最高水平為12586 U/mL。
2.7 最佳工藝條件驗證
按2.5確定的最佳培養(yǎng)基配方和2.4確定的最適發(fā)酵條件進行3次平行實驗,平均產(chǎn)酶水平為12 639 U/mL(見表9),實驗結(jié)果表明,發(fā)酵工藝的重現(xiàn)性好,菌株的產(chǎn)酶水平穩(wěn)定。

表9 最適發(fā)酵條件驗證Table 9 The validation of optimal fermentation conditions
3 結(jié)論
通過對黑曲霉同源表達(dá)高產(chǎn)菌株培養(yǎng)基及發(fā)酵條件的優(yōu)化,菌株的發(fā)酵周期為4天,發(fā)酵培養(yǎng)基以玉米芯80 g/L,麩皮18 g/L,糖蜜14 g/L,NaNO38g/L,30 mL無氮Mandels營養(yǎng)液為最適培養(yǎng)基配方。發(fā)酵溫度為28~30℃,接種量為1.5 mL/瓶,搖床轉(zhuǎn)速以220 r/min發(fā)酵產(chǎn)酶水平最高,在上述優(yōu)化的條件下基因工程菌A70產(chǎn)酶水平為12 664 U/mL,較優(yōu)化前的3 601 U/mL提高約3.5倍。同源表達(dá)木聚糖酶的黑曲霉菌株A70發(fā)酵產(chǎn)酶水平較高,生產(chǎn)成本低,該實驗為用黑曲霉表達(dá)系統(tǒng)高效表達(dá)其他酶類提供了參考。
[1] Owen P.Ward.Production of recombinant proteins by filamentous fungi[J].Biotechnology Advances,2012,1119-1139.
[2] Roth AH.Dersch.P.A novel expression system for intracellular production and purification of recombinant affinitytagged proteins in Aspergillus niger[J].Appl Microbiol Biotechnol(2010)86:659-670
[3] Amaro-Reyes A,Garci'a-Almenda'rez BE,Va'zquez-Mandujano DG,et al.Homologue expression of a b-xylosidase from native Aspergillus niger[J].J Ind Microbiol Biotechnol,2011(38):1311-1319.
[4] Punt PJ,VAN Biezen N,Conesa A,et al.Filamentous fungi as cell factories for heterologous protein production.Trends in Biotechnology,2001,20:200-6.
[5] Levasseur A,Asther M,Record E.Overproduction and characterization of xylanase B from Aspergillus niger.Can J Microbiol,2005,51:177-183
[6] 鄭瑞娟.黑曲霉木聚糖酶在同源宿主中的分泌性表達(dá)及酶學(xué)特性研究[D].上海:同濟大學(xué)碩士學(xué)位論文,2006.
[7] Record E,Asther M,Sigoillot C,et al.Overproduction of the Aspergillus niger feruloyl esterase for pulp Bleaching application.Appl Microbiol Bioteehnol.20O3(62):349-355
[8] Levasseur A,Benoit I,Astger M,et al.Homologous expression of the feruloyl esterase B gene from Aspergillus niger and characterization of the recombinant enzyme..Protein Expression and purifieation.2004(37):126-133.
[9] 白愛枝,閆祖威,唐國敏,等.黑曲霉木聚糖酶結(jié)構(gòu)基因和5'調(diào)控區(qū)基因克隆及其分析[J].華北農(nóng)學(xué)報,2009,24(5):73-76.
[10] Bailey MJ,Biely P,Poutanen KJ.Interlaboratory testing of methods for assay of xylanase[J].J Biotechnol,1992,23:257-270
[11] Mandels M,Weber J.The production of cellulase[J].Adv Chem Ser,1969(95):391-413
[12] Verdoes J C,Punt P J,Stouthamer A H,et al.,The effect of multiple copies of the upstream region on expression of the Aspergillus niger glucoamylase-encoding gene[J].Gene,1994,145(2):179-87.
[13] 白愛枝,鮑秀珍,潘仁瑞.碳源和氮源對黑曲霉產(chǎn)木聚糖酶的影響[J].飼料工業(yè),2006,27(2):17-20.
