放牧對草地土壤微生物的影響
2016-01-28 08:09:29茍燕妮南志標(biāo)
草業(yè)學(xué)報 2015年10期
茍燕妮,南志標(biāo)
(草地農(nóng)業(yè)生態(tài)系統(tǒng)國家重點實驗室,蘭州大學(xué)草地農(nóng)業(yè)科技學(xué)院,甘肅 蘭州 730020)
放牧對草地土壤微生物的影響
茍燕妮,南志標(biāo)*
(草地農(nóng)業(yè)生態(tài)系統(tǒng)國家重點實驗室,蘭州大學(xué)草地農(nóng)業(yè)科技學(xué)院,甘肅 蘭州 730020)
摘要:土壤質(zhì)量是草地生態(tài)系統(tǒng)生產(chǎn)力可持續(xù)發(fā)展的關(guān)鍵所在。健康的土壤必定擁有豐富的物種多樣性、活躍的生命體、高效的內(nèi)部養(yǎng)分循環(huán)及較好的抗干擾能力。土壤微生物作為土壤系統(tǒng)中各種生物進(jìn)程的主要推動者,其作用不可小覷。因此,我們可以利用土壤微生物的數(shù)量、群落結(jié)構(gòu)和多樣性等特征監(jiān)測土壤質(zhì)量。本文綜述了放牧對草地土壤微生物數(shù)量、生物量、群落結(jié)構(gòu)和多樣性的影響。指出通過了解土壤微生物與放牧行為間的關(guān)系,如何評估土壤質(zhì)量,選取適宜的放牧梯度,完善草地管理模式,并介紹了土壤微生物群落結(jié)構(gòu)和多樣性研究方法的利弊。解決兩個關(guān)鍵問題:1)土壤微生物的各種特征是如何響應(yīng)放牧行為的。2)如何通過這些指標(biāo)評價土壤質(zhì)量,然后提出合理的放牧模式,改善和維護(hù)草地的生產(chǎn)力。
關(guān)鍵詞:放牧;土壤;微生物生物量;微生物多樣性
The impacts of grazing on the soil microorganism population of grassland
GOU Yan-Ni, NAN Zhi-Biao*
StateKeyLaboratoryofGrasslandAgro-ecosystems,CollegeofPastoralAgriculturalScienceandTechnology,LanzhouUniversity,Lanzhou730020,China
Abstract:Soil quality is crucial to sustainable development of grassland ecosystems and to their productivity. Healthy soil must have rich species diversity, active microorganism populations, an efficient internal nutrient cycle and good buffering against disturbance. Soil microorganisms are the main driver of various biological processes and are therefore very important in soil systems. Hence, soil microbial community characteristics such as population numbers, community structure and species diversity can be used as a measure of soil health. The present paper summarizes the effects of grazing on the grassland soil microbial populations, community structure and diversity. We propose that it is essential to understand the impact of grazing management on soil microorganisms. A methodology is introduced for evaluation of soil health, and selection of a suitable grazing level to improve grassland management, and the advantages and disadvantages are discussed. Two key questions are addressed; namely the responses of the soil microbial community to grazing activity, and how to use these indicators to evaluate soil quality and propose a reasonable grazing practice to sustainably improve the productivity of grassland.
Key words:grazing; soil; microbial biomass; microbial diversity
放牧是調(diào)節(jié)草地生產(chǎn)力最經(jīng)濟有效的管理方式[1-2],對草地生態(tài)系統(tǒng)地上和地下均有影響[3-4]。有蹄類動物通過加強土壤微生物營養(yǎng)富集和根際作用,提高礦物質(zhì)有效利用率,最終促進(jìn)植物營養(yǎng)能量的流動和光合作用[5]。土壤微生物作為草地地下生態(tài)系統(tǒng)最大的資源庫,推動整個環(huán)境中物質(zhì)和能量的流動和轉(zhuǎn)移,對維持草地生態(tài)系統(tǒng)有重要的作用[6]。土壤微生物對土壤質(zhì)量變化敏感,尤其是土壤微生物多樣性作為土壤生物學(xué)性質(zhì)的敏感指標(biāo),能夠較早地指示草原生態(tài)環(huán)境變化和生態(tài)系統(tǒng)功能的變化[7]。研究掌握土壤微生物對放牧強度的響應(yīng)在草原生態(tài)系統(tǒng)保護(hù)、恢復(fù)及重建中具有重要理論和實踐意義。
因此,為了明確放牧與土壤微生物群落結(jié)構(gòu)、營養(yǎng)物質(zhì)循環(huán)和分解過程之間的重要關(guān)系,關(guān)于放牧對土壤微生物群落影響的研究越來越多,主要有以下幾個方面:土壤微生物數(shù)量、生物量、活性、群落結(jié)構(gòu)和多樣性的影響。研究表明,家畜的采食、踩踏和糞尿回地都會對土壤生物區(qū)系帶來影響[8-11]。而且隨著放牧率的增大,營養(yǎng)循環(huán)加速、凋落物生物量和土壤微生物生物量均增多。但是重度放牧?xí)耐寥蕾Y源,從而減少地上和地下的生物量[12-13]。隨著研究的深入,學(xué)者們發(fā)現(xiàn),由于不同的地理位置和季節(jié)氣候下土壤微生物對放牧的響應(yīng)方式不一致[14-16],因此,放牧與土壤微生物間關(guān)系復(fù)雜,而且又受環(huán)境因素的影響而呈現(xiàn)多變的變化趨勢。所以本文對不同環(huán)境條件下放牧與土壤微生物間的關(guān)系做一綜述,以期為以后的研究提供一些可參考的信息和依據(jù)。
1放牧對草地微生物數(shù)量的影響
土壤微生物是陸地生態(tài)系統(tǒng)中非常龐大的一個資源庫。每克土壤含有幾百萬個微生物[17],2000~18000種細(xì)菌[18]。近年來,Gans等[17]運用現(xiàn)代DNA測序技術(shù)研究得出,每克土壤里約有107種微生物。土壤微生物數(shù)量是土壤微生物最基礎(chǔ)的研究對象,有些研究者在分析土壤微生物時通常會先對微生物的數(shù)量和分布變化情況進(jìn)行數(shù)據(jù)采集,現(xiàn)今,最為常見的計數(shù)方法為平板表面涂抹計數(shù)法。
目前,土壤微生物主要研究對象為3大類群:細(xì)菌、真菌和放線菌。不同類型草地各放牧強度下三大類群微生物的數(shù)量變化趨勢由表1呈現(xiàn)。趙吉[19]對內(nèi)蒙古冷蒿(Artemisiafrigida)小禾草草原進(jìn)行4年的輪牧試驗發(fā)現(xiàn),土壤各微生物類群數(shù)量均隨放牧率的增大而減少,重牧區(qū)的微生物總數(shù)下降約30%。尤其是自由放牧區(qū)的微生物數(shù)量顯著低于放牧試驗區(qū)。他們認(rèn)為過度放牧不利于土壤微生物的生長,并且發(fā)現(xiàn)微生物數(shù)量與土壤有機碳含量顯著正相關(guān)。張成霞和南志標(biāo)[20]對隴東黃土高原天然草地土壤微生物和土壤理化性質(zhì)相關(guān)性進(jìn)行研究發(fā)現(xiàn):土壤微生物總數(shù)和細(xì)菌數(shù)量隨放牧強度的增大呈先減少后增多的趨勢,而真菌和放線菌數(shù)量隨放牧強度的增大而減少。土壤微生物總數(shù)在重牧條件下最多,這是由細(xì)菌數(shù)量最多引起的。因為,不同草地類型和利用方式下,與真菌和放線菌相比,細(xì)菌占有絕對優(yōu)勢,所以,土壤微生物數(shù)量主要取決于細(xì)菌數(shù)量[21]。對于細(xì)菌數(shù)量在重牧地出現(xiàn)最高值這一現(xiàn)象,張成霞和南志標(biāo)[20]認(rèn)為是由于牲畜的踐踏使重牧地土壤變得緊實,通氣性下降,從而使嫌氣性細(xì)菌數(shù)量略有增加,再加上細(xì)菌適應(yīng)不良環(huán)境的能力強,因而導(dǎo)致這種現(xiàn)象的發(fā)生。關(guān)于土壤微生物與土壤理化性質(zhì)間相關(guān)性結(jié)果[22]表明,細(xì)菌數(shù)量與銨態(tài)氮、硝態(tài)氮及pH值呈正相關(guān),與全氮和速效磷呈負(fù)相關(guān)。劉世貴等[22]對川西北退化草甸草地土壤微生物主要生理類群數(shù)量進(jìn)行研究發(fā)現(xiàn),土壤微生物總數(shù)隨草地退化程度的加深而減少,而且3種退化草地中嫌氣性纖維素分解細(xì)菌數(shù)量最多,這為重牧地細(xì)菌數(shù)量最多提供了參考依據(jù)。李春莉等[23]對內(nèi)蒙古短花針茅(Stipabreviflora)草原研究顯示:土壤細(xì)菌、真菌和放線菌數(shù)量隨放牧強度變化趨勢均為:重牧地<不放牧<輕牧地,表明適度的放牧率會引起土壤微生物數(shù)量增多,而不放牧和過度放牧不利于土壤微生物的生長。土壤微生物與土壤有機質(zhì)和土壤全氮呈較強的正相關(guān)關(guān)系。單貴蓮等[24]對內(nèi)蒙古半干旱典型圍封草原研究發(fā)現(xiàn),圍封草地與自由放牧地相比,細(xì)菌、真菌和放線菌數(shù)量無顯著增加,且隨圍封年限的延長呈遞增趨勢,但當(dāng)圍封年限達(dá)到20年時,細(xì)菌數(shù)量顯著增多,而真菌和放線菌數(shù)量有所下降。顧愛星等[25]對天山北坡溫帶極端干旱天然草地研究發(fā)現(xiàn),退化草地的微生物數(shù)量高于未退化草地,且重度退化草地的微生物數(shù)量最多,土壤微生物數(shù)量隨草地退化程度的加深呈現(xiàn)先增加后減少的關(guān)系。同時根據(jù)Wang和Sheng[26]關(guān)于松嫩平原農(nóng)牧交錯帶天然草地的研究發(fā)現(xiàn),土壤細(xì)菌、放線菌和真菌數(shù)量隨放牧強度的增加也呈現(xiàn)先增多后減少的趨勢,中牧地的土壤微生物數(shù)量最多。
由以上學(xué)者的研究可見,草地土壤微生物數(shù)量與放牧強度間關(guān)系呈現(xiàn)3種變化趨勢。1)土壤微生物隨放牧強度的增加或者退化程度的加深而減少,也就是放牧強度越大,草原退化越嚴(yán)重則土壤微生物數(shù)量越少。2)隨放牧強度的增加先增多后減少的趨勢,即一定的放牧率引起土壤微生物增多,且適度的放牧率微生物數(shù)量達(dá)到最大值,而過度放牧不利于土壤微生物的生長所以減少。3)隨放牧強度的增加而減少,但在重度放牧?xí)r突然增多。多數(shù)研究結(jié)果呈現(xiàn)第二種變化趨勢。根據(jù)研究者們對試驗結(jié)果的分析發(fā)現(xiàn),土壤微生物數(shù)量與土壤某些理化性質(zhì)(比如:有機碳、全氮、pH值等)呈正相關(guān)關(guān)系。可見,一般情況下土壤微生物數(shù)量多時表示土壤質(zhì)量較好,放牧率合適,草地健康狀況良好(表1)。

表1 不同草地類型不同放牧強度下土壤微生物數(shù)量統(tǒng)計表
2放牧對土壤微生物生物量的影響
土壤微生物生物量是土壤營養(yǎng)元素的資源庫,是植物生長所需養(yǎng)分的重要來源[27-28]。有研究認(rèn)為與微生物個體數(shù)量相比,微生物生物量更能反映土壤中微生物的實際含量和潛在能力,所以土壤微生物生物量是反映土壤質(zhì)量和性能的一項重要的指標(biāo)[29-30]。在草地生態(tài)系統(tǒng)中,大部分的營養(yǎng)物質(zhì)被植物吸收利用,這些營養(yǎng)物質(zhì)又直接通過動物排泄物進(jìn)入能量循環(huán)過程,導(dǎo)致土壤中碳、氮加速分解轉(zhuǎn)化。已有少數(shù)學(xué)者研究發(fā)現(xiàn),放牧地比未放牧地的有機碳和氮的含量高,且微生物生物量也較高[31-33]。但是,有些學(xué)者認(rèn)為,草地放牧?xí)r存在潛在的氨揮發(fā)損失,所以有機氮含量較少[34-35]。可見土壤微生物生物量受放牧影響也較復(fù)雜。不同放牧方式下土壤微生物生物量變化見表2。
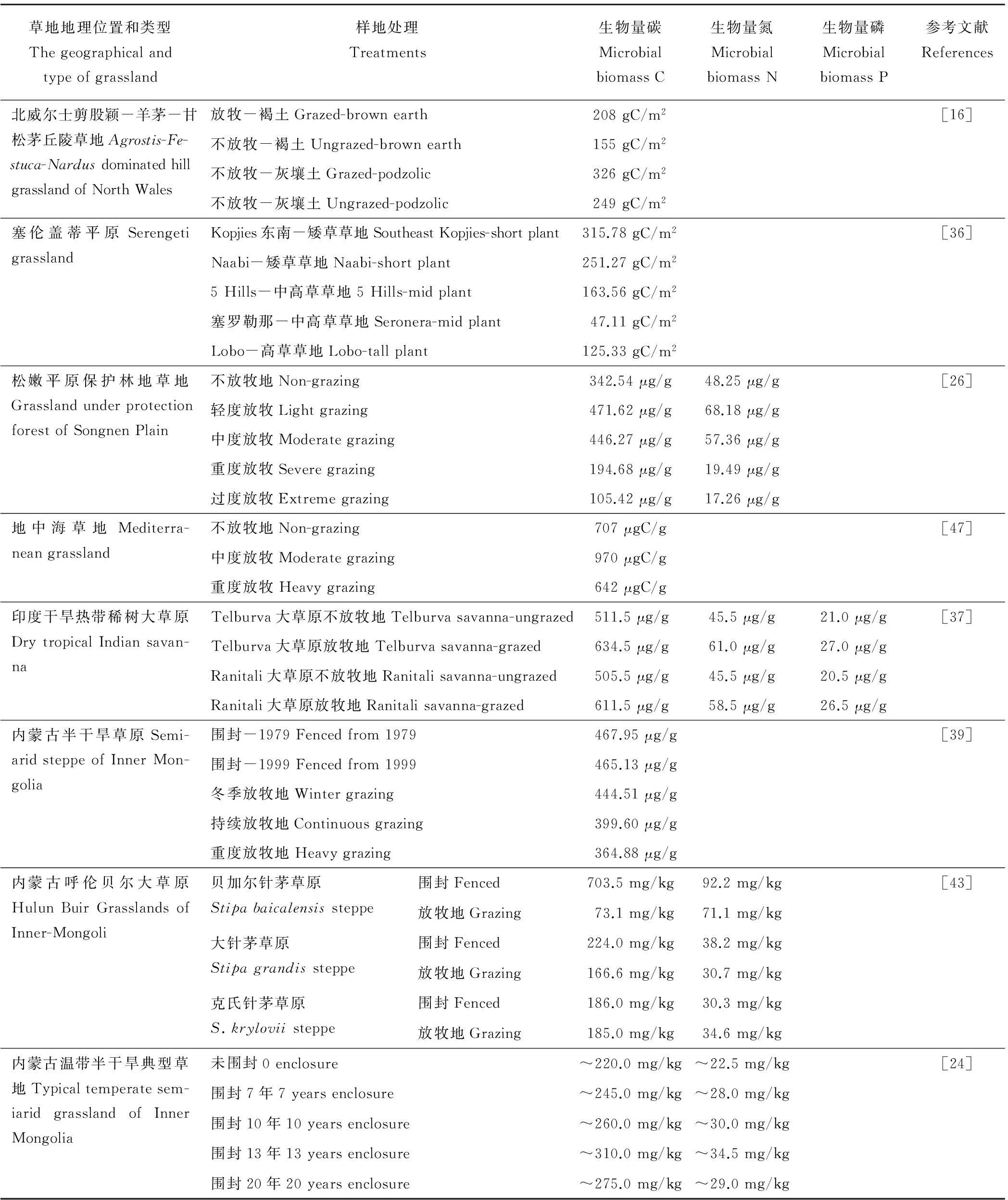
表2 不同草地類型不同放牧強度下土壤微生物生物量統(tǒng)計
Bardgett等[16]對兩種不同類型草原[剪股穎羊茅(Agrostisfestuca)草原和甘松茅(Nardusstricta)草原]放牧地研究發(fā)現(xiàn),兩類草原均表現(xiàn)出不放牧草地的表層土壤微生物生物量顯著低于放牧地,停止放牧?xí)r微生物生物量減少。Ruess和McNaughton[36]對塞倫蓋蒂平原矮草草地研究發(fā)現(xiàn),隨著放牧強度的增大,土壤微生物生物量碳含量增加,且與土壤有機碳呈正相關(guān)關(guān)系。Singh等[37]認(rèn)為放牧增加土壤有效營養(yǎng)物質(zhì)和微生物生物量,他們對印度干熱帶稀樹大草原研究發(fā)現(xiàn),放牧增加土壤微生物生物量碳(15%~18%)、氮(14%~23%)、磷(19%~29%),且與土壤有機氮、磷呈正相關(guān)關(guān)系。Bardgett等[38]對山地草甸放牧地研究發(fā)現(xiàn),放牧地的真菌生物量顯著高于未放牧地,而細(xì)菌生物量無顯著變化。研究者認(rèn)為,這種現(xiàn)象可能是由于酸性土壤的草地以及家畜排泄物有益于真菌的生長,并且放牧地出現(xiàn)嗜糞類微生物種類,導(dǎo)致放牧地真菌生物量高于未放牧地。
Qi等[39]對內(nèi)蒙古半干旱放牧地進(jìn)行研究發(fā)現(xiàn)重牧梯度下土壤微生物生物量碳最低,且與土壤有機碳和土壤全氮呈正相關(guān)關(guān)系。他們認(rèn)為土壤有機碳和全氮是決定土壤微生物生物量的主要影響因子,而且重牧地土壤有機碳和全氮減少是由于家畜過度采食導(dǎo)致植物生物量變少而引起的[40],同時裸地斑塊由于風(fēng)蝕作用導(dǎo)致土壤有機質(zhì)減少[41-42],也使土壤微生物生物量減少[12]。所以,重牧梯度下土壤有機碳含量低導(dǎo)致土壤微生物生物量降低。趙帥[43]對呼倫貝爾針茅(Stipacapillata)草原研究,結(jié)果顯示圍封樣地的微生物生物量碳、氮含量顯著高于放牧地。李玉潔等[44]研究表明,土壤微生物生物量隨休牧年限的增加而增多,休牧9年的微生物生物量碳、氮含量均最高。單貴蓮等[24]研究表明,圍封草地與自由放牧草地相比,微生物生物量碳、氮含量顯著增加,且隨圍封年限的延長呈增加趨勢。Northup等[13]關(guān)于熱帶禾草草原不同放牧梯度的研究顯示,隨放牧梯度的增大土壤微生物生物量減少。他們認(rèn)為這是由于放牧導(dǎo)致地表覆蓋度降低,植被凋落物生物量減少,從而減少了土壤有機物質(zhì)和碳的投入,限制了微生物生長的可用能源導(dǎo)致的[45]。
張?zhí)N薇等[46]關(guān)于人工草地的研究結(jié)果顯示,在放牧期間中牧區(qū)的微生物生物量碳、氮均最高,而在停牧期間輕牧區(qū)和不放牧區(qū)較高;隨放牧?xí)r間的延長,放牧強度越大微生物生物量碳、氮含量迅速降低。Wang和Sheng[26]研究表明土壤微生物生物量碳、氮隨放牧強度的增大呈先增多后減少的趨勢。以上兩位學(xué)者的研究都與Li 等[47]的研究結(jié)果一致。Li等[47]在地中海高山草甸試驗結(jié)果表明,中度放牧條件下的土壤呼吸速率和土壤微生物生物量最高,而重牧地的土壤微生物生物量最低。而且中牧地的土壤全氮、有機質(zhì)及鈉離子和鉀離子顯著高于重牧地。說明這些土壤理化性質(zhì)有益于土壤微生物群落的生長,從而加強了土壤營養(yǎng)物質(zhì)的循環(huán),使生產(chǎn)能力最大化。所以中度放牧梯度為最適放牧率。
總結(jié)以上關(guān)于土壤微生物生物量的研究,我們可以發(fā)現(xiàn)土壤微生物量與放牧強度間存在3種關(guān)系(表2):1)放牧增加土壤微生物生物量,且隨放牧強度的增加而增多。2)放牧減少土壤微生物生物量,且隨放牧強度的增加而減少。3)土壤微生物生物量隨放牧強度的增加,呈先增多后減少的趨勢。首先,雖然土壤微生物量與放牧間的關(guān)系復(fù)雜,但是從以上學(xué)者們的研究中發(fā)現(xiàn)土壤微生物生物量與土壤有機碳、有機氮、全氮、磷等其他土壤理化性質(zhì)呈正相關(guān)關(guān)系。不論在何種放牧率條件下,土壤有機碳等物質(zhì)含量高時,土壤微生物生物量就高。其次,學(xué)者們研究結(jié)果不一致,其中最主要的原因是因為放牧率不明確,由于試驗不同而放牧率不同,在大多數(shù)研究中并未明確放牧率,研究者們只是根據(jù)各自試驗區(qū)域的情況而劃分為輕度、中度、重度、過度放牧地。可見放牧率的范圍很模糊,這也為定量分析造成影響。因此,關(guān)于放牧對土壤微生物量的研究,一定要結(jié)合土壤理化性質(zhì)的分析,并且要以定量的放牧率為基礎(chǔ)。
3放牧對土壤微生物多樣性的影響
生物多樣性、干擾行為及生態(tài)系統(tǒng)功能三者間的相互關(guān)系一直是國際生態(tài)學(xué)界討論的熱點,Schwartz等[48]總結(jié)前人的研究結(jié)果發(fā)現(xiàn),95%的試驗結(jié)果證明生物多樣性與生態(tài)系統(tǒng)功能和過程間關(guān)系密切,但因為物種的特異性,它們之間的關(guān)系無規(guī)律。Hughes[49]指出干擾行為與生物多樣性間存在復(fù)雜的關(guān)系,并且總結(jié)出幾種常用的假說來解釋它們之間的相互作用。在草地生態(tài)系統(tǒng)中,土壤是一個包含有大量微生物生活的復(fù)雜生態(tài)系統(tǒng),土壤微生物多樣性與土壤代謝能力緊密相關(guān)[50],對環(huán)境的影響比在理化性質(zhì)方面更加重要[51]。有學(xué)者發(fā)現(xiàn)土壤微生物多樣性指標(biāo)與生態(tài)系統(tǒng)抵御外界干擾能力密切相關(guān)[52-53]。放牧是草地生態(tài)系統(tǒng)中長久以來最主要的一種干擾行為。因此,很多學(xué)者致力于放牧與土壤微生物多樣性間關(guān)系的研究,以期明確它們之間的關(guān)系,選擇最適放牧率,從而維持草地生態(tài)系統(tǒng)健康發(fā)展。隨著研究方法的不斷完善和出新,學(xué)者們運用不同的研究方法對各種類型草地放牧與土壤微生物多樣性間的關(guān)系進(jìn)行研究。
3.1 圍封與自由放牧對土壤微生物多樣性的影響
趙帥[43]運用PLFAs法和16S rDNA-DGGE法對呼倫貝爾針茅草原研究,結(jié)果顯示放牧條件下細(xì)菌磷脂脂肪酸含量顯著低于圍封草地,并發(fā)現(xiàn)存在7個菌門:變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、擬桿菌門(Bacteroidetes)、芽單胞菌門(Gemmatimonadetes)、酸桿菌門(Acidobacteria)、厚壁菌門(Firmicutes)和梭桿菌門(Fusobacteria)。李玉潔等[44]運用BIOLOG法對貝加爾針茅草原研究,發(fā)現(xiàn)自由放牧、休牧3年和休牧6年樣地間的土壤微生物群落碳源利用方式及代謝功能無差異,而休牧9年樣地的微生物群落具有不同的碳源利用方式和代謝功能。李梓正等[54]采用16S rDNA-DGGE對呼倫貝爾草原研究,結(jié)果表明:羊草(Leymuschinensis)草甸草原和貝加爾針茅草甸草原的微生物豐富度和多樣性在8月份最高,并且土壤微生物優(yōu)勢種群不受草地退化程度的影響。張海芳等[55]通過BIOLOG法對內(nèi)蒙古貝加爾針茅草原研究,發(fā)現(xiàn)自由放牧土壤微生物豐富度、均勻度和優(yōu)勢度均最高,圍封次之,刈割最低。周文萍等[56]采用高通量測序法對不同放牧強度下的AM真菌群落進(jìn)行研究,結(jié)果顯示AM真菌多樣性指數(shù)和均勻度指數(shù)在長期封育小區(qū)最低,且顯著低于中度放牧區(qū),而各放牧小區(qū)間差異不顯著。鄒雨坤等[57]運用PLFAs法對羊草草原進(jìn)行研究,發(fā)現(xiàn)刈割條件下的PLFA總量顯著高于圍封和放牧。Bardgett等[16]的研究結(jié)果為,長期圍封地表層土壤的真菌豐富度和真菌/細(xì)菌PLFA值顯著低于放牧地,但是細(xì)菌多樣性不受放牧地的影響。
3.2 不同放牧梯度對微生物多樣性的影響
Patra等[58]對不同放牧梯度下的功能微生物進(jìn)行研究,發(fā)現(xiàn)重牧地的細(xì)菌PLFAs高于輕牧地但真菌PLFAs沒有差異,而且重牧地與輕牧地間細(xì)菌群落結(jié)構(gòu)差異明顯,這與McCaig等[59]和Grayston等[60]的研究結(jié)果一致。Wakelin等[61]運用PCR-DGGE和TRFLP兩種方法研究發(fā)現(xiàn),放牧對土壤真菌和細(xì)菌群落結(jié)構(gòu)有影響,而且真菌豐富度隨放牧強度的增大而增多。Zhou等[62]對不同放牧梯度的細(xì)菌多樣性進(jìn)行研究,結(jié)果表明輕牧地和中牧地的細(xì)菌多樣性顯著高于未放牧地和重牧地。Howe等[63]研究認(rèn)為中度放牧強度下細(xì)菌多樣性顯著增多。劉天增和南志標(biāo)[64]運用BIOLOG法研究發(fā)現(xiàn)不放牧和中度放牧樣地的土壤微生物多樣性是最高的,而重度放牧下的土壤微生物多樣性最低[65]。Yang等[6]通過高通量測序方法研究表明,微生物遺傳多樣性隨放牧強度增大而減少。Nacke等[66]運用高通量測序法對森林和草地放牧地進(jìn)行研究發(fā)現(xiàn),未放牧的森林地土壤細(xì)菌多樣性高于放牧地,施肥未放牧地的細(xì)菌多樣性最高,然后是施肥放牧地,最低的是不施肥放牧地,可見放牧使草地細(xì)菌多樣性降低。Ford等[67]對沿海地區(qū)的鹽澤和沙丘兩種草地類型研究,結(jié)果顯示放牧對細(xì)菌PLFAs和真菌PLFAs影響不明顯。
3.3 放牧對土壤原生動物多樣性的影響
關(guān)于放牧對原生動物和線蟲的研究發(fā)現(xiàn),有些研究區(qū)域放牧?xí)咕€蟲的豐富度增加2倍[39],Sch?nborn[68]研究發(fā)現(xiàn)土壤重度緊實使原生動物種類顯著減少,土壤輕微緊實對土壤原生動物有抑制作用,這可能是由土壤孔隙度和土壤水分含量降低造成的。同時,Qi等[39]還發(fā)現(xiàn),中度放牧下變形蟲的生物多樣性最高,這是因為變形蟲中的優(yōu)勢種軀體柔軟可以進(jìn)入到其他原生動物進(jìn)入不了的微孔中,然后獲得食物和水進(jìn)行生活[69]。
由各位學(xué)者們研究可知,放牧對土壤微生物多樣性的影響規(guī)律不一致:第一,未放牧與放牧區(qū)間比較,一些學(xué)者研究顯示未放牧區(qū)的土壤微生物多樣性多于放牧區(qū),而有學(xué)者發(fā)現(xiàn)長期封育的小區(qū)多樣性指數(shù)低于放牧區(qū);第二,不同放牧強度樣地間比較,土壤微生物多樣性與放牧干擾間的關(guān)系表現(xiàn)為:1) 放牧強度對土壤微生物多樣性無顯著影響;2) 土壤微生物多樣性隨放牧強度或草地退化程度的加深而增多;3) 土壤微生物多樣性隨放牧強度增多而減少以及中度放牧強度下土壤微生物多樣性最多。這與Hughes[49]提出的關(guān)于干擾與生物多樣性間關(guān)系假說中的其中幾種假說一致。
4土壤微生物群落結(jié)構(gòu)和多樣性研究方法
土壤微生物多樣性,即群落結(jié)構(gòu)及功能、遺傳多樣性,在評估土壤質(zhì)量方面越來越受重視[70]。
4.1 磷脂脂肪酸譜圖法(PLFAs)
結(jié)構(gòu)多樣性的定義是系統(tǒng)內(nèi)部各部分或者組成元素所占比例情況,比如可用物種、基因組、群落數(shù)量來表示[71]。如今最常采用的是PLFAs法。由于土壤微生物擁有各自特定的磷脂脂肪酸(PLFAs),因此,可以直接從土壤中提取活的微生物細(xì)胞膜中的PLFAs,通過測定PLFAs的種類和含量得到微生物群落結(jié)構(gòu)組成的信息。自1990年以后,這種方法已經(jīng)廣泛應(yīng)用于各種環(huán)境問題引起微生物群落組成變化的研究[67,72-73],使土壤微生物多樣性得以更深入的研究。PLFAs法省時快速、耗費少、可重復(fù)而且能夠定量表征微生物的多樣性及群落結(jié)構(gòu)組成變化[74]。但是PLFAs法仍有不足:首先,關(guān)于脂肪酸表征特定微生物存在3個爭論點。第一,某些脂肪酸尚未確定類型,而且某些脂肪酸類型與微生物種類間對應(yīng)關(guān)系不明確。比如,PLFAs cy17:0 和cy19:0通常用于表征革蘭氏陰性細(xì)菌,但是Schoug 等[75]發(fā)現(xiàn)某些革蘭氏陽性菌里也大量存在這兩種PLFAs。Nichols等[76]在細(xì)菌中發(fā)現(xiàn)PLFAs 16:1ω5,而這種脂肪酸通常用于表征叢枝菌根真菌。第二,微生物生長環(huán)境會導(dǎo)致脂肪酸的類型及含量發(fā)生變化。比如,森林地土壤中,PLFAs 18:1ω9總是用于表征真菌群落,但是在農(nóng)耕地土壤中含量特別少,不能用來指代真菌[77]。第三,PLFAs法鑒定微生物種類水平不能到種,而且由于古細(xì)菌特殊的極性脂質(zhì)結(jié)構(gòu)無法用PLFAs方法進(jìn)行分析[78]。其次,PLFAs多樣性并不能真實反映土壤微生物多樣性。比如,細(xì)菌和真菌分別只有十幾種主要的PLFAs,而土壤中的細(xì)菌和真菌種類大約有上千種。所以,通過PLFAs計算得到的多樣性指數(shù)只能代表很小一部分的微生物多樣性。總之,PLFAs主要用于分析微生物種群結(jié)構(gòu)組成的變化。
4.2 遺傳分析技術(shù)
土壤微生物的遺傳多樣性分析是微生物多樣性的基礎(chǔ)和本質(zhì)。微生物在基因水平上的分類學(xué)多樣性主要依靠DNA編碼rRNA進(jìn)行分析。16S rRNA由于細(xì)菌和古細(xì)菌系統(tǒng)發(fā)育分析,18S rRNA用于真菌系統(tǒng)發(fā)育分析。遺傳多樣性的研究方法都基于PCR進(jìn)行分析的。下面主要介紹兩種方法:PCR-DGGE、高通量測序法。
4.2.1變形梯度凝膠電泳(PCR-DGGE)PCR-DGGE法廣泛應(yīng)用于微生物指紋識別,是基于16S rDNA片段的堿基組成和二級結(jié)構(gòu)變化進(jìn)行分析檢測的。運用特定的引物擴增16S rDNA靶序列,然后將擴增產(chǎn)物在含有梯度變性劑的聚丙烯酰胺凝膠中進(jìn)行電泳后形成譜帶,同樣長度但序列不同的DNA片段滯留于凝膠的不同位置,這些條帶的數(shù)量和位置來反映群落的優(yōu)勢種及其多樣性[79]。Bloem和Breure[80]運用PCR-DGGE法對60個農(nóng)場的土壤微生物多樣性進(jìn)行了監(jiān)測,發(fā)現(xiàn)不同的土壤類型及利用方式的微生物群落和多樣性不同。這一技術(shù)可以同時對多個樣品進(jìn)行比較分析,并且檢測到群落中的優(yōu)勢群落信息,然后通過對條帶切膠進(jìn)行序列分析,從而鑒定微生物群落種類。而且相較其他分子技術(shù),最突出的優(yōu)點為操作方便、耗時短且可重復(fù)實驗[81]。但是,同其他技術(shù)一樣,PCR-DGGE法也只能檢測到群落的部分多樣性。因為,每克土壤可能擁有1000種不同的微生物,但是這種方法只能獲得20~50個條帶,得到的系統(tǒng)進(jìn)化信息可能只有全部的1%[45],而且只能對500~1000 bp大小的基因片段進(jìn)行分離。此外,PCR會導(dǎo)致一些細(xì)菌種類基因多拷貝或者拷貝之間的異質(zhì)性問題,最終出現(xiàn)土壤微生物的多樣性被過多估計的情況。
4.2.2454焦磷酸測序法(Roche 454)Roche 454法是第二代高通量測序技術(shù)之一,已被廣泛應(yīng)用于宏基因組研究。可對環(huán)境中全部微生物的基因進(jìn)行測定,分析群體微生物的基因組成和多樣性及豐富度。Roche 454法的技術(shù)路線為:設(shè)計、合成含454接頭序列和Barcodede 的引物-PCR擴增-PCR產(chǎn)物定量、均一化-454測序儀測序-數(shù)據(jù)回歸樣品-生物信息學(xué)分析。筆者總結(jié)他人研究發(fā)現(xiàn),關(guān)于微生物多樣性研究的最大的難點是,無法獲得土壤中全部微生物的種類信息。但是,第二代高通量測序技術(shù)解決了這一問題。尤其是Roche 454法與傳統(tǒng)測序技術(shù)相比有以下幾個優(yōu)點:1) 實驗操作簡單、結(jié)果可重復(fù)性強、樣品DNA直接進(jìn)行測序、實驗周期短。2) 測序序列長、通量高,獲取信息量大,數(shù)據(jù)無偏向性,可以如實反映樣品微生物全部信息。3) 可以同時對土壤微生物中的優(yōu)勢種群、稀有種群及未培養(yǎng)菌群進(jìn)行檢測,到達(dá)微生物種的分類水平。在草地不同放牧管理方式對土壤微生物影響的研究中,近幾年越來越受重視。學(xué)者們已經(jīng)采用Roche 454法對青藏高原[6]、老撾熱帶草原[65]、德國森林地和草地[66]的不同管理方式對土壤微生物細(xì)菌和真菌影響進(jìn)行了深入的研究,獲得全面有意義的研究結(jié)果。筆者總結(jié)Roche 454法主要不足之處為:1) 測序費用過高,不利于一般小規(guī)模實驗研究。2) 所得數(shù)據(jù)結(jié)果信息量龐大,研究者必須從中仔細(xì)整理發(fā)現(xiàn)有價值、有意義的數(shù)據(jù),且要運用到較多分析軟件。總之,筆者認(rèn)為,Roche 454法是推進(jìn)環(huán)境微生物全面研究的革命性技術(shù),研究者們應(yīng)當(dāng)充分應(yīng)用發(fā)揮其優(yōu)點,為相關(guān)研究做出貢獻(xiàn)。
5研究展望
很多研究已經(jīng)證實,土壤功能微生物在養(yǎng)分循環(huán)的各個過程中起著重要的作用,因此全面了解土壤微生物多樣性及其相關(guān)功能對于草地可持續(xù)發(fā)展有著重要的意義。土壤微生物的數(shù)量、多樣性及群落結(jié)構(gòu)均是指示土壤質(zhì)量和草地生產(chǎn)力的重要指標(biāo)。適當(dāng)?shù)姆拍粮淖兺寥牢⑸锏纳瞽h(huán)境,有利于微生物的生長,增多微生物的多樣性,改變微生物的群落結(jié)構(gòu),進(jìn)而提高土壤質(zhì)量,使草地生產(chǎn)能力最大化。
關(guān)于放牧對土壤微生物的影響以及通過土壤微生物各種特性評估土壤質(zhì)量的研究越來越受重視。目前,雖然基本確定了應(yīng)該針對微生物的哪幾種指數(shù)進(jìn)行分析,但是很難明確哪種指數(shù)最適于作為指示指標(biāo),為了解決這個問題,需要對土壤微生物隨放牧行為的變化進(jìn)行更加深入的研究,進(jìn)而對于如何評價土壤質(zhì)量做出合理的解釋。此外,前文總結(jié)了放牧引起土壤微生物各種變化,但是還不清楚土壤微生物的數(shù)量和多樣性受到抑制時土壤質(zhì)量較好,還是土壤微生物產(chǎn)生功能冗余時表示土壤質(zhì)量較好。基于以上兩個問題,本文認(rèn)為未來的相關(guān)研究應(yīng)注意以下幾點:
1)要從土壤功能微生物著手研究。首先,土壤是各種物質(zhì)轉(zhuǎn)化進(jìn)程進(jìn)行的場所,比如:土壤碳轉(zhuǎn)化、氮轉(zhuǎn)化、磷鐵硫等元素的轉(zhuǎn)化以及土壤酶的作用,這些物質(zhì)的循環(huán)都是由土壤微生物驅(qū)動完成的。其次,土壤微生物數(shù)量龐大、體積微小、種類非常豐富。從宏觀和整體上,即大的分類群方面難以把握,造成分析結(jié)果往往沒有側(cè)重點,缺少針對性,從而很難揭示土壤微生物受干擾行為影響發(fā)生變化的本質(zhì)和規(guī)律。因此,加強功能微生物的研究,化整體為部分,將土壤某一物質(zhì)轉(zhuǎn)化進(jìn)程和相對的功能微生物結(jié)合起來,對它們的相關(guān)關(guān)系加以研究,更有利于深入研究土壤微生物是如何響應(yīng)干擾行為,能夠提供更為有效、直接的基礎(chǔ)數(shù)據(jù)。
2)要加強植物多樣性和土壤微生物多樣性相互關(guān)系的研究。首先,供試區(qū)域的植物種類及其多樣性是構(gòu)成整個環(huán)境生態(tài)體系的最重要的部分。放牧行為最先直接影響地被植物,進(jìn)而對土壤微生物形成間接作用。其次,供試區(qū)域不同,植被種類不同,從而土壤微生物類群不同。放牧行為與植物多樣性及土壤微生物多樣性,三者間相互聯(lián)系,互相影響。因此,注重這方面的研究,對于明確草地土壤微生物對放牧行為的響應(yīng)機制有重要的意義。
3)要將土壤理化性質(zhì)與土壤微生物相結(jié)合進(jìn)行分析。土壤微生物不僅可以作為一個綜合指標(biāo),單獨用于評估土壤質(zhì)量。而且作為土壤生化性質(zhì),又是土壤各種性質(zhì)中的一個指數(shù)。因此,必須將它與土壤的理化性質(zhì)相結(jié)合,才能全面合理的表征土壤質(zhì)量。同時,根據(jù)它與土壤理化性質(zhì)間的相互關(guān)系,可以明確土壤微生物的什么指數(shù)具體是怎么響應(yīng)放牧行為的,與放牧強度間是呈正相關(guān)關(guān)系還是呈負(fù)相關(guān)關(guān)系抑或不相關(guān)。
4)最終建立生態(tài)模型,對草地生態(tài)系統(tǒng)做出合理預(yù)測和評估。在關(guān)于草地生態(tài)系統(tǒng)的研究中,對我國草地資源生態(tài)可持續(xù)發(fā)展性和可利用性進(jìn)行評價和預(yù)測是很重要的一部分研究。我們對各種導(dǎo)致草地退化的干擾行為和草地恢復(fù)機制的研究,最終目的都是為了掌握草地退化過程中可預(yù)見性的問題,從而做出相應(yīng)的對策,以期維護(hù)草地長久良好的發(fā)展下去。因此,土壤微生物作為土壤重要的生化性質(zhì),將它與其他環(huán)境指標(biāo)結(jié)合,建立更加合理科學(xué)的生態(tài)模型是非常重要的一部分工作。
References:
[1]McNaughton S J. Grazing as an optimization process: grass-ungulate relationships in the Serengeti. American Naturalist, 1979, 113(5): 691-703.
[2]Dyer M I, Turner C L, Seastedt T R. Herbivory and its consequences. Ecological Applications, 1993, 3(3): 10-16.
[3]Frank D A, Groffman P M. Ungulate vs. landscape control of soil C and N processes in grasslands of Yellowstone National Park. Ecology, 1998, 79(7): 2229-2241.
[4]Bezemer T M, van Dam N M. Linking aboveground and belowground interactions via induced plant defenses. Trends in Ecology & Evolution, 2005, 20(11): 617-624.
[5]Hamilton III E W, Frank D A. Can plants stimulate soil microbes and their own nutrient supply? Evidence from a grazing tolerant grass. Ecology, 2001, 82(9): 2397-2402.
[6]Yang Y F, Wu L W, Lin Q Y,etal. Responses of the functional structure of soil microbial community to livestock grazing in the Tibetan alpine grassland. Global Change Biology, 2013, 19(2): 637-648.
[7]Qu T B, Zhang J F, Du W C,etal. Research progress on effects of grazing on diversity of microorganisms in grassland soil. Contemporary Eco-Agriculture, 2012, (3): 14-20.
[8]Bardgett R D, Wardle D A, Yeates G W. Linking above-ground and below-ground interactions: how plant responses to foliar herbivory influence soil organisms. Soil Biology and Biochemistry, 1998, 30(14): 1867-1878.
[9]Wardle D A, Nicholson K S. Synergistic effects of grassland plant species on soil microbial biomass and activity: implications for ecosytem-level effects of enriched plant diversity. Functional Ecology, 1996, 10: 410-416.
[10]Kohler F, Hamelin J, Gillet F,etal. Soil microbial community changes in wooded mountain pastures due to simulated effects of cattle grazing. Plant and Soil, 2005, 278(1-2): 327-340.
[11]Maharning A R, Mills A A, Adl S M. Soil community changes during secondary succession to naturalized grasslands. Applied Soil Ecology, 2009, 41(2): 137-147.
[12]Holt J. Grazing pressure and soil carbon, microbial biomass and enzyme activities in semi-arid northeastern Australia. Applied Soil Ecology, 1997, 5(2): 143-149.
[13]Northup B K, Brown J R, Holt J A. Grazing impacts on the spatial distribution of soil microbial biomass around tussock grasses in a tropical grassland. Applied Soil Ecology, 1999, 13(3): 259-270.
[14]Wang K H, McSorley R, Bohlen P,etal. Cattle grazing increases microbial biomass and alters soil nematode communities in subtropical pastures. Soil Biology and Biochemistry, 2006, 38(7): 1956-1965.
[15]Regan K M, Nunan N, Boeddinghaus R S,etal. Seasonal controls on grassland microbial biogeography: Are they governed by plants, abiotic properties or both? Soil Biology and Biochemistry, 2014, 71: 21-30.
[16]Bardgett R D, Leemans D K, Cook R,etal. Seasonality of the soil biota of grazed and ungrazed hill grasslands. Soil Biology and Biochemistry, 1997, 29(8): 1285-1294.
[17]Gans J, Wolinsky M, Dunbar J. Computational improvements reveal great bacterial diversity and high metal toxicity in soil. Science, 2005, 309(5739): 1387-1390.
[18]Daniel R. The metagenomics of soil. Nature Reviews Microbiology, 2005, 3(6): 470-478.
[19]Zhao J. Effect of stocking reats on soil microbial number and biomass in steppe. Acta Agrestia Sinica, 1999, 7(3): 223-227.
[20]Zhang C X, Nan Z B. Changeable characteristics of three soil microbial groups under different grazing intensities in Loess Plateau. Pratacultural Science, 2011, 27(11): 131-136.
[21]Wang C T, Long R J, Wang G X,etal. Relationship between plant communities, characters, soil physical and chemical properties, and soil microbiology in alpine meadows. Acta Prataculturae Sinica, 2010, 19(6): 25-34.
[22]Liu S G, Ge S R, Long Z F. Studies on soil microorganism numbers andmicrobiota of degenerated rangelands in northwest region of Sichuan P R.China. Acta Prataculturae Sinica, 1994, 3(4): 70-76.
[23]Li C L, Zhao M L, Han G D,etal. The effect of grazing on soil micro-organism and soil nutrients and their seasonal dynamics inStipabrevifloraSteppe. Journal of Arid Land Resources and Environment, 2009, 23(4): 184-190.
[24]Shan G L, Chen G, Ning F,etal. Dynamics of soil microorganism and enzyme activity in typical steppe of restoration succession process. Acta Agrestia Sinica, 2012, 20(2): 292-297.
[25]Gu A X, Fan Y M, Wu H Q,etal. Relationship between the number of three main microorganisms and the soil environment of degraded grassland on the north slope of the Tianshan Mountains. Acta Prataculturae Sinica, 2010, 19(2): 116-123.
[26]Wang X Z, Sheng L X. Effect of grazing intensity on microorganisms quantity and microbial biomass of soil in grassland under protection forest of songnen plain. Journal of Animal and Veterinary Advances, 2012, 11(24): 4549-4552.
[27]Smith J L, Paul E A. The significance of soil microbial biomass estimations. Soil Biochemistry, 1990, 6: 357-396.
[28]Kennedy A C, Smith K L. Soil microbial diversity and the sustainability of agricultural soils. Plant and Soil, 1995, 170(1): 75-86.
[29]Chen G C. Soil microbial quantity determination method and its application in red soil. Chinese Journal of Soil Science, 1999, 30(6): 197-200.
[30]Gao Y C, Zhou W S, Chen W X,etal. Estimation for biomass and turnover of soil microorganisms. Chinese Journal of Ecology, 1993, 12(6): 6-10.
[31]Risser P G, Birney E C, Blocker H D,etal. The True Prairie Ecosystem[M]. Pennsylvania: Hutchinson Ross Publishing Company Stroudsburg, 1981.
[32]Ross D J, Tate K R. Microbial biomass in soil: effects of some experimental variables on biochemical estimations. Soil Biology and Biochemistry, 1984, 16(2): 161-167.
[33]Sarathchandra S U, Perrott K W, Upsdell M P. Microbiological and biochemical characteristics of a range of New Zealand soils under established pasture. Soil Biology and Biochemistry, 1984, 16(2): 177-183.
[34]Martin J P, Chapman H D. Volatilization of ammonia from surface-fertilized soils. Soil Science, 1951, 71(1): 25-34.
[35]Stillwell M A, Woodmansee R G. Chemical transformations of urea-nitrogen and movement of nitrogen in a shortgrass prairie soil. Soil Science Society of America Journal, 1981, 45(5): 893-898.
[36]Ruess R W, McNaughton S J. Grazing and the dynamics of nutrient and energy regulated microbial processes in the Serengeti grasslands. Oikos, 1987, 49(1): 101-110.
[37]Singh R S, Srivastava S C, Raghubanshi A S,etal. Microbial C, N and P in dry tropical savanna: effects of burning and grazing. Journal of Applied Ecology, 1991, 28(3): 869-878.
[38]Bardgett R D, Hobbs P J, Frosteg?rd ?. Changes in soil fungal: bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biology and Fertility of Soils, 1996, 22(3): 261-264.
[39]Qi S, Zheng H, Lin Q,etal. Effects of livestock grazing intensity on soil biota in a semiarid steppe of Inner Mongolia. Plant and Soil, 2011, 340: 117-126.
[40]Gao Y Z, Giese M, Lin S,etal. Belowground net primary productivity and biomass allocation of a grassland in Inner Mongolia is affected by grazing intensity. Plant and Soil, 2008, 307(1-2): 41-50.
[41]Hoffmann C, Funk R, Wieland R,etal. Effects of grazing and topography on dust flux and deposition in the Xilingele grassland, Inner Mongolia. Journal of Arid Environments, 2008, 72(5): 792-807.
[42]Schneider K, Huisman J, Breuer L,etal. Ambiguous effects of grazing intensity on surface soil moisture: A geostatistical case study from a steppe environment in Inner Mongolia, PR China. Journal of Arid Environments, 2008, 72(7): 1305-1319.
[43]Zhao S. Effects of Grazing and Fenced on Soil Microbial Diversity in Stipa Steppes of Hulunbeier, Inner Mongolia[D]. Beijing: Chinese Academy of Agricultural Sciences, 2011.
[44]Li Y J, Li G, Song X L,etal. The influence of rest-grazing on the soil microbial community functional diversity ofStipabaicalensisSteppe. Acta Prataculturae Sinica, 2013, 22(6): 21-30.
[45]Casamayor E O, Sch?fer H, Baeras L,etal. Identification of and spatio-temporal differences between microbial assemblages from two neighboring sulfurous lakes: comparison by microscopy and denaturing gradient gel electrophoresis. Applied and Environmental Microbiology, 2000, 66(2): 499-508.
[46]Zhang Y W, Han J G, Han Y W,etal. The content of soil micro-biomass carbon and nitrogen of different grazing Intensities on Pasture. Acta Agrestia Sinica, 2003, 11(4): 343-346.
[47]Li Q, Mayzlish E, Shamir I,etal. Impact of grazing on soil biota in a Mediterranean grassland. Land Degradation & Development, 2005, 16(6): 581-592.
[48]Schwartz M W, Brigham C A, Hoeksema J D,etal. Linking biodiversity to ecosystem function: implications for conservation ecology. Oecologia, 2000, 122(3): 297-305.
[49]Hughes A. Disturbance and diversity: an ecological chicken and egg problem. Nature Education Knowledge, 2012, 3(10): 48.
[50]Nannipieri P, Ascher J, Ceccherini M,etal. Microbial diversity and soil functions. European Journal of Soil Science, 2003, 54(4): 655-670.
[51]Insam H. Developments in soil microbiology since the mid 1960s. Geoderma, 2001, 100(3): 389-402.
[52]Johnsen K, Jacobsen C S, Torsvik V,etal. Pesticide effects on bacterial diversity in agricultural soils-a review. Biology and Fertility of Soils, 2001, 33(6): 443-453.
[53]Sugden A M. Diversity & ecosystem resilience. Science, 2000, 290(5490): 233-235.
[54]Li Z Z, Zhu L B, Lin Y C,etal. Seasonal variation of soil bacterial community under different degrees of degradation of Hulunbuir grassland. Acta Ecologica Sinica, 2010, 30(11): 2883-2889.
[55]Zhang H F, Li G, Song X L,etal. Functional diversity of soil microbial communities inStipabaicalensissteppe in Inner Mongolia as affected by different land use patterns. Chinese Journal of Ecology, 2012, 31(5): 1143-1149.
[56]Zhou W P, Xiang D, Hu W J,etal. Influences of long-term enclosure on the restoration of plant and AM fungal communities on grassland under different grazing intensities. Acta Ecologica Sinica, 2013, 33(11):3383-3393.
[57]Zhou Y K, Zhang J N, Yang D L,etal. Phospholipid fatty acid analysis of microbial community structure under different land use patterns in soil ecosystems ofLeymuschinensissteppes. Acta Prataculturae Sinica, 2011, 20(4): 27-33.
[58]Patra A K, Abbadie L, Clays-Josserand A,etal. Effects of grazing on microbial functional groups involved in soil N dynamics. Ecological Monographs, 2005, 75(1): 65-80.
[59]McCaig A E, Glover L A, Prosser J I. Molecular analysis of bacterial community structure and diversity in unimproved and improved upland grass pastures. Applied and Environmental Microbiology, 1999, 65(4): 1721-1730.
[60]Grayston S J, Griffith G S, Mawdsley J L,etal. Accounting for variability in soil microbial communities of temperate upland grassland ecosystems. Soil Biology and Biochemistry, 2001, 33(4): 533-551.
[61]Wakelin S A, Gregg A L, Simpson R J,etal. Pasture management clearly affects soil microbial community structure and N-cycling bacteria. Pedobiologia, 2009, 52(4): 237-251.
[62]Zhou X, Wang J, Hao Y,etal. Intermediate grazing intensities by sheep increase soil bacterial diversities in an Inner Mongolian steppe. Biology and Fertility of Soils, 2010, 46(8): 817-824.
[63]Howe H F, Brown J S, Zorn-Arnold B. A rodent plague on prairie diversity. Ecology Letters, 2002, 5(1): 30-36.
[64]Liu T Z, Nan Z B. Advances in nitrification and nitrifying bacteria in grassland soil. Pratacultural Science, 2011, 28(6): 951-958.
[65]Lienhard P, Terrat S, Chemidlin N. Pyrosequencing evidences the impact of cropping on soil bacterial and fungal diversity in Laos tropical graddland. Agronomy for Sustainable Development, 2013, 34: 525-533.
[66]Nacke H, Thürmer A, Wollherr A,etal. Pyrosequencing-based assessment of bacterial community structure along different management types in German forest and grassland soils. PloS One, 2011, 6(2): e17000.
[67]Ford H, Rousk J, Garbutt A,etal. Grazing effects on microbial community composition, growth and nutrient cycling in salt marsh and sand dune grasslands. Biology and Fertility of Soils, 2013, 49(1): 89-98.
[68]Sch?nborn W. Population dynamics and production biology of testate amoebae (Rhizopoda, Testacea) in raw humus of two coniferous forest soils. Archiv für Protistenkunde, 1986, 132(4): 325-342.
[69]Gao Y, Zhu W, Chen W. Structure of the protozoan community in soil and its ecological functions. Chinese Journal of Ecology, 2000, 19(1): 59.
[70]Visser S, Parkinson D. Soil biological criteria as indicators of soil quality: soil microorganisms. American Journal of Alternative Agriculture, 1992, 7(1-2): 33-37.
[71]?vre?s L. Population and community level approaches for analysing microbial diversity in natural environments. Ecology Letters, 2000, 3(3): 236-251.
[72]Clegg C D. Impact of cattle grazing and inorganic fertiliser additions to managed grasslands on the microbial community composition of soils. Applied Soil Ecology, 2006, 31(1): 73-82.
[73]Bardgett R D, Jones A C, Jones D L,etal. Soil microbial community patterns related to the history and intensity of grazing in sub-montane ecosystems. Soil Biology and Biochemistry, 2001, 33(12): 1653-1664.
[74]Frosteg?rd ?, Tunlid A, B??th E. use and misuse of PLFA measurements in soils. Soil Biology and Biochemistry, 2011, 43: 1621-1625.
[75]Schoug ?, Fischer J, Heipieper H J,etal. Impact of fermentation pH and temperature on freeze-drying survival and membrane lipid composition of Lactobacillus coryniformis Si3. Journal of Industrial Microbiology & Biotechnology, 2008, 35(3): 175-181.
[76]Nichols P, Stulp B K, Jones J G,etal. Comparison of fatty acid content and DNA homology of the filamentous gliding bacteria Vitreoscilla, Flexibacter, Filibacter. Archives of Microbiology, 1986, 146(1): 1-6.
[77]Olsson P A. Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal fungi in soil. FEMS Microbiology Ecology, 1999, 29(4): 303-310.
[78]Lin X G. Soil Microbial Research Principles and Methods[M]. Beijing: High Education Press, 2010.
[79]Muyzer G, De Waal E C, Uitterlinden A G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Applied and Environmental Microbiology, 1993, 59(3): 695-700.
[80]Bloem J, Breure A M. Microbial indicators. Trace Metals and other Contaminants in the Environment, 2003, 6: 259-282.
[81]Ma Y X, Holmstrm C, Webb J. Application of denaturing gradient gel electrophoresis (DGGE) in microbial ecology. Acta Ecologica Sinica, 2003, 23(8): 1561-1569.
參考文獻(xiàn):
[7]曲同寶, 張建峰, 杜瑋超, 等. 放牧對草地土壤微生物多樣性影響研究進(jìn)展. 當(dāng)代生態(tài)農(nóng)業(yè), 2012, (3): 14-20.
[19]趙吉. 不同放牧率對冷篙小禾草草原土壤微生物數(shù)量和生物量的影響. 草地學(xué)報, 1999, 7(3): 223-227.
[20]張成霞, 南志標(biāo). 不同放牧強度下隴東天然草地土壤微生物三大類群的動態(tài)特征. 草業(yè)科學(xué), 2011, 27(11): 131-136.
[21]王長庭, 龍瑞軍, 王根緒, 等. 高寒草甸群落地表植被特征與土壤理化性狀, 土壤微生物之間的相關(guān)性研究. 草業(yè)學(xué)報, 2010, 19(6): 25-34.
[22]劉世貴, 葛紹榮, 龍章富. 川西北退化草地土壤微生物數(shù)量與區(qū)系研究. 草業(yè)學(xué)報, 1994, 3(4):70-76.
[23]李春莉, 趙萌莉, 韓國棟, 等. 放牧對短花針茅草原土壤微生物和土壤養(yǎng)分的影響及其季節(jié)動態(tài). 干旱區(qū)資源與環(huán)境, 2009, 23(4): 184-190.
[24]單貴蓮, 陳功, 寧發(fā), 等. 典型草原恢復(fù)演替過程中土壤微生物及酶活性動態(tài)變化研究. 草地學(xué)報, 2012, 20(2): 292-297.
[25]顧愛星, 范燕敏, 武紅旗, 等. 天山北坡退化草地土壤環(huán)境與微生物數(shù)量的關(guān)系. 草業(yè)學(xué)報, 2010, 19(2): 116-123.
[29]陳國潮. 土壤微生物量測定方法現(xiàn)狀及其在紅壤上的應(yīng)用. 土壤通報, 1999, 30(6): 197-200.
[30]高云超, 朱文珊, 陳文新. 土壤微生物生物量周轉(zhuǎn)的估算. 生態(tài)學(xué)雜志, 1993, 12(6): 6-10.
[43]趙帥. 放牧與圍封對呼倫貝爾針茅草原土壤微生物多樣性的影響[D]. 北京: 中國農(nóng)業(yè)科學(xué)院, 2011.
[44]李玉潔, 李剛, 宋曉龍, 等. 休牧對貝加爾針茅草原土壤微生物群落功能多樣性的影響. 草業(yè)學(xué)報, 2013, 22(6): 21-30.
[46]張?zhí)N薇, 韓建國, 韓永偉, 等. 不同放牧強度下人工草地土壤微生物量碳, 氮的含量. 草地學(xué)報, 2003, 11(4): 343-346.
[54]李梓正, 朱立博, 林葉春, 等. 呼倫貝爾草原不同退化梯度土壤細(xì)菌多樣性季節(jié)變化. 生態(tài)學(xué)報, 2010, 30(11): 2883-2889.
[55]張海芳, 李剛, 宋曉龍, 等. 內(nèi)蒙古貝加爾針茅草原不同利用方式土壤微生物功能多樣性.生態(tài)學(xué)雜志, 2012, 31(5): 1143-1149.
[56]周文萍, 向丹, 胡亞軍, 等. 長期圍封對不同放牧強度下草地植物和 AM 真菌群落恢復(fù)的影響. 生態(tài)學(xué)報, 2013, 33(11): 3383-3393.
[57]鄒雨坤, 張靜妮, 楊殿林, 等. 不同利用方式下羊草草原土壤生態(tài)系統(tǒng)微生物群落結(jié)構(gòu)的PLFA分析. 草業(yè)學(xué)報, 2011, 20(4): 27-33.
[64]劉天增, 南志標(biāo). 草地硝化微生物與硝化作用研究進(jìn)展. 草業(yè)科學(xué), 2011, 28(6): 951-958.
[78]林先貴. 土壤微生物研究原理與方法[M]. 北京: 高等教育出版社, 2010.
[81]馬悅欣, Holmstrm C, Webb J. 變性梯度凝膠電泳(DGGE)在微生物生態(tài)學(xué)中的應(yīng)用. 生態(tài)學(xué)報, 2003, 23(8): 1561-1569.
http://cyxb.lzu.edu.cn
茍燕妮, 南志標(biāo). 放牧對草地土壤微生物的影響. 草業(yè)學(xué)報, 2015, 24(10): 194-205.
GOU Yan-Ni, NAN Zhi-Biao. The impacts of grazing on the soil microorganism population of grassland. Acta Prataculturae Sinica, 2015, 24(10): 194-205.
通訊作者*Corresponding author. E-mail: zhibiao@lzu.edu.cn
作者簡介:茍燕妮(1987-), 女, 甘肅臨洮人,在讀博士。E-mail: gouyannigg@126.com
基金項目:國家重點基礎(chǔ)研究發(fā)展計劃(“973”計劃)項目(編號:2014CB138702)資助。
收稿日期:2014-11-06;改回日期:2015-03-18
DOI:10.11686/cyxb2014460
