PRRSV體外感染PAMs對IFN-γ mRNA轉錄水平的影響
2016-02-07 07:50:45閆曉霞劉歡歡曾夢穎高洪嚴玉霖
中國獸藥雜志 2016年10期
閆曉霞,劉歡歡,曾夢穎,高洪,嚴玉霖
(云南農業大學動物科學技術學院,昆明 650201)
PRRSV體外感染PAMs對IFN-γ mRNA轉錄水平的影響
閆曉霞,劉歡歡,曾夢穎,高洪,嚴玉霖*
(云南農業大學動物科學技術學院,昆明 650201)
為探討豬繁殖與呼吸綜合征病毒(PRRSV)體外感染豬肺泡巨噬細胞(PAMs)對IFN-γ mRNA轉錄水平的影響,選用PRRSV血清抗體和抗原均為陰性的4周齡健康仔豬3頭,無菌分離PAMs,隨機將其分為對照組、PRRSV感染組(PRRSV組)、PRRSV+PHA組和RRRSV+Dex組,分別在培養6、12、24、48和60 h收集PAMs。運用熒光定量PCR檢測IFN-γ和PRRSV mRNA轉錄情況。結果顯示,PRRSV mRNA轉錄量在感染PAMs后6~24 h逐漸升高,在48 h有所下調,在60 h再次升高,IFN-γ mRNA轉錄量在6~48 h逐漸升高,24、48 h顯著高于對照組,而到60 h又恢復到對照組水平;與同時間點的PRRSV組相比,PRRSV+PHA組中PRRSV mRNA轉錄量有所下調,IFN-γ mRNA轉錄量有所上調;而RRRSV+Dex組PRRSV mRNA轉錄量有所上調,IFN-γ mRNA轉錄量所下調。結果表明,IFN-γ 在一定程度上可以抑制PRRSV在PAMs中的增殖,IFN-γ的抑制可能是PRRSV導致細胞損傷的機制之一。
豬繁殖與呼吸綜合征病毒;肺泡巨噬細胞;干擾素-γ
豬繁殖與呼吸綜合征病毒(Porcine reproductive and respiratory syndrome virus,PRRSV)屬于動脈炎病毒科動脈炎病毒屬,是一種有囊膜的不分節段的單股正鏈RNA病毒[1-2]。根據基因組及致病性的差異,PRRSV可分為2個型,即歐洲型(LV株為代表株)和美洲型(ATCC-VR2332株為代表株)[3]。PRRSV主要侵害肺泡巨噬細胞(Pulmonary alveolar macrophage,PAMs),從而導致彌散性間質性肺炎,損害機體免疫機能,進一步對機體造成繼發性感染[4],給世界的養豬業帶來了巨大的經濟損失[5]。干擾素(interferon,IFN)是一類具有廣譜抗病毒、抗腫瘤和增強免疫功能的細胞因子,主要由活化的T 細胞、NK 細胞和巨噬細胞產生,可誘導細胞產生多種抗病毒蛋白,促進抗病毒免疫[6-7]。 國內外對PRRSV感染PAMs的研究主要局限在I型干擾素。鑒于此,本研究運用實時熒光定量PCR技術,對PRRSV 感染PAMs不同時期IFN-γmRNA轉錄水平的變化進行檢測,從而了解IFN-γ與PRRSV 抗感染之間的關系,為PRRSV的防控提供參考依據和技術支持。
1 材料與方法
1.1 材料
1.1.1 病毒 PRRSV云南株YN-2011(登錄號:JX857698)由本實驗室分離純化保存,其TCID50為1 mL 105.06。
1.1.2 實驗動物 經ELISA 和RT-PCR方法檢測PRRSV血清抗體和抗原均為陰性的4周齡健康仔豬3頭。
1.1.3 主要儀器與試劑 XDS-2倒置顯微鏡購自重慶光電儀器總公司;CO2培養箱購自美國Thermo scientific公司;BIO-RAD-CFX熒光定量PCR儀購自美國BIO-RAD公司。RPMI-1640培養基、胎牛血清購自美國Hyclone公司;PRRSV抗體ELISA檢測試劑盒購自北京愛德士元亨生物科技有限公司;PHA-L購自美國Sigma公司;Trizol試劑購自美國Invitrogen公司;PrimeScriptTMRT reagent Kit with gDNA eraser、SYBR Premix Ex TaqTM (Tli RNaseH Plus)購自寶生物工程( 大連) 有限公司。
1.2 方法
1.2.1 肺泡巨噬細胞的制備 將試驗豬置于無菌室解剖臺上,頸靜脈放血致死,剖開胸腔,結扎氣管后連同心臟取出完整的肺,用0.01 g/L pH 7.4的PBS充分漂洗肺表面,清除血塊,從氣管往肺注入0.01 g/L pH 7.4的PBS 50~100 mL,輕輕拍打肺表面1~2 min后回收灌洗液,用單層無菌過濾篩過濾,收集全部灌洗液,1500 r/min離心5 min,最后用含有雙抗的 PBS洗滌PAMs兩次,1500 r/min離心5 min,即為所得PAM。RPMI-1640培養基重懸細胞并計數,調整細胞濃度為2×106mL-1,以每孔2 mL鋪在6孔細胞板于37 ℃、 5% CO2培養箱培養2 h,當PAMs貼壁后,棄上清,待用。
1.2.2 試驗分組與處理 將生長在6 孔板上的PAMs 細胞分為4 組。正常對照組;PRRSV感染組;PRRSV+PHA組:在經過PRRSV感染后的PAMs中加入5 μg/mL的PHA-L進行處理[8];RRRSV+Dex組:在經過PRRSV感染后的PAMs中加入0.1 mg/mL的Dex進行處理[9]。每組設3個重復。各組加入的病毒量為100 μL,對照組接種等體積的細胞培養液,分別于感染后6、12、24、48和60 h收集PAMs,-80 ℃保存,用于熒光定量PCR檢測。
1.2.3 熒光定量PCR檢測PRRSV和IFN-γ mRNA轉錄水平 使用Trizol提取總RNA,應用PrimeScriptTMRT reagent Kit with gDNA eraser試劑盒逆轉錄總RNA成為cDNA,于-20 ℃保存。Realtime-PCR反應體系: SYBR Premix Ex TaqII(Tli RNaseH Plus)12.5 μL, 上、下游引物(表1)各1.0 μL,cDNA 2 μL,dH20 8.5 μL。按以下程序進行Realtime-PCR反應:95 ℃預變性30 s;95 ℃ 5 s,Tm 30 s;72 ℃ 30 s,共40個循環。每個樣本設置3個重復孔。

表1 實時熒光定量PCR引物
1.2.4 IFN-γ與PRRSV mRNA轉錄水平相關性分析 根據各試驗組各時間點IFN-γ與PRRSV mRNA相對轉錄量,以PRRSV mRNA轉錄水平為縱坐標,IFN-γ mRNA轉錄水平為橫坐標,做轉錄水平相關性分析。
2 結果與分析
2.1 細胞中IFN-γ mRNA轉錄 根據不同時間點Ct值,用2-△△Ct計算不同組IFN-γ mRNA相對轉錄量。IFN-γ mRNA在各組不同時間點的轉錄水平如圖1。PRRSV組中,IFN-γ mRNA轉錄量在6~48 h逐漸升高,6 h顯著低于對照組(P<0.01),24和48 h顯著高于對照組(P<0.01),而到60 h又恢復到對照組水平;PRRSV+PHA組IFN-γ mRNA轉錄水平顯著高于對照組及PRRSV組;RRRSV+Dex組IFN-γ mRNA轉錄水平在6、12、24和60 h顯著低于對照組和PRRSV組(P<0.01或P<0.05)。
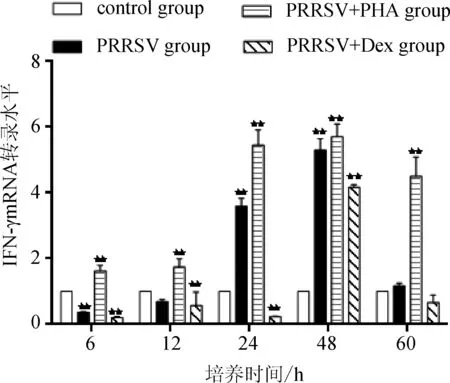
圖1 IFN-γ mRNA在各組不同時間點的轉錄水平(*P<0.05,**P<0.01)

圖2 PRRSV mRNA在各組不同時間點的轉錄水平(*P<0.05,**P<0.01)
2.2 細胞中PRRSV mRNA轉錄 根據不同時間點Ct值,用2-△△Ct計算不同組PRRSV mRNA相對轉錄量。由圖2可以看出,PRRSV組PRRSV mRNA轉錄量在感染后6~24 h逐漸升高,48 h有所下調,60 h再次升高;PRRSV+PHA組PRRSV mRNA的轉錄量與相同時間點PRRSV組中PRRSV mRNA的轉錄量相比顯著下調(P<0.01或P<0.05);RRRSV+Dex組PRRSV mRNA的轉錄量與相同時間點PRRSV感染組相比有所上調,在12、24 h差異顯著(P<0.01或P<0.05)。
2.3 IFN-γ與PRRSV mRNA轉錄水平相關性分析 不同試驗組之間IFN-γ與PRRSV mRNA相關性分析見圖3。根據|R|∈[0.1,0.3)為弱相關,|R|∈[0.3,0.5)為中等相關,|R|∈[0.5,1.0]為強相關得出:PRRSV感染組R2=0.01728,R=0.13,弱相關;PRRSV+PHA組R2=0.0176,R=0.13,弱相關;RRRSV+Dex組R2=0.086,R=0.29,弱相關。
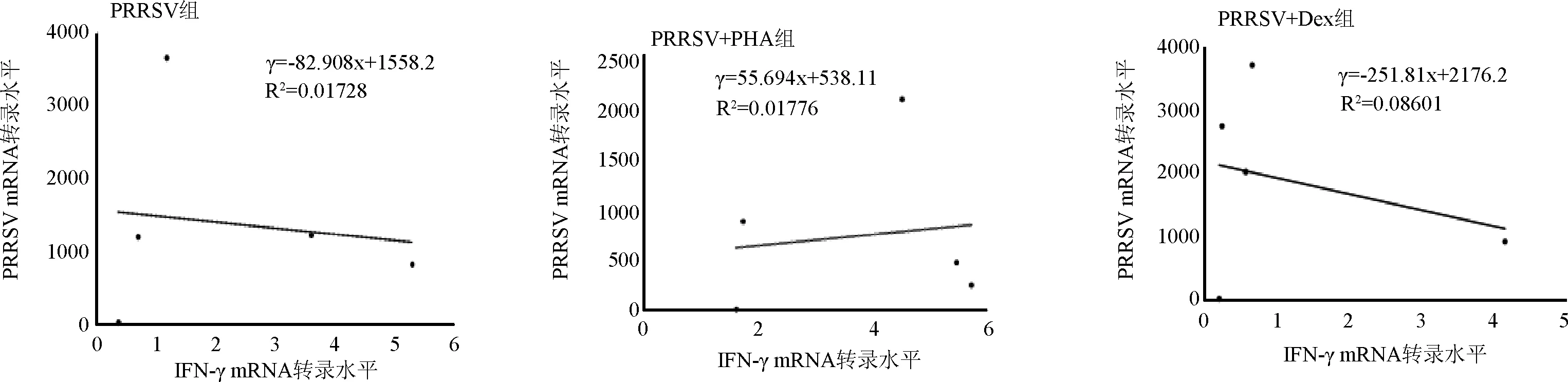
圖3 不同實驗組 IFN-γ與PRRSV轉錄水平相關性分析
3 討論與小結
IFN-γ主要是由抗原、有絲分裂素等刺激、活化的CD4+Th1、CD8+T細胞及NK細胞所分泌的一類可溶性糖蛋白,有較強的抗病毒活性和免疫調節作用,如誘導單核細胞、巨噬細胞、樹突狀細胞、星狀細胞等MHCⅡ類抗原的表達,使其參與抗原遞呈和特異性免疫的識別過程,促進巨噬細胞 FcγR 表達,協同誘導腫瘤壞死因子(tumor necrosis factors, TNF )并促進巨噬細胞殺傷病原微生物[10]。抗原、T細胞促分裂素(PHA)、細胞因子(IL-2、IL-12、IL-18)會誘導IFN-γ的合成,地塞米松、糖皮質激素、IL-10對INF-γ的產生具有抑制作用[11]。因此選取PHA作為陽性對照,地塞米松作為陰性對照。本研究應用q-PCR技術對不同組中IFN-γ的mRNA轉錄水平進行分析,結果發現PRRSV組中,IFN-γ mRNA轉錄量在6~48 h逐漸升高,說明PRRSV可以刺激PAMs分泌IFN-γ。而6 、12 h低于對照組,說明該時期可能處于病毒感染的初期,刺激PAMs分泌IFN-γ的能力較弱,而到了60 h又恢復到對照組水平是因為病毒在PAMs中的增殖,大量的PAMs已脫落、凋亡。以上結果與夏九鮮等人的研究結果相類似[12]。而PRRSV+PHA組和RRRSV+Dex組IFN-γ mRNA轉錄水平分別高于和低于同時間點的PRRSV組,這一結果與T細胞促分裂素(PHA)會誘導IFN-γ的合成,地塞米松對INF-γ的產生具有抑制作用的研究相符[11]。
已有研究表明IFN-γ可以抑制PRRSV 在體外培養的細胞中增殖。本研究應用q-PCR技術對不同組中PRRSV的mRNA轉錄水平進行檢測,結果發現PRRSV+PHA組PRRSV mRNA的轉錄量與相同時間點PRRSV組中PRRSV mRNA的轉錄量相比顯著下調,這一結果與利用IL-12處理PAMs刺激其分泌IFN-γ最終可以降低PRRSV在PAMs中的增殖的結果相類似[8]。RRRSV+Dex組PRRSV mRNA的轉錄量與相同時間點PRRSV感染組相比有所上調,這與佟杰[13]等研究地塞米松可以加強PRRSV在體內的復制,從而使病毒感染造成的損傷加重的結果相類似。另外,本研究結果顯示PRRSV組IFN-γ mRNA轉錄水平與PRRSV轉錄水平存在弱負相關關系,表明PRRSV在PAMs細胞內的增殖復制抑制了IFN-γ的產生,這也是PRRSV導致機體免疫抑制的機制之一。
研究結果對于臨床用藥具有一定的指導意義,結果表明PRRSV感染后可用PHA等藥物或促進劑刺激干擾素的產生,對PRRSV的復制增殖具有一定的抑制作用,這與臨床中用干擾素治療PRRSV豬只的研究類似[14];DEX是糖皮質激素類藥物,雖具有抗炎、抗過敏和抗毒作用,但本研究結果顯示DEX抑制IFN-γ的產生,從而促進了PRRSV的增殖復制,因此PRRSV感染后不建議使用DEX,這與佟杰等[13]連續7 d低劑量注射DEX會造成仔豬機體免疫抑制,增加其死亡率的結果一致。
[1] Edgar R C,Alexel B E,Enric M,etal. Immunological Features of the Non-Structural Proteins of Porcine Reproductive and Respiratory Syndrome Virus[J]. Viruses,2015,7:873-886.
[2] Faaberg K S,Balasuriyam U B R, Brinton A,etal. Family arteriviridae in Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses,London,UK, 2011.
[3] Shyrley P A,Librado C,Irene M,etal. A comparative study of the local cytokine response in the lungs of pigs experimentally infected with different PRRSV-1 strains: Upregulation of IL-1α in highly pathogenic strain induced lesions[J]. Veterinary Immunology and Immunop-athology,2015,164:137-147.
[4] Chen Y,He S,Sun L,etal. Genetic variation, pathogenicity, and immunogenicity of highly pathogenic porcine reproductive and respiratory syndrome virus strain XH-GD at different passage levels[J].Archives of Virology,2015,161(1): 77-86.
[5] Wang R,Zhang Y J. Antagonizing interferon-mediated immune response by porcine reproduc-tive and respiratory syndrome virus[J].Biomed Res Int,2014,(2): 855-866.
[6] Han S K,Song J Y. Gamma Irradiation-reduced IFN-g Expression, STAT1 Signals, and cell-mediated Immunity[J]. Journal of Biochemistry and Molecular Biology,2002,35(6):583-589.
[7] 白雪,趙建增,梁志選,等.IFN-γ研究進展及其在豬繁殖與呼吸綜合征研究中的應用[J]. 中國畜牧獸醫,2010,37(12):145-149.
[8] Quincy L,Rafael E,Rafael E,etal. Interleukin-12 (IL-12) ameliorates the effects of porcine respiratory and reproductive syndrome virus (PRRSV) infection[J]. Veterinary Immunology and Immunopathology,2005,107:105-118.
[9] 佟杰. 下丘腦-垂體-腎上腺軸在高致病性豬繁殖與呼吸綜合征病毒感染仔豬中作用的研究[D]. 北京:中國農業科學院研究生院,2015.
[10]代麗,單安山,孫進華. γ干擾素的研究進展及在畜牧中的應用[J]. 中國畜牧獸醫,2009,36(4):51-57.
[11]Shindler H, Lutz M B,Rollighoff M,etal. The production of IFN-gamma by IL-12 /IL-18 activated macrophages requires STAT4 signaling and is inhibited by IL-4[J].Immuonl,2001,166 (5):3075-3082.
[12]夏九鮮,李銳,周顯珍,等. PRRSV 感染不同時期IFN-γ 表達量分析[J]. 現代農業科技,2013,5:2-76.
[13]佟杰,王剛,于穎,等. 地塞米松對高致病性豬繁殖與呼吸綜合征病毒感染仔豬的影響[J]. 中國預防獸醫學報,2015,37(8):590-594.
[14]胡奕,宋杰,趙寶華. 豬繁殖與呼吸綜合征防控方法研究進展[J]. 動物醫學進展,2009,30(2):69-73.
(編輯:侯向輝)
The Effects of PRRSV Infected PAMs on Transcription Levels of IFN-γinVitro
YAN Xiao-xia, LIU Huan-huan, ZENG Meng-ying, GAO Hong, YAN Yu-lin*
(CollegeofAnimalScienceandTechnology,YunnanAgriculturalUniversity,Kunming, 650201,China)
To explore the effects of porcine reproductive and respiratory syndrome virus (PRRSV) infected pulmonary alveolar macrophage (PAMs) on transcription levels of IFN-γinvitro,three four weeks old piglet free of PRRSV was used in this experiment. The PAMs were isolated aseptically and divided into control group, PRRSV infection group (PRRSV group),PRRSV+PHA group,PRRSV+Dex group and culturedinvitro. The PAMs were collected at 6, 12, 24, 48 and 60 hours post infection. The mRNA transcription of IFN-γ and PRRSV were analyzed by Real-Time PCR. The results shows that the mRNA levels of PRRSV in PRRSV group were gradually increased at 6~24 h and were decreased at 48 h ; the transcription of IFN-γ were gradually increased at 6~48 and significantly higher at 24 and 48 h than control group. PRRSV+PHA group the mRNA levels of PRRSV were slightly lower and the IFN-γ is higher than PRRSV group; RRRSV+Dex group the mRNA transcription of PRRSV were higher but the IFN-γ is lower.The results indicated IFN-γ can inhibit the proliferation of PRRSV in PAMs,the inhibition of IFN -γ is probably one of the mechanisms that PRRSV cause cellular damage.
PRRSV;PAMs;IFN-γ
2016-07-07
A
1002-1280 (2016) 10-0001-05
S852.65
國家自然科學基金項目(31160496,31360599);云南農業大學2015年研究生科技創新項目(2015ykc4)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
火花(2019年12期)2019-12-26 01:00:28
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
人大建設(2019年12期)2019-05-21 02:55:32
汽車工程學報(2017年2期)2017-07-05 08:13:02
學苑創造·A版(2015年11期)2016-01-14 09:03:27
