藻類葉綠素a/葉綠素b型捕光蛋白復(fù)合體結(jié)構(gòu)與功能的研究進(jìn)展
2016-02-22 20:06:42徐冉侯和勝佟少明
天津農(nóng)業(yè)科學(xué) 2016年2期
徐冉++侯和勝++佟少明
摘 要:藻類的Chl a/Chl b型捕光蛋白復(fù)合體,結(jié)合葉綠素a和葉綠素b,主要存在于綠藻、裸藻、輪藻和黃藻中,具有3個(gè)α-螺旋的跨膜區(qū),其中第一、第三螺旋的氨基酸序列在植物界中高度保守,是葉綠素a的結(jié)合位點(diǎn);第二個(gè)跨膜區(qū)的保守的氨基酸序列則為葉綠素b的結(jié)合位點(diǎn)。LHC在捕獲和傳遞光能,光保護(hù)和過(guò)剩能量耗散,調(diào)節(jié)光能在兩個(gè)光系統(tǒng)中的分配,維持類囊體膜的結(jié)構(gòu)等方面起著重要的作用。雖然對(duì)高等植物的LHC的研究日漸深入,但對(duì)藻類捕光色素蛋白復(fù)合體的認(rèn)識(shí)遠(yuǎn)不及高等植物。本文通過(guò)比較藻類Chl a/Chl b型的LHC與高等植物L(fēng)HC結(jié)構(gòu)和功能的異同點(diǎn),可為進(jìn)一步加深對(duì)藻類LHC結(jié)構(gòu)和功能的理解奠定基礎(chǔ)。
關(guān)鍵詞:光合系統(tǒng);捕光蛋白復(fù)合體;光合作用
中圖分類號(hào):Q244 文獻(xiàn)標(biāo)識(shí)碼:A DOI 編碼:10.3969/j.issn.1006-6500.2016.02.006
Research Progress of the Chl a/Chl b Type Light-harvesting Complex Protein in Algae
XU Ran, HOU Hesheng ,TONG Shaoming
(Key Laboratory of Plant Biotechnology of Liaoning Province, School of Life Science, Liaoning Normal University, Dalian, Liaoning 116081,China)
Abstract :Chlorophyll a/chlorophyll blight-harvesting complex (LHC) which can bind two kinds of pigments including chlorophyll a and chlorophyll b mainly exists in the green algae, euglenophyta, charophyta and xanthophyta. It has three α-helixes, which the first and the third helix are highly conserved at the amino acids sequence in the plant kingdom. The first and the third helix contain the binding sites of chlorophyll a, while the second helixes contains the binding sites of chlorophyll b. LHC is the most important apparatus to absorb solar energy and also plays an important role in photoprotection, heat dissipation and in proper energy distribution between the two photosystems. Additionally, LHC plays very important roles in maintaining the thylakoid structure in grana. Although the study about LHC is comprehensive and in-depth, understanding of the structure and function of LHC of the algae is still less than higher plants. The similarities and differences of the structure and function of LHC between various algae and higher plants have been found, but the structure and function of LHC of algae need to be further investigated.
Key words:photosynthetic system; light-harvesting complex; photosynthesis
藻類主要生活在海洋、湖泊與河流中,在水生生態(tài)系統(tǒng)中發(fā)揮著初級(jí)生產(chǎn)者的重要作用,共有約2 100屬,27 000種,可分為綠藻門(mén)、裸藻門(mén)、輪藻門(mén)、黃藻門(mén)、金藻門(mén)、硅藻門(mén)、甲藻門(mén)、褐藻門(mén)、紅藻門(mén)和藍(lán)藻門(mén)等10門(mén)[1]。捕光蛋白復(fù)合體(Light-harvesting complex,LHC)是一類由核基因(Lhc)編碼的蛋白質(zhì)與各類色素結(jié)合,形成的具有各種構(gòu)型的色素蛋白復(fù)合體[2],以單體、二聚體以及其它的聚集狀態(tài)鑲嵌于類囊體膜上,能夠捕獲光能,并把能量迅速傳至反應(yīng)中心(Reaction center ,RC)引起光化學(xué)反應(yīng)。在藻類中,根據(jù)所含LHC種類的不同,又可將10門(mén)藻類分成3大類[2]:(1)葉綠素a(Chl a)/葉綠素b(Chl b)捕光蛋白復(fù)合體型;(2)Chl a/葉綠素c(Chl c)捕光蛋白復(fù)合體型;(3)藻膽體型。本研究將通過(guò)闡述Chl a/ Chl b捕光蛋白復(fù)合體型的藻類與高等植物L(fēng)HC結(jié)構(gòu)和功能的異同點(diǎn),來(lái)進(jìn)一步加深對(duì)藻類LHC結(jié)構(gòu)和功能的理解。
1 LHC色素
與高等植物相比,藻類Chl a/ Chl b捕光蛋白復(fù)合體中Chl b含量較高,Chl a/Chl b比值普遍較低,且因門(mén)類或種類不同而差別較大,從0.6~2.7不等,海洋綠藻的比值最低,一般在0.6~1.0[3],原始綠藻的捕光色素比較特殊,同時(shí)含有Chl a、Chl b及Chl c 3種葉綠素(Chl a/Chl b≈0.7~0.9)以及一種特殊的葉黃素-草綠黃素(Prasinoxanthin),如Micromonadophyceae中的Mantoniella squamata和Micromonaspullsilla。目前,雖然將這類原始綠藻的捕光復(fù)合體歸入Chl a/ Chl b蛋白復(fù)合體中,但也有觀點(diǎn)認(rèn)為它是雜色藻向綠藻進(jìn)化的過(guò)渡態(tài)。雖然不同藻類色素比值均在一定范圍內(nèi),但當(dāng)藻類處于各種脅迫環(huán)境時(shí),其色素的比值與復(fù)合體的構(gòu)型會(huì)發(fā)生不同程度的改變,與之相比,高等植物Chl a/Chl b比值則更為穩(wěn)定,大多在2~3[4]之間。
2 LHC蛋白
綠藻等藻類的PS I和PS II含有各自的捕光蛋白,分別為L(zhǎng)HC I和 LHC II, 由相應(yīng)的核基因(Lhca和Lhcb)編碼,翻譯后的多肽結(jié)合葉綠素和胡蘿卜素等色素插入到類囊體膜上,在不同藻類中,捕光蛋白的結(jié)構(gòu)和生理功能也不盡相同。
2.1 藻類及高等植物的LHCI蛋白
PSⅠ主要由RC和LHC I組成。LHC I含約100個(gè)Chl,Chl a/Chl b≈3~4,具有4條多肽,分子量≈10~24 kDa,其中21 kDa的多肽是PS I長(zhǎng)波長(zhǎng)熒光(730 nm)發(fā)射的來(lái)源,而23 kDa和24 kDa多肽則是短波長(zhǎng)熒光(680 nm)發(fā)射的來(lái)源[5]。
與高等植物相比,藻類的PS I中只有Lhca3編碼的蛋白同系物存在,起著連接LHC I捕光系統(tǒng)和PS I的重要作用。在綠藻中,當(dāng)鐵缺失時(shí),Lhca3的N-末端被識(shí)別并移除,導(dǎo)致LHC I構(gòu)型改變,從而有效減少多余能量從LHC I轉(zhuǎn)移至PS I;在衣藻中,Lhca3基因缺失可引發(fā)所有LHC I蛋白損耗,從而減少光能的吸收與傳遞;在杜氏鹽藻中,鐵脅迫可以引起Lhca3變化,引起LHC I結(jié)構(gòu)改變,從而增大PS I捕光系統(tǒng),平衡光系統(tǒng)中的激感現(xiàn)象[6]。Lhca2和Lhca9是綠藻特有的Lhca基因,其編碼蛋白組成的亞基直接與PS I相連,將多余能量從低聚的LHC I傳遞到PS I。與色素相比,藻類LHC中捕光蛋白的組成更為穩(wěn)定。
2.2 藻類及高等植物的LHC II蛋白
在PS II中,藻類的LHC II蛋白與高等植物具有較高的相似性,均含有由Lhcb1~Lhc6基因編碼的6種多肽與色素非共價(jià)結(jié)合形成的LHC II中的幾種。
在藻類LHCB蛋白家族中,LHCB1含量最多,研究最為深入。每個(gè)LHCBI含有232個(gè)氨基酸殘基,分子量≈28 kDa,具有3個(gè)跨膜α-螺旋,每個(gè)多肽結(jié)合8個(gè)Chl a,6個(gè)Chl b,2個(gè)葉黃素,1個(gè)新黃素和不定量的紫黃素分子,其中兩個(gè)葉黃素分子位于中心,周圍圍繞著緊密結(jié)合的Chl a和Chl b分子,除此之外,每個(gè)LHCB1單體又含有兩個(gè)反式的黃體素(Lut),一個(gè)9-順式新黃質(zhì)(Noe)和一個(gè)不確定的類胡蘿卜素(Car)分子[7],其中Lut與LHCB1的結(jié)合主要用于調(diào)控α螺旋的正確折疊和能量傳遞;強(qiáng)光下,不確定的Car結(jié)合位點(diǎn)結(jié)合紫黃質(zhì)(Vio),在脫環(huán)氧化酶的作用下形成玉米黃素(Zea),利用葉黃素循環(huán)來(lái)抵御光破壞。
成熟的LHCB2與LHCB1具有很高的相似性(約85%以上)[8],其含量在LHCB中僅次于LHCB1,約含228個(gè)氨基酸殘基,分子量為27 kDa,在氨基酸N端有14個(gè)位點(diǎn)與LHCB1不同,其中11個(gè)在基質(zhì)側(cè)、2個(gè)在類囊體囊腔側(cè)、1個(gè)在第一個(gè)跨膜螺旋區(qū),這些結(jié)構(gòu)上的差異導(dǎo)致兩種蛋白質(zhì)功能上的不同,如磷酸化過(guò)程中,LHCB2比LHCB1速率快。
LHCB1和LHCB2在植物體內(nèi)主要以三聚體的形式存在,由于二者是隨機(jī)結(jié)合,所以LHCB2/LHCB1含量的比值會(huì)有所不同,同時(shí)也可以寡聚體的形式存在。例如在黃化植物恢復(fù)轉(zhuǎn)綠的過(guò)程中,二者的三聚體結(jié)構(gòu)就會(huì)轉(zhuǎn)化為寡聚體形式存在。此外,當(dāng)PS II捕光系統(tǒng)變大時(shí),LHCB2/LHCB1含量的比值增加,如低光下,植物葉片的Lhcb2的表達(dá)量是高光強(qiáng)下的3~4倍[9],且Chl b/a比值變大。因此,有人推測(cè)LHCB2可能參與不同光強(qiáng)下的長(zhǎng)期調(diào)控。
LHCB3含有223個(gè)氨基酸殘基,分子量為25 kDa,與LHCB1氨基酸序列相似性在70%以上[10],不參與磷酸化,不易脫離PS II,目前對(duì)于其在體內(nèi)存在的狀態(tài)主要有以下3種觀點(diǎn):(1)與LHCB1和LHCB2一起形成三聚體;(2)獨(dú)自形成三聚體;(3)以單體的形式存在。雖然各自都有一定的實(shí)驗(yàn)證據(jù)支持,但尚無(wú)定論。
LHCB4在綠藻LHCB家族中分子量最大且Vio含量最高,含257個(gè)氨基酸殘基,具有異質(zhì)性。Chl結(jié)合位點(diǎn)為混合位點(diǎn),結(jié)合6個(gè)Chl a ,2個(gè)Chl b。盡管LHCB4與LHCB1的相似性僅為34.3%[11],但在跨膜螺旋區(qū),結(jié)合色素的配體和掩埋離子對(duì)的殘基是相對(duì)保守的。LHCB4與其它LHCB多肽的主要差異是LHCB4的N端有42個(gè)親水氨基酸殘基插入,并在此區(qū)含有磷酸化位點(diǎn)。
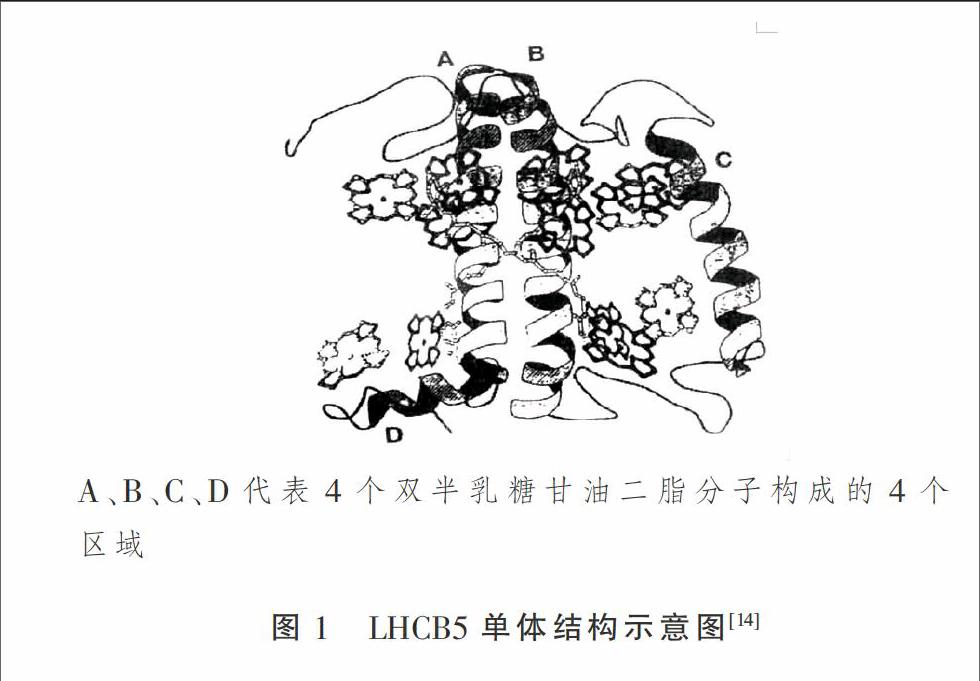
LHCB5含有247個(gè)氨基酸殘基,結(jié)合6個(gè)Chl a,3個(gè)Chl b,與LHCB1的相似性約為48.7%[12],他們的螺旋A區(qū)和C端結(jié)構(gòu)非常相似,主要差異在螺旋C區(qū)(圖1),此部分的相似性很低。LHCB5結(jié)合有Cu2+,因此,推測(cè)其可能是葉黃素循環(huán)中的氧化酶之一。在LHCBl和LHCB2缺乏的植物中,LHCB5可以代替它們和LHCB3形成三聚體結(jié)構(gòu),并且還能形成C2S2M2型[13](C:反應(yīng)中心;S:與C結(jié)合緊密的三聚體;M:與C結(jié)合程度中等的LHCII)LHC ll-PS II超復(fù)合體結(jié)構(gòu),但LHCB5缺少磷酸化的功能,所以該植物不能在強(qiáng)光下生長(zhǎng),說(shuō)明LHCB5僅能在一定程度上代替LHCB1和LHCB2的功能。
LHCB6是LHCB中較小的蛋白,含210個(gè)氨基酸殘基,結(jié)合6個(gè)Chl a和4個(gè)Chl b分子,與其他LHCB相比,LHCB6與LHCB1相似性最低,約為29.2%[15],其蛋白構(gòu)象缺少暴露在囊腔的C端和螺旋D區(qū),這使其結(jié)合的色素在提取中極易丟失,但Chl b可起到穩(wěn)定其結(jié)構(gòu)的作用。LHCB6缺乏時(shí),植物PS Ⅱ捕光受到干擾,與PS Ⅱ的RC之間的連接減弱,使類囊體膜上的微觀組織發(fā)生變化,光合效率降低,但類囊體膜中的色素和蛋白含量幾乎不變。因此,LHCB 6 對(duì)于PSⅡD的結(jié)構(gòu)和功能都非常重要[16]。
除此之外,LHCB7~LHCB9只在極個(gè)別物種中存在。多數(shù)藻類中含有與高等植物L(fēng)HCB4~6功能相似的多肽,且在跨膜α-螺旋處具有高度保守性,但是分子量大小不同,也正是這些微小差別決定了它們?cè)诠δ苌系奶禺愋裕缛R茵衣藻、鹽生杜氏藻等。
3 LHC蛋白的功能
3.1 光能的捕獲及傳遞
研究表明,在PS I中,類胡蘿卜素分子吸收光能后迅速將其傳遞給周圍的7個(gè)Chl a分子,然后能量在Chl a與Chl b分子之間傳遞,直至達(dá)到穩(wěn)定平衡。
在PSⅡ中,每個(gè)單體結(jié)合220~230[17]個(gè)捕光色素分子,多數(shù)以二聚體形式存在。LHCⅡ?qū)@得的能量經(jīng)過(guò)復(fù)雜的過(guò)程傳遞至RC并以穩(wěn)定化學(xué)能的形式儲(chǔ)存起來(lái)。由于能量是在Chl a和Chl b分子間傳遞,因此LHCⅡ所發(fā)出的熒光由二者共同發(fā)射。
3.2 激發(fā)能分配的調(diào)控
LHCB蛋白通過(guò)磷酸化和脫磷酸化參與PS I與PS II間的激發(fā)能分配,此分配過(guò)程由質(zhì)體醌(Plastoquinone, PQ)庫(kù)的氧化還原狀態(tài)控制的激酶來(lái)調(diào)節(jié)。
當(dāng)PQ庫(kù)處于還原狀態(tài)且與細(xì)胞色素b6/f(Cytb6f)的Qo位點(diǎn)結(jié)合時(shí),磷酸激酶活化,催化LHCB蛋白磷酸化,使其從富含PS II的基粒膜區(qū)遷移到富含PS I的間質(zhì)膜區(qū)或基粒的邊緣膜區(qū),使能量利于向PS I的分配;當(dāng)PQ庫(kù)處于氧化狀態(tài)時(shí),磷酸激酶被鈍化,磷酸化的LHCB蛋白脫磷酸化,遷移的LHCB重新回到基粒膜區(qū),使能量利于向PS II的分配[18]。在遷移過(guò)程中,LHCB可以以單體、二聚體、三聚體和寡聚體形式存在。在紅藻中,盡管有一些蛋白可以磷酸化,但這些蛋白并不參與激發(fā)能分配的調(diào)節(jié)。
3.3 能量的耗散與光保護(hù)
研究發(fā)現(xiàn),藻類主要通過(guò)降低能量捕獲和消耗過(guò)多能量這兩種機(jī)制來(lái)避免強(qiáng)光對(duì)自身的損傷,如強(qiáng)光下,一些Lhc基因表達(dá)量會(huì)不同幅度下降;而在非輻射耗散(Nonphotochemical dissipation, NPD)中則主要通過(guò)LHC蛋白參與的復(fù)雜機(jī)制來(lái)實(shí)現(xiàn)。
NPD通過(guò)質(zhì)子化過(guò)程改變色素分子的位置及其附近的電荷密度誘導(dǎo)能量的傳遞以達(dá)到熒光淬滅的目的,LHCB4~6由于含Zea和Vio較多而參與其中,主要通過(guò)降低激發(fā)單線態(tài)的最低能量或刺激LHC II的聚集狀態(tài)來(lái)提高能量的耗散 [19]。
4 結(jié)論與展望
目前,對(duì)于藻類捕光蛋白色素復(fù)合體的研究具有明顯的不均衡現(xiàn)象,一些Chl蛋白復(fù)合體,如原綠藻LHC中的某些多肽,已經(jīng)在空間結(jié)構(gòu)、能量傳遞及分子生物學(xué)方面取得了許多重要的成果[20],但對(duì)大多數(shù)Chl a/ Chl b捕光蛋白復(fù)合體型藻類LHC的整體特點(diǎn),包括多肽的組成、相互關(guān)系以及色素的結(jié)合情況等卻并不清楚,相信隨著研究手段的不斷提高,在不久的將來(lái)這一領(lǐng)域定會(huì)獲得突破性的進(jìn)展。
參考文獻(xiàn):
[1] XIA B M. Checklist of marine algae of China[M]// Liu R Y. Checklist of marine biota of China seas.Beijing: Science Press,2008:35-40, 97-107, 252-284.
[2] JONATHAN A. Structural and functional diversification of the light-harvesting complexes in photosynthetic eukaryotes[J].Photosynth Res, 2010,106:57-71.
[3] WOLF G R, CUNNINGHAM F X, GRABOWSKI B, et al. Isolation and characterization of photosystems I and II from the red alga Porphyridium cruentum[J]. Biochim Biophys Acta, 1994, 1188: 357-366.
[4] KOZIOL A G, BORZA T, ISHIDA K, et al.Tracing the evolution of the light-harvesting antennae in chlorophyll a /b-containing organisms[J]. Plant Physiol, 2007,143: 1802-1816.
[5] DION G D, JULIE A P. Light-harvesting complex gene expression is controlled by both transcriptional and post-transcriptional mechanisms during photoacclimation in Chl amydomonas reinhardtii[J]. Plant Physiol, 2003, 118:193-205.
[6] QIN X, SUGA M, KUANG T, et al. Photosynthesis. Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex[J]. Science, 2015, 348(6238):989-995.
[7] LUCI?SKI R, JACKOWSKI G. The structure, functions and degradation of pigment-binding proteins of photosystem Ⅱ[J]. Acta Biochimica Polonica, 2006, 53(4):693-708.
[8] ELISABETTA C, CATHY Y. Coherently wired light-harvesting in photosynthetic marine algae at ambient temperature[J]. Nature,2010,463:644-647.
