可可廣翅蠟蟬的行為節律觀察
2016-03-01 12:58:46范廣玉劉寧國劉學敏陳文龍
貴州農業科學 2016年4期
關鍵詞:生物學
范廣玉, 劉寧國, 楊 群, 劉學敏, 陳文龍
(貴州大學 昆蟲研究所, 貴州山地農業病蟲害重點實驗室, 貴州 貴陽 550025)
可可廣翅蠟蟬的行為節律觀察
范廣玉, 劉寧國, 楊 群, 劉學敏, 陳文龍*
(貴州大學 昆蟲研究所, 貴州山地農業病蟲害重點實驗室, 貴州 貴陽 550025)
為進一步做好可可廣翅蠟蟬的預測預報及防治工作,采用室內飼養觀察的方法對其羽化、交配、產卵的行為及節律進行研究,并對其田間產卵特征進行調查。結果表明:可可廣翅蠟蟬成蟲羽化發生在晚上和凌晨,白天未見羽化;成蟲交配主要發生在白天,晚上很少交配,凌晨未見;成蟲全天均可產卵,高峰期集中在上午與凌晨;田間主要將卵產在莖粗為1.5 mm左右的嫩莖上,產卵深度約0.9 mm,產卵刻痕主要集中在距頂梢3 cm范圍,平均每刻痕卵量約為11粒。
可可廣翅蠟蟬; 羽化; 交配; 產卵; 行為節律
生物的生理活動及外在行為具有一定的周期節律特征[1-4]。其晝夜節律具有種的特異性,這不僅是昆蟲個體間基因交流的基礎,也是生產上進行預測及防治的重要依據[5-8]。近年來,隨著氣候條件、茶園生態環境、茶樹品種及其栽培方式的改變,茶樹蠟蟬種群正逐漸從次要害蟲變為主要害蟲[9],截止目前,有關蠟蟬種類如柿廣翅蠟蟬(RicaniasublimbataJacobi)、八點廣翅蠟蟬(R.speculumWalker)、碧蛾蠟蟬(GeishadistinctissimaWalker)、白蛾蠟蟬(LawanalimitataMelichar)、緣紋廣翅蠟蟬(R.marjinlisWalker)等的行為節律均已有相關報道[10-33]。
可可廣翅蠟蟬(RicaniacacaonisWalker)在貴州的具體發生及危害狀況尚未見報道,筆者通過調查發現,該蟲在貴州都勻市、貴陽市均有發生,其危害程度每年呈上升趨勢,尤其在貴陽黔陶地區危害嚴重。因此,為做好該蟲預報及防治工作,探明其行為節律意義重大。邵元海等[9,34-35]對該蟲的生態學及防治進行報道,但對其生物學及行為節律研究較少,且多局限于江蘇地區,而地域性差異致使該蟲在不同地區的生活習性必然有所不同。因此,探明可可光翅蠟蟬在貴州地區的生物學及生態學特性是實現當地對該蟲的預測預報及有效防治的重要途徑,而其行為節律更是重中之重。為此,筆者于2013年采用室內飼養觀察結合田間調查的方法對其羽化、交配、產卵的行為及節律進行研究,以期為該蟲的預測預報及有效防治提供理論依據。
1 材料與方法
1.1 研究對象
以可可光翅蠟蟬為研究對象,蟲卵采自貴陽市花溪熙皇趙司貢茶茶場。
1.2 試驗時間及地點
試驗于2013年3—10月在貴州大學昆蟲研究所與貴陽市花溪熙皇趙司貢茶茶場(海拔1 300~1 500 m的山坡上,新老茶園共2 00 hm2)進行。
1.3 成蟲的行為節律觀察
1.3.1 成蟲羽化 在若蟲孵化前,將剪自茶園的帶卵枝條基部用濕棉團包裹,置于300 mL的透明塑料杯中,杯口用紗網封蓋,待若蟲孵化后使用嫩茶枝進行單頭飼養,記錄蛻皮次數,對蛻皮至末齡的若蟲進行不定時觀察。自有蟲羽化開始,每2 h觀察1次,記錄每次觀察所發生羽化的蟲口數量,并對其羽化行為進行拍照描述,共100次重復。
1.3.2 成蟲交配 將采自茶園的雌、雄成蟲進行配對,置于裝有茶樹嫩枝的300 mL塑杯進行飼養,每杯1對,共50次重復。嫩枝基部用濕棉團包裹并對茶枝進行不定期更換,以保證枝葉新鮮。每2 h觀察1次,記錄每次觀察發生交配的蟲口對數,并對其交配行為進行拍照描述。
1.3.3 雌成蟲產卵 1) 室內行為習性觀察。將已發生交配的雌成蟲置于300 mL的塑杯進行單頭飼養,取直徑1.5 mm左右的茶樹嫩枝,將葉片剪除(以便于觀察產卵行為),其基部用足夠大的濕棉團包裹,置于杯底并固定,供成蟲產卵;將剪取的葉片取2~3片置于杯中,對葉片進行每日更換以保證新鮮,供雌成蟲取食。每日觀察其產卵情況,自有蟲產卵開始,每3 h觀察1次,記錄每次觀察發生產卵的蟲口數,并對其產卵行為進行拍照描述。2) 野外產卵特征調查。于產卵期過后,取一塊茶樹種植密度大、生長年齡長的樣地,按Z字取樣法抽取30株茶樹,每株隨機抽取1枝具3~4個分支的枝條,記錄每枝條的產卵刻痕數,每刻痕所在部位,每刻痕距枝條頂梢距離,每刻痕深度,每刻痕卵粒數,每卵粒間距。
1.4 數據統計與分析
試驗數據分析應用軟件為EXCEL 2003及SPSS 18.0。
2 結果與分析
2.1 成蟲羽化(封二圖Ⅰ)
當5齡若蟲到達末期準備發生羽化時,胸背裂開一條較窄縫隙,但并未完全裂開,裂縫處仍有一層薄膜相連,此時若蟲仍有運動能力,但常棲居于一處靜止不動,等待羽化。在發生羽化時,其腹部伸直拉長,頭部頂殼裂開,其頭部慢慢自裂縫處鉆出。之后隨著腹部不斷的進行前后蠕動,裂縫不斷加深,沿胸背中脊向胸背末端逐漸延伸,直至腹部第二腹節處停止,在此過程中其翅隨著胸背的露出而逐漸顯化,前、中、后足亦慢慢自原來體殼中抽出。當翅剛顯化出來時并未伸展,其近外緣處向下彎曲,折疊于胸部兩側,形似蘑菇。待后足完全抽出后,腹部暫停蠕動約30 s,之后前、中、后足開始慢慢擺動,前足向前方伸展,超過頭部,中足做前后重復性擺動,后足向兩側張開,呈150°~180°角,并且上翹頂住翅的邊緣,促進翅的伸展,隨之整個軀體自原來體殼中爬出。
以上整個過程,從頭部頂殼裂開到整個軀體鉆出體殼,需經40~60 min,此后再經10~20 min,翅慢慢伸展開來。至此,整個羽化過程結束。剛羽化的成蟲通體乳白,前、后翅均透明,翅脈不甚清晰,亦不見斑紋、刻點,此時成蟲常停留在皮殼前方,不進行運動取食,之后隨時間推移,前翅顏色逐漸加深,由透明的白色逐漸轉為較透明的灰色,進而轉為不透明的黑灰色或黃褐色,翅脈逐漸清晰,其翅上的各種斑紋和刻點亦慢慢顯化,而后翅亦由原來的白色透明轉為淺灰色透明。成蟲從剛羽化完成到發展為骨骼堅硬、能飛善蹦的成蟲約需要7~10 h。
從圖1可見,可可廣翅蠟蟬羽化均發生在晚上和凌晨,白天不發生羽化。其中,羽化高峰期集中在晚上22:00~00:00及凌晨00:00~02:00,二者之間無顯著差異,但顯著高于其他時間段;其次為20:00~22:00與02:00~04:00,而18:00~20:00及04:00~06:00羽化量較少。
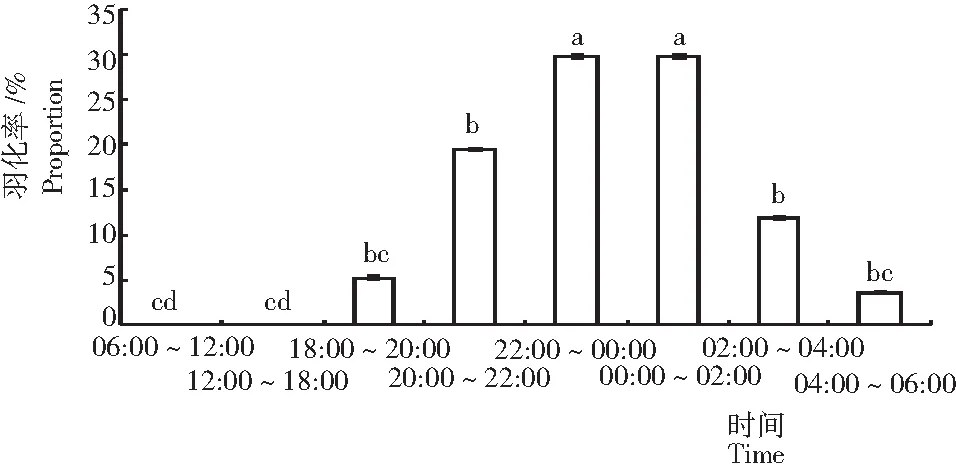
注:圖中小寫字母代表α=0.05水平的顯著性差異(Duncan法)(下同)。
Note: Lowercase letters in the Fig. indicate significance of difference at 0.05 level. The same below.
圖 1 可可廣翅蠟蟬的羽化節律
Fig.1 Emergence rhythm ofR.cacaonis
2.2 成蟲交配(封二圖Ⅱ)
在進行交配時,雌、雄蟲身體以尾節為頂角保持140°~180°,雄蟲尾節向一側彎曲,之后抱器張開,緊握雌蟲第九腹板基部的凹陷處,為其交配提供著力點,此時陽莖外露。雄蟲通過雌蟲交配孔使陽莖插入雌蟲體內,此交配孔位于雌蟲第7與第8腹節之間的節間膜處,陽莖經過不斷抽插后直至交配囊,此時雄蟲尾節后上角便抵在雌蟲交配孔旁邊的節間膜處,當雄蟲陽莖停止抽插后,其肛節緊扣雌蟲第7腹板,并可見雄蟲肛刺突輕微晃動。可可廣翅蠟蟬在交配過程中如遇驚擾,可進行小范圍的爬行或跳躍,此后交配行為仍然繼續。在交配即將結束時,雌蟲身體進行輕微的左右晃動,后足凌空輕踢,身體向前爬行,此時尾節兩側腹板張開,促使雄蟲將陽莖拔出體外,約1~3 min后雄蟲抱器松開,陽莖自雌蟲體內抽出,雄蟲抱器與肛節各歸原位,雌蟲尾節兩側腹板亦重新閉合,至此交配行為結束,整個交配過程約需40~70 min。
從圖2可見,可可廣翅蠟蟬成蟲的交配主要發生在白天,晚上很少有蟲交配,而在凌晨未發現有蟲交配,交配高峰期發生在上午08:00~12:00,且以08:00~10:00居多,顯著高于其他時間段。下午以14:00~16:00發生交配的蟲口數居多,但與12:00~14:00及16:00~18:00時間段之間差異不顯著。
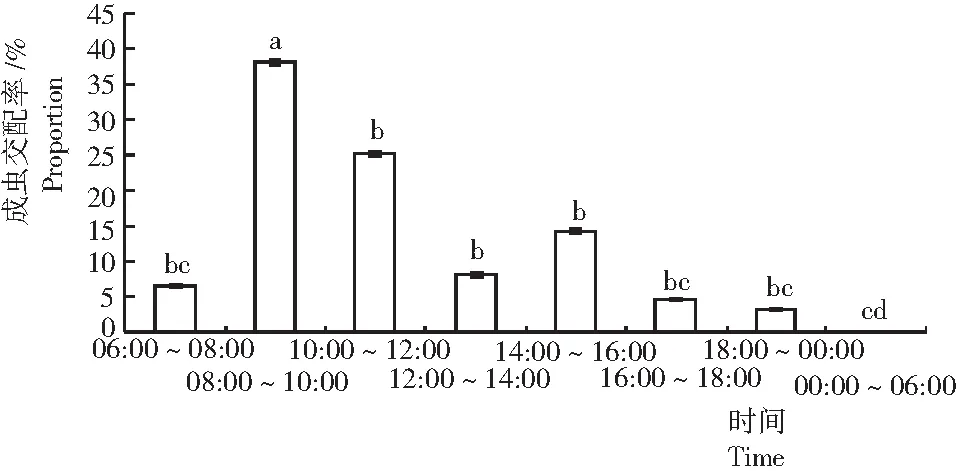
圖2 可可廣翅蠟蟬的交配節律
2.3 產卵(封二圖Ⅲ)
2.3.1 產卵行為及節律觀察 可可廣翅蠟蟬雌蟲自交配至產卵所需時間浮動較大,短則需要2 d,長則需要5 d,其產卵須經刻槽、產卵、掩埋3個階段。在產卵時腹部末端彎曲下垂,與產卵枝條垂直,第九腹板張開,第1、2產卵瓣外露,形如利刀,兩對產卵瓣交替鉆刻和切割枝條,一邊切割一邊將木屑挖出,最終形成能夠盛卵的傾斜刻槽,角度約45°。待形成的刻槽適合盛卵時產卵瓣便停止切割,此時腹部第九節兩側腹板張開將枝條夾住,接著腹部不斷地收縮與舒張,將卵通過產卵孔排出體外,斜產于凹槽之內,卵的尖端朝上,膨大一端朝下。待將卵盛好后第九腹板松開枝條,產卵瓣將挖出的木屑覆于卵上,至此整個產卵過程結束。產卵需10~13 min/粒, 待一顆卵產完之后蟲體向前爬行約 0.8 mm 繼續產卵,最終形成魚鰭狀的條形刻痕。
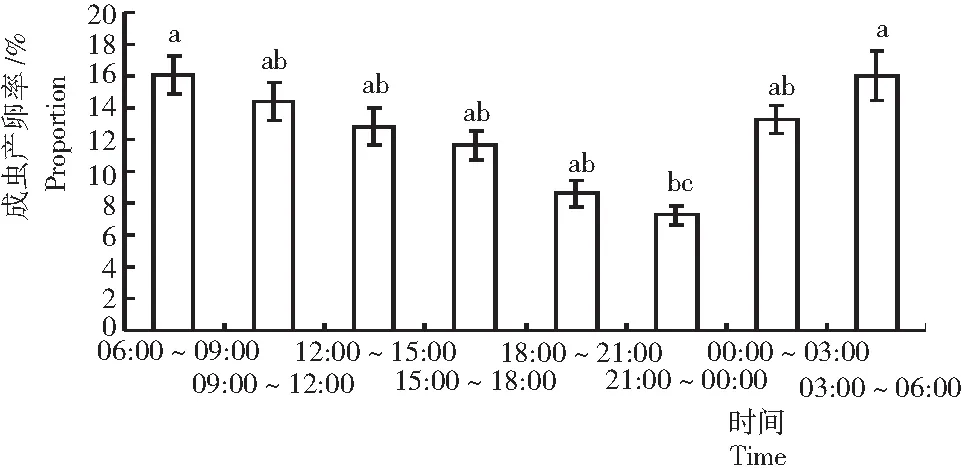
圖3 可可廣翅蠟蟬的產卵節律
從圖3可見,可可廣翅蠟蟬全天均可產卵,產卵高峰期集中在上午與凌晨,占整日產卵量的60%以上;自早晨06:00至晚上00:00產卵量逐漸降低,之后進入凌晨后產卵量又逐漸升高。
2.3.2 野外產卵特征 可可廣翅蠟蟬的產卵刻痕呈條形,為土黃色,每卵粒間呈魚鰭狀排列。從表1可見,每枝條平均刻痕數為2~4個,而分布于莖上的刻痕數為2~3個,為葉片上刻痕數的5~8倍。對于莖上的產卵刻痕,平均每刻痕含11~13粒卵,卵粒間距約為0.73 mm;對于葉片上的產卵刻痕,平均每刻痕含7~9粒卵,其卵粒數少于莖上刻痕,但其產卵粒間距約為1.00 mm左右,大于莖上刻痕的卵粒間距。
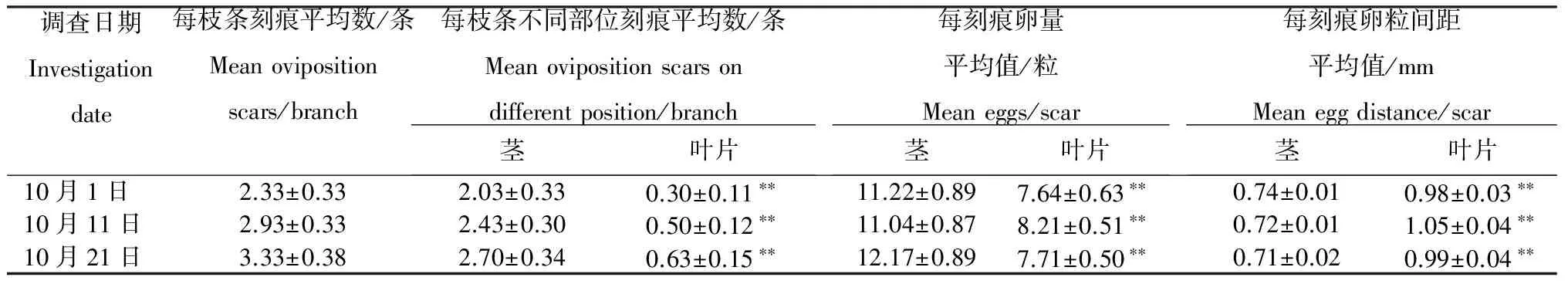
表1 可可廣翅蠟蟬的產卵特征
注:*表示經t-檢驗差異顯著(P<0.05), **表示經t-檢驗差異極顯著 (P<0.01)。
Note: * and ** indicates significance of difference atP<0.05 andP<0.01 level respectively.
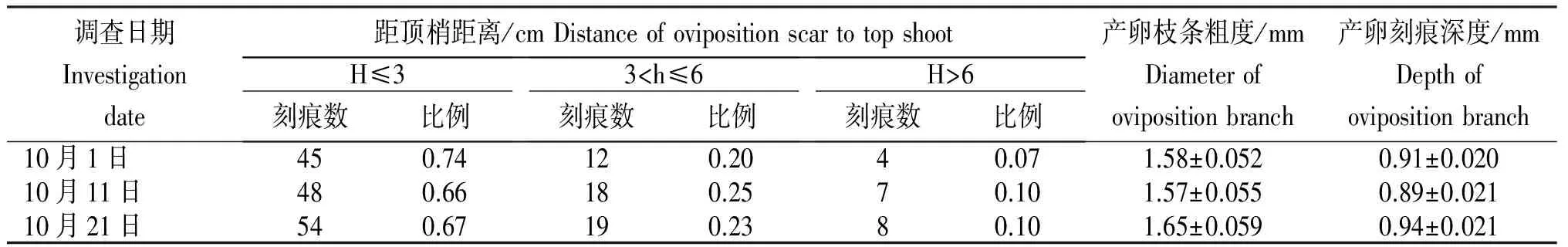
表2 莖上可可廣翅蠟蟬產卵刻痕的特征
由于可可廣翅蠟蟬主要產卵于枝條嫩莖上,并且危害巨大, 所以進一步對該刻痕進行深入調查。 從表2可見,該蟲產卵時選擇的枝條粗度在1.5 mm左右,主要為當年抽發的已發生木質化或半木質化的較嫩枝條,其產卵深度約為0.9 mm,可深達木質部,嚴重影響枝條養分的運輸。產卵刻痕主要分布在距梢頂≤3 cm的枝條上,占總刻痕數的65%~75%;其次為3~6 cm,占總刻痕數的20%~25%;而在>6 cm的枝條上產卵刻痕數很少, 僅在10%左右。
3 結論與討論
通過對可可廣翅蠟蟬3種行為節律的觀察發現,成蟲羽化主要集中在晚上和凌晨,白天不發生羽化,羽化高峰期集中在22:00~02:00,接近羽化總量的60%。成蟲交配主要集中在白天,上午較多,占總交配對數的65%以上,且以08:00~10:00最多,可達35%~40%。雌成蟲交配后短則2 d,長則5 d即可產卵,雌蟲全天均可產卵,其產卵高峰期集中在上午與凌晨,占總產卵量的60%以上。
通過田間調查發現,該蟲可將卵產于粗度為1.5 mm左右的嫩莖及葉片背面主脈上,但以嫩莖上的產卵刻痕數居多,約為葉片主脈的5~8倍。莖上每刻痕的平均卵量多于葉片,且卵粒密度也大于葉片,造成此種現象的原因可能是嫩莖直徑大于葉片主脈,能夠更好地對卵粒進行掩埋,從而提供優越的保護場所,易于越冬。對于嫩莖上的產卵刻痕,其刻痕平均深度為0.9 mm,主要分布在距頂梢3 cm以內的枝條上,約占總刻痕數量的65%~75%,造成此種刻痕分布的主要原因可能是越靠近頂梢,其莖干越嫩,不但易于成蟲刺吸取食,更易于該蟲產卵器劃破枝條進行產卵。
本文對可可廣翅蠟蟬的羽化、交配和產卵節律進行時間上的量化分析,并對其行為過程進行觀察和描述,這雖然對該蟲的田間防治及進一步科學研究均具有重要意義,但多局限于現象描繪,因此在下一步工作中,應對其內在行為機制進行深入研究,從而探明各行為的生理意義。
[1] 涂小云,陳元生.蛾類昆蟲行為節律[J].生物災害科學,2013,36(1):18-21.
[2] 吳少會,向 群,薛芳森.昆蟲的行為節律[J].江西植保,2006,29(4):147-157.
[3] Pittendrigh C S.Temporal organization:reflection of Darwinian watcher[J].Annual Review of Physiology,1993,55:17-54.
[4] Takahishi J S.Molecular neurobiology and genetics of circadian rhythm in mammals[J].Annual Review of Neuroscience,1995,18:3531-3553.
[5] 秦玉川.昆蟲行為學導論[M].北京:科學出版社,2008.
[6] 張清泉,張雪麗,陸 溫,等.昆蟲交配行為、繁殖適度和性信息素在國內的研究進展[J].廣西農業科學,2009,40(2):164-168.
[7] 劉吉敏,黃其椿,檀志全,等.龍眼樹荔枝蝽越冬成蟲發生規律及交配節律[J].南方農業學報,2012,43(8):1135-1138.
[8] 孫麗娟,戴華國,衣維賢,等.二化螟水稻類群與菱白類群成蟲羽化節律和交配節律研究[J].昆蟲知識,2002,39(6):421-423.
[9] 徐德良.茶樹蠟蟬種群生態及控制技術研究[D].蘇州:蘇州大學,2009.
[10] 陳彩賢.三斑廣翅蠟蟬生活習性觀察[J].廣西農業科學,1998(3):139-141.
[11] 鄧晗嵩.同翅目蛾蠟蟬科(f-tomopter:flatidae)生活習性及防治研究[J].黔西南民族師范高等專科學校學報,2007(4):122-124.
[12] 董文玲.白蛾蠟蟬生物學特性及防治研究[J].林業調查規劃,2006,31(A02):159-161.
[13] 李慧玲,劉豐靜,王定鋒,等.白蛾蠟蟬的發生和防治[J].茶葉科學技術,2012(4):24-25.
[14] 何 娟,于瑋臺,陳文龍.貴陽地區茶樹碧蛾蠟蟬的發生規律及防治[J].貴州農業科學,2013,41(7):105-107.
[15] 劉永生,胡 波,張清良,等.八點廣翅蠟蟬生物學特性與防治初報[J].湖北林業科技,1999(2):29-30.
[16] 劉永生,張清良.柿廣翅蠟蟬生物學特性及防治初報[J].山西果樹,2000(4):5-6.
[17] 李蘇萍,陳秀龍,韓國柱,等.山東廣翅蠟蟬生物學特性及防治措施[J].中國森林病蟲,2006,25(3):36-38.
[18] 羅曉明,羅天相,劉 莎.柿廣翅蠟蟬的發生與防治[J].河南農業科學,2014(3):41-42.
[19] 劉朝萍.柑橘白蛾蠟蟬的發生與防治技術[J].植物醫生,2009,22(5):25-25.
[20] 梅志堅.茶樹碧蛾蠟蟬的發生與防治[J].茶葉科學技術,2004(3):43-43.
[21] 沈 強,王菊英,柳建定,等.山東廣翅蠟蟬的生物學特性及防治[J].昆蟲知識,2007,44(1):116-118.
[22] 江偉林.山東廣翅蠟蟬在蓮都地區的發生及綜合防治[J].浙江柑橘,2013(1):35-36.
[23] 汪榮灶,祝 婕.茶廣翅蠟蟬生物學初步研究[J].中國茶葉,2010(5):29.
[24] 于瑋臺,陳文龍.碧蛾蠟蟬行為習性研究[J].山地農業生物學報,2013,32(2):123-127.
[25] 喻愛林,單繼紅,涂業茍,等.油茶高產無性系碧蛾蠟蟬的生物學特性及防治[J].江西植保,2006,29(4):181-182.
[26] 喻愛林.油茶八點廣翅蠟蟬的生物學特性及防治[J].江西林業科技,2007(3):34-35.
[27] 趙豐華,呂立哲,任紅樓.信陽茶樹新害蟲——蠟蟬[J].中國茶葉,2010(10):16-17.
[28] 肖鐵光,周社文,王文藝,等.V脊菱蠟蟬生物學特性和消長規律初步研究[J].湖南農業大學學報:自然科學版,2006,32(1):70-72.
[29] 趙豐華,呂立哲,任紅樓,等.豫南茶園柿廣翅蠟蟬生物學特征[J].中國茶葉,2011(5):18-19.
[30] 顧昌華.白斑廣翅蠟蟬的生物學特性[J].貴州農業科學,1997,25(5):50-51.
[31] 柴正群,楊發軍,朱建青,等.云南普洱茶樹碧蛾蠟蟬的危害與防治[J].現代園藝,2012(21):64-64.
[32] 陳慶紅.柿廣翅蠟蟬生物學特性及其防治初步研究結果[J].湖北植保,2000(3):24-25.
[33] 譚萬忠,李 培,張克誠,等.重慶地區茶樹病害種類及武夷菌素對茶病的田間控制效果[J].西南大學學報:自然科學版,2008,30(4):123-127.
[34] 邵元海,熊 輝.可可廣翅蠟蟬的生物學特性及防治[J].蠶桑茶葉通訊,2009(5):38-39.
[35] 邵元海,周靜峰,徐德良.可可廣翅蠟蟬若蟲空間分布型及抽樣技術研究[J].茶葉,2009,35(4):219-220,223.
(責任編輯: 王 海)
Behavior Rhythm ofRicaniacacaonis
FAN Guangyu, LIU Ningguo, YANG Qun, LIU Xuemin, CHEN Wenlong*
(InstituteofEntomology,GuizhouKeyLaboratoryforPlantPestManagementofMountainousRegion,GuizhouUniversity,Guiyang,Guizhou550025,China)
The behavior and rhythm of emergence, mating and oviposition ofR.cacaoniswere observed by indoor raising method and the oviposition feature in field was investigated to further complete forecasting and control ofR.cacaoniswell. Results: The emergence ofR.cacaonisadults occurs at night and before dawn. The mating behavior ofR.cacaonisadults happens in the day mainly.R.cacaonisadults lay eggs whole day and the peak time concentrates in the morning and before dawn.R.cacaonisadults mainly lay eggs in young stems with 1.5 mm diameter in field and the oviposition depth is about 0.9 mm. The oviposition scars mainly concentrate in 3 cm range to top shoot and the average eggs per scar is about 11.
Ricaniacacaonis; emergence; mating; oviposition; behavior rhythms
2015-09-08; 2015-12-30修回
貴州省科技廳農業攻關項目“茶樹重要病蟲害無公害防除技術研究與推廣”[黔科合字NY字(2010)3024]
范廣玉(1987-),男,在讀碩士。研究方向:動物生態及害蟲綜合治理。E-mail:383441879@qq.com
*通訊作者:陳文龍 (1965-),男,教授,碩士生導師,從事植物病蟲害研究。E-mail:CWL001@163.com
1001-3601(2016)04-0158-0053-04
S433
A
猜你喜歡
陜西學前師范學院學報(2022年10期)2022-10-31 14:23:44
中國飼料(2021年17期)2021-11-02 08:15:10
現代畜牧科技(2021年9期)2021-10-13 06:38:54
現代畜牧科技(2021年4期)2021-07-21 06:13:00
生物學通報(2021年5期)2021-03-16 06:42:16
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
數學物理學報(2017年2期)2017-06-05 09:12:30
中國組織化學與細胞化學雜志(2016年4期)2016-02-27 11:16:07
醫學研究雜志(2015年11期)2015-06-10 06:44:03
