昆蟲溫度耐受性評價方法的研究進展
2016-03-02 01:50:28李鴻波何永福
貴州農(nóng)業(yè)科學 2016年1期
李鴻波, 何永福
(貴州省植物保護研究所, 貴州 貴陽 550006)
昆蟲溫度耐受性評價方法的研究進展
李鴻波, 何永福*
(貴州省植物保護研究所, 貴州 貴陽 550006)
昆蟲屬變溫動物,對環(huán)境溫度具有很強的依賴性。昆蟲對溫度脅迫的適應(yīng)能力直接決定其種群動態(tài)和地理分布。為科學評價昆蟲的溫度耐受性,對已報道的有關(guān)昆蟲溫度耐受性評價方法進行全面總結(jié),并對其優(yōu)缺點進行評價。
溫度耐受性; 評價方法; 昆蟲
昆蟲是變溫動物,其生命活動受到溫度(高溫和低溫)、濕度、光照(輻射)、重金屬、食物、寄生及捕食等諸多生物和非生物因子的影響。其中,溫度是最基礎(chǔ)的環(huán)境因子之一。在時間和空間范圍,其直接和間接地決定昆蟲的生態(tài)學和進化等[1]。如,溫度影響昆蟲的免疫功能、交配、繁殖以及生長[2],同時,溫度也是造成昆蟲死亡的重要因子,昆蟲對溫度的適應(yīng)能力決定其分布和種群動態(tài)[3]。
在自然界,環(huán)境溫度是多變的,這對昆蟲的繁衍構(gòu)成極大威脅。昆蟲為了能適應(yīng)環(huán)境,必須具備一定的耐受能力,否則,將面臨滅亡。全球氣候變化除包括極端高溫外,還包括極端低溫,而且二者出現(xiàn)的頻率增多。因而極端溫度對昆蟲種群的影響,昆蟲應(yīng)對極端溫度的行為進化與適應(yīng)成為生態(tài)學與進化生物學領(lǐng)域研究的核心。昆蟲溫度耐受性的研究已有數(shù)十年的歷史,評價方法也有多種,所得的結(jié)果也不盡相同。
總體而言,這些評價方式可以分為2類:一類是將昆蟲直接暴露脅迫溫度下(急劇式),二是通過緩慢的升溫或者降溫暴露到脅迫溫度下(斜坡式)[4-6]。為科學地評價昆蟲的溫度耐受性,作者對其評價方法進行了全面的總結(jié)。
1 昆蟲耐寒性的評價方法
1.1 昆蟲過冷卻點的測定
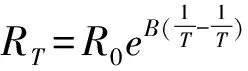
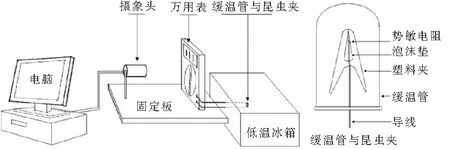
圖1 測定昆蟲過冷卻點的簡易裝置
盡管多位學者對測定方法進行了改進,但仍然存在一些缺點。首先,無法測定微小昆蟲(如,西花薊馬,煙粉虱等)的過冷卻點。因此,在今后的研究中需對探頭進行改進,提高其靈敏度,方可滿足一些小型昆蟲過冷卻點的測定。其次,要盡量聯(lián)合其他指標評價昆蟲耐寒性的強弱。
1.2 半致死溫度或半致死時間
由于許多昆蟲屬于避凍型,常常在高于過冷卻點的溫度死亡,因而過冷卻點不能作為衡量耐寒性的指標,至少不能完全代表昆蟲的耐寒性。因此,昆蟲的耐寒性主要取決于在低溫下的存活率。在昆蟲毒理學中常采用LC50表示殺蟲劑或化學物質(zhì)對昆蟲的毒力。類似,目前主要采用半致死溫度(Ltem50)或半致死時間(Ltime50)評價昆蟲耐寒性。Ltem50主要指在特定時間下,導致昆蟲50%個體死亡所對應(yīng)的溫度;而Ltime50是指在特定低溫條件下,導致昆蟲50%個體死亡所對應(yīng)的時間。通常是將供測試的昆蟲置于5~7個低溫下,所設(shè)置的溫度范圍和溫度間隔因昆蟲的種類而不同。所設(shè)置的溫度不能太高也不能太低。溫度過高昆蟲全部存活,而溫度過低又導致個體全部死亡,無法估算出致死中溫。總的原則是最高溫度下的死亡率為0%~10%,而最低溫度下的死亡率為90%~100%,保證中間溫度的死亡率為50%~60%。同樣,將供試昆蟲暴露在特定低溫下不同的時間時,保證最短時間和最長時間所對應(yīng)的死亡率分別在0%~10%和90%~100%就能計算出Ltime50。
通常情況下,要想得到理想的Ltem50或Ltime50需要進行預(yù)備試驗。Ltem50或Ltime50可以利用邏輯斯蒂曲線進行擬合求得,其擬合曲線一般呈S型[13]。公式為y=k/(1+ea+bx)。式中,y為存活率,x為溫度,k、a、b為相關(guān)估計系數(shù)。人們利用該模型對飛蝗[14]、斑潛蠅[15]、紅火蟻[16]、方翅網(wǎng)蝽[11]和蘋淺褐卷蛾[17]的Ltem50進行估算。Jing等[14]研究發(fā)現(xiàn),飛蝗在不同季節(jié)的抗寒性具有顯著差異,夏季、秋季和冬后在低溫下暴露6 h的Ltem50依次為-24.4℃、-28.6℃和-11.6℃,說明,飛蝗抗寒性高低依次為秋季>夏季>冬后[11]。在實際研究中,Ltem50或Ltime50與氣候數(shù)據(jù)結(jié)合,可以大致判斷某種害蟲的分布范圍和在某一地區(qū)的越冬情況;同時在室內(nèi)也常作為利用低溫殺滅倉儲害蟲的依據(jù)。因此,Ltem50或Ltime50已成為評價抗寒性最常用的指標。
1.3 傷害低溫總和
在低溫條件下暴露時間也是影響昆蟲存活率不可忽略的因素。在評價昆蟲的耐寒性時,暴露溫度常常是作為強度因子,而暴露時間則是數(shù)量因子。昆蟲耐寒性同時受到暴露溫度和暴露時間的交互影響。為了將2個因子同時用于評價昆蟲的耐寒性,Nedve提出“傷害低溫總和(Sum of injurious temperature, SIT) ”的概念,其是指導致有效冷傷害的時間和有效低溫的共同作用,即所獲低溫脅迫的累積劑量。利用三維結(jié)構(gòu)圖(圖2A)能很好地反映存活率、暴露低溫和暴露時間三者之間的關(guān)系[18]。
從圖2B可知,溫度和偽蘋果蠹的存活率呈正相關(guān),即溫度越低導致存活率越低;而偽蘋果蠹的存活率與暴露時間呈負相關(guān),即在一定低溫下暴露時間的增加導致該害蟲存活率降低。此三者之間的關(guān)系采用數(shù)學模型可表示為
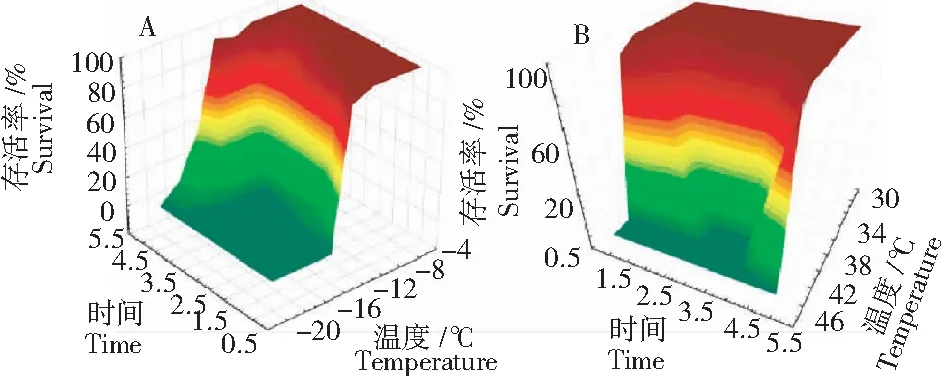
注:A,低溫; B,高溫。
Note:A, low temperature; B, high temperature.
圖2 不同溫度和時間下偽蘋果蠹蛾的存活率
Fig.2 Mean survival ofT.haumatotibialeucotretaat different test temperatures for different experimental durations
式中,t表示時間,T表示溫度;S(t,T)為昆蟲種群在時間t和溫度T條件下的死亡率(%),a, b, c為常數(shù)。利用該模型可計算昆蟲在恒定低溫下的存活率,但昆蟲在自然界所經(jīng)歷的環(huán)境溫度是多變的,因此,該模型存在2個缺點: 1) 不能用于估算昆蟲在變溫下的存活率; 2) 未考慮影響昆蟲抗寒性的其他因素。
為此,需要建立一個更加全面的模型評價昆蟲的耐寒性[3]。鐘景輝[19]對上述公式進行改進,得公式:
S(t,T)=e(a+b∑tiTi)/[ 1+(a+b∑tiTi)]
式中,S為死亡率,可通過試驗測定;ti為特定低溫暴露時間,其單位可根據(jù)試驗具體情況選擇天、小時或分鐘等時間單元;Ti為特定低溫暴露溫度,單位為℃;i為變化溫度處理數(shù),i=1,2,3,…;∑tiTi為變化低溫處理的低溫累積,可以通過各溫度及其暴露的時間計算;a、b為常數(shù),其中-a/b的生物學意義為造成昆蟲50%個體死亡時的低溫累積,即“半致死低溫累積”(Semi-lethal sum of chill injurious temperature, LSCIT50),以此指標比較耐寒性的強弱。通過上述公式計算松突圓蚧成蟲春夏秋冬四季的LSCIT50分別為-181.62、-56.54、-175.87和-219.38,說明該害蟲的耐寒性具有季節(jié)性差異,其抗寒性強弱依次為冬季>春季>秋季>夏季。
與前人的方法相比,改進后的方程既充分考慮了低溫暴露強度和低溫暴露時間2個因素的交互影響,又克服了前人研究不能評價變化低溫暴露試驗中昆蟲耐寒性的缺點,但該模型沒有考慮到對昆蟲造成冷傷害的低溫累積上限(Upper limited sum of chill injurious temperature, ULSCIT)。因此,該方法只適用于不同研究對象耐寒性的比較或在已知試驗設(shè)置的低溫累積必然造成冷傷害的條件下昆蟲耐寒性的評價[19]。
1.4 冷傷害昏迷恢復時間
冷傷害昏迷恢復時間(Chill coma recovery time)作為衡量昆蟲耐寒性的指標[1,20-22]被廣泛接受。其定義是經(jīng)低溫處理的昆蟲在較高的溫度環(huán)境中從冷傷害昏迷中蘇醒到正常活動狀態(tài)所需的時間。Marais E等[22]收集Drosophilamelanogaster和D.ananassae2種果蠅成蟲于玻璃瓶中,置于-5℃的水浴中30~45 min后,將成蟲和若蟲倒入培養(yǎng)皿中,迅速放入20℃的恒溫培養(yǎng)箱中讓其恢復。通過培養(yǎng)箱的玻璃門記錄果蠅從水浴中取出到開始能正常爬行的時間,即為冷傷害昏迷恢復時間。該研究發(fā)現(xiàn),D.melanogaster在20℃下的恢復時間為8 min,而D.ananassae的恢復時間約為25 min,說明,D.melanogaster的抗寒性強于D.ananassae。該方法的優(yōu)點是操作比較簡單,但缺點是比較費時。
2 昆蟲耐熱性的評價方法
2.1 Ltem50和Ltime50
昆蟲耐熱性的Ltem50和Ltime50測定方法同抗寒性。
2.2 半致死高溫累積
半致死高溫累積(Semi-lethal sum of heat injurious temperature, LSHIT50)的評價方法同LSCIT50。由于該模型沒有考慮到對昆蟲造成熱傷害的高溫累積下限(Lower limited sum of heat injurious temperature, LLSHIT),因此只適用于不同研究對象耐熱性的比較或在已知試驗設(shè)置的高溫累積必然造成熱傷害的條件下昆蟲耐熱性的評價。同樣,昆蟲的耐熱性受到暴露溫度和暴露時間交互作用的影響。存活率、暴露高溫和暴露時間三者之間的關(guān)系可以利用三維結(jié)構(gòu)圖(圖2B)表示[18]。在高溫下,昆蟲的存活率與暴露溫度和暴露時間都呈負相關(guān),即暴露溫度越高,暴露時間越長,昆蟲的存活率越低。
2.3 臨界最高溫度
臨界最高溫度(Critical temperature maximum, CTmax)被用于定義脊椎和無脊椎動物的耐熱性已有60年的歷史。臨界最高溫度的早期定義是指運動器官受到損傷,昆蟲喪失逃避能力時的溫度[23]。Lutterschmidt等[24]認為,CTmax是一個很好的用于評價生物高溫生理和散熱能力的標準。
傳統(tǒng)上,用于定義CTmax的生理狀態(tài)是不斷變化的,包括失去翻正反應(yīng)(loss of righting response),肌肉痙攣的開始(onset of muscular spasms),熱癱瘓(heat paralysis),熱昏迷(heat coma),甚至熱擊倒(knockdown)[25]。Cowles R B等[23]將CTmax定義為運動器官和體壁透氣功能的正常運作上限溫度用于果蠅耐熱性研究。前人的研究表明,CTmax主要是通過測量上述涉及的生理狀態(tài)發(fā)生時昆蟲體壁的溫度進行估算。隨著研究的發(fā)展,研究者常采用更加客觀的方法,即用呼吸運動熱量極限法和檢測活動能力法相結(jié)合的方式估計CTmax[26-27]。呼吸運動熱量極限法適用于測量具有非隨意肌物種的CTmax,以體壁通氣功能的喪失表示;而活動能力監(jiān)測法則適宜具有隨意肌物種CTmax的檢測,以運動能力的喪失表示。該方法的優(yōu)點是既適于對非隨意肌也適于對隨意肌物種CTmax的測定。體壁通氣功能的喪失主要通過二氧化碳輸出模式的改變判斷。以果蠅成蟲為例,高溫馴化品系的CTmax為2.4 μL/(h·mg),而對照品系的CTmax為1.9 μL/(h·mg),說明,經(jīng)高溫馴化后,果蠅的耐寒性得到加強[27]。總體而言,該方法的優(yōu)點是能真實地反映昆蟲的耐熱性,缺點是儀器設(shè)備的價格十分昂貴。
2.4 擊倒溫度和擊倒時間
熱擊倒溫度(Knock down temperature)是指生物在加熱過程中倒下那一刻所對應(yīng)的溫度,而熱擊倒時間(Knock down time)是指生物從暴露在高溫下到擊倒所需的時間。一般的操作方法是將昆蟲裝在透明的容器內(nèi),置于加熱容器中,從室溫開始以一定的速率(如0.01℃/min)開始加熱。在加熱的初始階段,昆蟲會異常的興奮。隨溫度的繼續(xù)升高,昆蟲的活動能力逐漸減弱。達昆蟲瞬時倒下那一刻的溫度就稱之為擊倒溫度。而擊倒時間的測定方法是,從室溫開始以一定速率開始加熱,當達某一溫度后,在此溫度下持續(xù)保持一段時間,直到昆蟲倒下那一刻,記錄所經(jīng)歷的時間,即為熱擊倒時間。目前,許多學者運用該指標評價模式昆蟲果蠅和一些農(nóng)業(yè)害蟲的耐熱性[28-30]。余昊[31]研究發(fā)現(xiàn),溫室白粉虱在45℃下的擊倒時間約為30 min,而B型煙粉虱的擊倒時間則接近45 min,說明B型煙粉虱的耐熱性要顯著強于溫室白粉虱。
3 結(jié)論與討論
全球氣候變化已是一個不爭的事實,而昆蟲如何適應(yīng)氣候變化則是未來生態(tài)學領(lǐng)域研究的一個熱門話題。因此,在評價昆蟲的溫度耐受性時,方法的選擇尤為重要。斜坡式的變溫被認為更接近昆蟲在自然條件下所經(jīng)歷的溫度變化,而急劇的溫度變化是昆蟲在自然界不常遇到的。當然,斜坡式的評價方式也存在對昆蟲的耐受性估計不足的問題。如,昆蟲在緩慢的溫度變化過程中遭遇饑餓和干燥從而影響最終結(jié)果的準確性;同時,由于該混淆效應(yīng)影響昆蟲溫度耐受性的變化和遺傳特性。然而,人們認為斜坡式的評價方式更能客觀地反映生態(tài)學事實[4],盡管其他脅迫因子的混淆效應(yīng)會影響昆蟲在自然界的溫度耐受性極限。此外,在斜坡式的評價方法中,昆蟲所遭遇的干燥和饑餓脅迫所帶來的影響微小,除非評價過程中涉及的是小動物和持續(xù)的時間較長。因此,在進行比較研究和理解潛在的混淆效應(yīng)時,方法的選擇十分重要。基于斜坡式的溫度耐受性評價仍然是理解和預(yù)測昆蟲對氣候變化響應(yīng)的重要方法。
[1] Hoffmann A A,S?rensen J G,Loeschcke V.Adaptation of Drosophila to temperature extremes: bringing together quantitative and molecular approaches[J].Journal of Thermal Biology,2003,28:175-216.
[2] 劉麗輝,張 帆,吳珍泉.溫度對西花薊馬(Frankliniellaoccidentalis)生長發(fā)育和存活率的影響[J].生態(tài)學報,2008,28(10):4891-4895.
[3] 景曉紅,康 樂.昆蟲耐寒性的測定與評價方法[J].昆蟲知識,2004,40(1):7-10.
[4] Terblanche J S,Hoffmann A A,Mitchell K A,et al. Ecologically relevant measures of tolerance to potentially lethal temperatures[J].Journal of Experimental Biology,2011,214:3713-3725.
[5] Nguyen C,Bahar M H,Baker G,et al.Thermal tolerance limits of diamondback moth in ramping and plunging assays[J].PLoS ONE,2014,9(1):e87535.
[6] Bahar M H,Hegedus D,Soroka J,et al.Survival and Hsp70 gene expression inPlutellaxylostellaand its larval parasitoid Diadegma insulare varied between slowly ramping and abrupt extreme temperature regimes[J].PLoS ONE,2013,8(9):e73901.
[7] 烏莎廷斯卡姬.昆蟲耐寒性原理[M].張淑德,巫國瑞,譯.北京:科學出版社,1960.
[8] 鞠瑞亭,杜予州.昆蟲過冷卻點的測定及抗寒機制研究概述[J].武夷科學,2002(18):252-257.
[9] 秦玉川,楊建才.一種便攜式測定昆蟲過冷卻點的方法[J].昆蟲知識,2000,47(4):236-238.
[10] 呂龍石,叢明亮,孟艷玲.一種昆蟲過冷卻點的簡易測定裝置[J].昆蟲知識,2010,47(6):1257-1260.
[11] Ju R T,Wang F,Xiao Y Y,et al.Supercooling capacity and cold hardiness of the adults of the Sycamore Lace Bug,Corythuchaciliate(Hemiptera: Tingidae)[J].CryoLetters,2010,31(6):445-453.
[12] Zhao Y X,Kang L.Cold tolerance of the leafminerLiriomyzasativae(Diptera:Agromyzidae)[J].Journal of Applied Entomology,2000,124:185-189.
[13] 徐 康,夏宜平,徐碧玉,等.以電導法配合Logistic方程確定茶梅‘小玫瑰’的抗寒性[J].園藝學,2005,32(1):148-150.
[14] Jing X H,Kang L.Seasonal changes in the cold tolerance of eggs of the migratory locust,LocustamigratoriaL.(Orthoptera:Acrididae).Environmental Entomology,2004,33(2):113-118.
[15] Chen B,Kang L.Cold hardiness and supercooling capacity in the pea leafminerLiriomyzahuidobrensis. Cryoletters,2002,23(3):173-182.
[16] 許益鐫,陸永躍,黃 俊,等.紅火蟻自然種群耐寒性的研究[J].昆蟲學報,2009,52(9):974-983.
[17] Bürgi L P,Mills N J.Cold tolerance of the overwintering larval instars of light brown apple mothEpiphyaspostvittana[J].Journal of Insect Physiology,2010,56:1645-1650.
[18] Nedved O,Lavy D,Verhoef H A. Modelling the time-temperature relationship in cold injury and effect of high-temperature interruptions on survival in a chill-sensitive collembolan. Functional Ecology,1998,12(5):816-824
[19] Boardman L,S rensen J G,Terblanche J S.Physiological responses to fluctuating thermal and hydration regimes in the chill susceptible insect,Thaumatotibialeucotreta[J].Journal of Insect Physiology,2013,59(8):781-94.
[20] 鐘景輝.松突圓蚧及其天敵花角蚜小蜂對極端溫度的耐受性[D].福州:福建農(nóng)林大學,2009.
[21] Gibert P,Moreteau B,Petavy G,et al.Chill-coma tolerance, a major climatic adaptation amongDrosophilaspecies[J].Evolution,2001,55:1063-1068.
[22] Marais E,Chown S L.Beneficial acclimation and the bogert effect[J].Ecological Letters,2008,11:1027-1036.
[23] Cowles R B,Bogert C M.A preliminary study of the thermal requirements of desert reptiles[J].Bulletin of the American Museum of Natural History,1994,83:261-296.
[24] Lutterschmidt W I,Hutchison V H.The critical thermal maximum: history and critique[J].Canadian Journal of Zoology,1997,75:1561-1574.
[25] Berrigan D,Hoffmann A A.Correlations between measures of heat resistance and acclimation in two species ofDrosophilaand their hybrids[J].Biological Journal of the Linnean Society,1998,64:449-462.
[26] Lighton J R B,Turner R J.Thermolimit respirometry: an objective assessment of critical thermal maxima in two sympatric desert harvester ants,Pogonomyrmexrugosus andP.californicus[J].Journal of Experimental Biology,2004,207:1903-1913.
[27] Folk D G,Hoekstra L A,Gilchrist G W.Critical thermal maxima in knock down selectedDrosophila:are thermal endpoints correlated?[J].Journal of Experimental Biology,2007,210:2649-2656.
[28] Hoffmann A A.Physiological climatic limits inDrosophila:patterns and implications[J]. Journal of Experimental Biology,2010,213:870-880.
[29] Mitchell K,Hoffmann A A.Thermal ramping rate influences evolutionary potential and species differences for upper thermal limits inDrosophila[J].Functional Ecology,2010,24:694-700.
[30] Overgaard J,HoffmannA A,Kristensen T N.Assessing population and environmental effects on thermal resistance inDrosophilamelanogasterusing ecologically relevant assays[J].Journal of Thermal Biology,2011,36:409-416.
[31] 余 昊.入侵粉虱熱激蛋白基因表達與功能[D].北京:中國農(nóng)業(yè)科學院,2009.
(責任編輯: 王 海)
Advances in Evaluation Measures of Thermal Tolerance in Insect
LI Hongbo, HE Yongfu*
(GuizhouInstituteofPlantProtection,Guiyang,Guizhou550006,China)
Insects are ectotherms, and have strong dependence on environmental temperature. Adaptation to temperature stress directly determines population dynamics, life cycles and geographic distribution of insects. To better evaluate thermal tolerance of insect, the authors summarized and evaluated the measures of temperature tolerance.
temperature tolerance; evaluation measures; insect
2015-03-25; 2015-12-28修回
貴州省農(nóng)業(yè)科學院人才引進啟動項目“sHsps在粘蟲兩型中的表達模式研究”(201301)
李鴻波(1985-),男,副研究員,博士,從事昆蟲生理生化與害蟲綜合治理研究。E-mail:lihongb2007@163.cm
*通訊作者:何永福(1965-),男,研究員,從事作物病蟲草害綠色防控技術(shù)應(yīng)用研究。E-mail:heyongfu08@163.com
1001-3601(2016)01-0022-0080-04
S431
A
猜你喜歡
石油瀝青(2021年4期)2021-10-14 08:50:44
世界科學技術(shù)-中醫(yī)藥現(xiàn)代化(2021年10期)2021-03-02 05:52:06
兒童故事畫報(2019年5期)2019-05-26 14:26:14
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
小雪花·成長指南(2015年7期)2015-08-11 15:03:12
小雪花·成長指南(2015年4期)2015-05-19 14:47:56
中國教育技術(shù)裝備(2015年19期)2015-03-01 02:43:07
中國工程咨詢(2015年2期)2015-02-14 02:59:26
俄羅斯問題研究(2012年1期)2012-03-25 09:54:51
