性狀的遺傳力對標記輔助導入效率的影響
2016-04-11 20:04:51白俊艷楊又兵黃勇祁艷霞李
江蘇農業科學 2016年2期
白俊艷++楊又兵++黃勇++祁艷霞++李廣錄


摘要:分析性狀的遺傳力對標記輔助導入效率的影響,期望為實踐中標記輔助導入提供參考。對于標記輔助導入1個QTL過程,采用兩側標記選擇進行前景選擇,MBLUP為背景選擇,對于前景性狀和2個背景性狀給出3種遺傳力水平,即0.20、0.10、0.15(低);0.50、0.30、0.40(中);0.80、0.50、0.70(高)。結果表明,導入QTL頻率和前景性狀遺傳進展在遺傳力為低、中、高之間差異很小,在橫交階段,中遺傳力和高遺傳力組的導入QTL頻率和前景性狀遺傳進展略高于低遺傳力組。
關鍵詞:標記輔助導入;遺傳力;MBLUP;前景性狀;背景性狀
中圖分類號: Q756文獻標志碼: A
文章編號:1002-1302(2016)02-0035-03
收稿日期:2015-02-11
基金項目:國家自然科學基金(編號:31201777)。
作者簡介:白俊艷(1975—),女,蒙古族,內蒙古赤峰人,博士,副教授,主要從事動物分子數量遺傳學方面的研究。E-mail:junyanbai@163.com。
在高密度的遺傳圖譜和QTL檢測及其定位基礎上,可以進行標記輔助導入(marker assisted introgression,MAI),即通過回交將1個品種(供體)的某個或某些優良基因利用標記信息輔助導入到另1個品種(受體),同時盡可能保持受體的遺傳背景不發生改變,使得該品種既獲得了供體的優良基因,又保持了原有的所有優良基因。Hanset等將氟烷陰性等位基因利用標記信息導入到皮特蘭豬中,采用與RYR座位緊密連鎖的標記進行前景選擇,將抗應激敏感的基因成功地導入到皮特蘭豬中,經過3次回交獲得氟烷陰性的皮特蘭品系[1]。Yancovich等將1個商業蛋雞系中的裸頸基因導入到1個商業肉雞系中,利用基因組進行背景選擇,以加快肉雞系的生長速度和體質量恢復[2]。Gootwine等借助與FecB座位緊密連鎖的2個微衛星標記OarAE101和BM1329對FecB進行前景選擇,將Booroola羊中的FecB基因利用標記信息輔助導入到Awassi乳用綿羊中[3]。由于標記輔助導入中的標記基因型分析費用太貴且育種過程復雜,因此大多數研究者運用計算機模擬標記輔助導入的育種過程,進而摸索和探討標記輔助導入的影響因素及規律[4-20]。本研究分析了性狀的不同遺傳力對標記輔助導入效率的影響,以期為實踐中的標記輔助導入提供參考。
1材料與方法
1.1試驗設計
標記輔助導入一個QTL的試驗設計,假設基礎群的供體群和受體群所有個體之間無親緣關系,在所有世代中要避免同胞交配,后代性別比例按1 ∶[KG-*3]1的概率確定,群體規模為 1 000,所有世代間不重疊。假設整個基因組分布在10條染色體上,所有標記均勻地分布在每條染色體上,每個標記座位有2個等位基因,假設需要導入的QTL記作QTL,背景QTL記作QTL1和QTL2。對于前景性狀和2個背景性狀這3個性狀,本研究考慮以下3種遺傳力水平,即0.20、0.10、0.15(低);0.50、0.30、0.40(中);0.80、0.50、0.70(高)(圖1)。
[FK(W22][TPBJY1.tif][FK)]
1.2性狀的表型值
(1)基礎群性狀的表型值y(yi=qi+ui+ei),其中yi為個體i的表型值,qi為個體i的QTL基因型值,ui為個體i的多基因效應值,對于2個親本群體,ui由正態分布N(0,σu2)隨機產生,ei為個體i的隨機環境離差,在所有世代中,ei均由正態分布N(0,σe2)隨機產生。(2)非基礎群的個體性狀表型值的產生方法與基礎群相同,不同的是多基因效應值的計算(ui=0.5us+0.5ud+mi),其中ui為個體i的多基因效應值,us和ud分別為個體i的父親和母親的多基因值,mi為個體i的孟德爾抽樣離差(Mendelian sampling),服從于正態分布N{0,(σu2/2)[1-(Fs+Fd)/2]},其中Fs和Fd分別為父親和母親的近交系數。
1.3選擇方法
1.3.1前景選擇假設導入QTL本身未知,同時利用與導入QTL相鄰最近的兩側標記進行間接選擇,選擇這2個標記均為雜合型的個體,稱為雙側標記選擇。
1.3.2背景選擇標記輔助BLUP選擇(MBLUP),首先利用以下MBLUP對2個背景性狀進行育種值估計,然后將育種值合并為指數,根據指數的高低來選種。根據Femando 和 Grossman提出的混合模型,假設隨機QTL效應和隨機多基因效應為性狀的遺傳基礎,則個體表型值可以用以下線性模型來描述,以矩陣形式表示為y=Zu+Wv+e,其中y為性狀觀察值向量,u為隨機剩余多基因效應值向量,其均值為0,方差協方矩陣為Aσu2,A為分子親緣相關矩陣,v為隨機QTL等位基因效應向量,其均值為0,方差協方矩陣為Gσv2,G為QTL配子相關矩陣,e為殘差向量,其均值為0,方差協方矩陣為Iσe2,I為單位向量,W和Z分別為v和u的結構矩陣。
2結果與分析
2.1導入QTL頻率和前景性狀遺傳進展
導入QTL頻率在不同性狀遺傳力之間差異很小。在回交階段,導入QTL頻率在不同性狀遺傳力之間差別很小,經過回交5個世代,前景性狀遺傳力為低、中和高的導入QTL頻率分別為0.233 1、0.244 8、0.244 0。到了橫交階段,經過橫交2個世代,前景性狀遺傳力為低、中和高下的導入QTL頻率分別為0.918 2、0.962 8、0.960 6。經過橫交5個世代,前景性狀遺傳力為低、中和高的導入QTL頻率分別增加到 0.944 6、0.974 8、0.972 7。可見,性狀遺傳力低的導入QTL頻率為較小,其他性狀遺傳力的導入QTL頻率基本一致,并且都高于前者。前景性狀遺傳進展在不同性狀遺傳力之間差異很小。經過橫交2個世代,這3種性狀遺傳力的前景性狀的平均育種值分別增加到3.727 9、4.013 3、4.018 6。由于前景選擇都利用兩側標記,因此前景性狀遺傳進展在前景性狀遺傳力為低、中、高之間差異不大(圖1)。
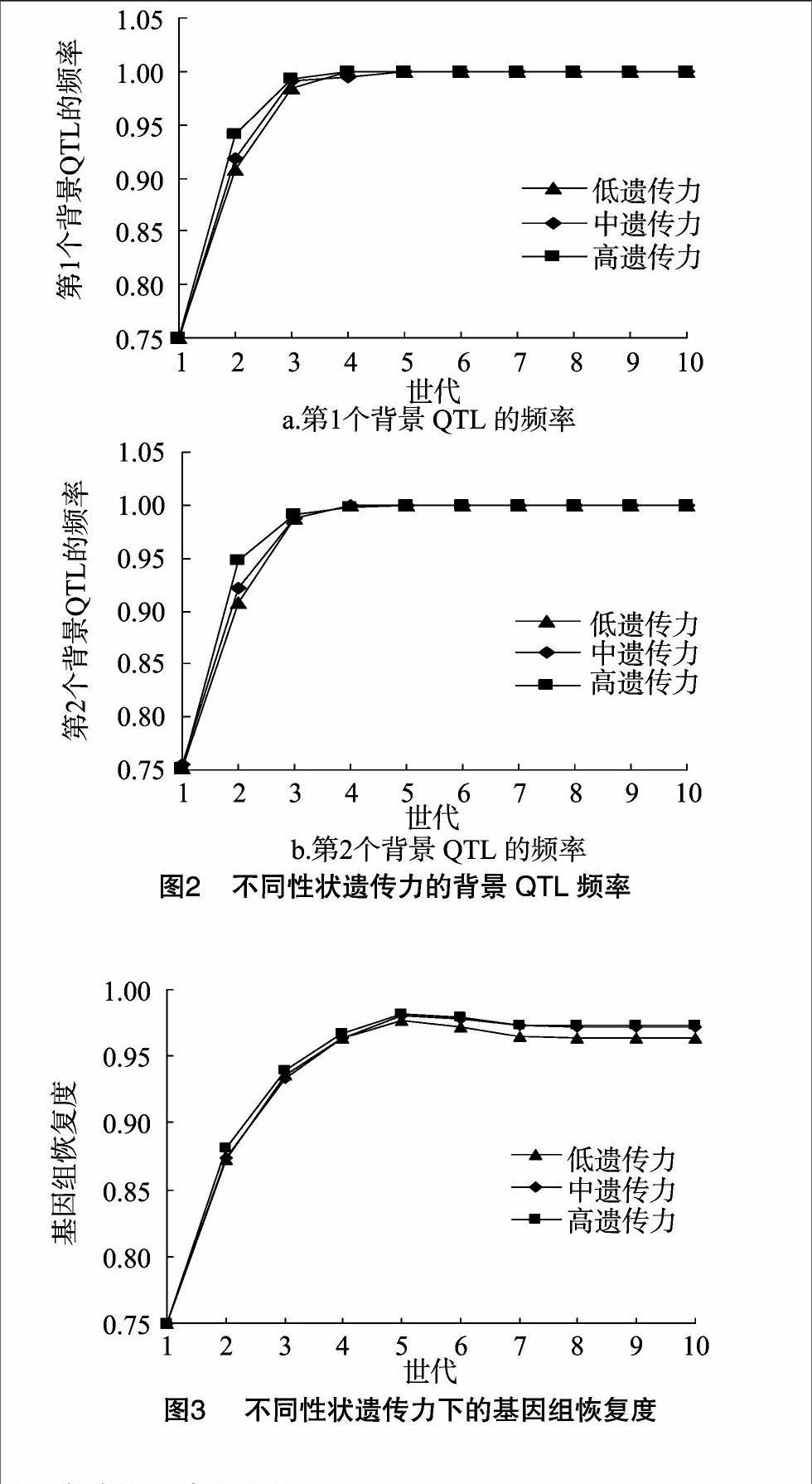
2.2背景QTL頻率
背景QTL頻率在不同性狀遺傳力之間略有差異。在回交階段,背景QTL頻率隨著背景性狀遺傳力為低、中和高而呈現遞增趨勢,尤其是經過回交2個世代,背景性狀遺傳力為低、中和高的第2個背景QTL頻率分別為0.907 2、0.922 0、0.947 1,并且經過回交4個世代,第2個背景QTL頻率都已經達到1,而到了橫交階段,背景QTL頻率在不同性狀遺傳力之間沒有差異,它們的頻率均為1。第1個背景QTL頻率在不同性狀遺傳力之間的變化趨勢與第2個背景QTL頻率的變化趨勢基本一致(圖2)。
2.3遺傳背景的恢復
受體遺傳背景的恢復在不同的性狀遺傳力之間略有差異。在回交階段,遺傳背景的恢復隨著回交世代的增加而呈現出迅速遞增趨勢,經過回交5個世代,背景性狀遺傳力為低、中、高的遺傳背景的恢復度分別為97.67%、98%、98.19%。經過橫交5個世代,背景性狀遺傳力為低、中和高下的遺傳背景的恢復度分別為96.31%、97.23%、97.34%。可見背景性狀為中、高遺傳力時,兩者遺傳背景的恢復度基本相同,都略高于低遺傳力(圖3)。
2.4背景性狀遺傳進展
背景性狀遺傳進展在不同性狀遺傳力之間的差異很大,2個背景性狀遺傳進展均呈現出隨背景性狀遺傳力為低、中、高的增加而呈現出遞增趨勢。在回交階段,經過回交5個世代,背景性狀遺傳力為低、中、高下的第2個背景性狀的平均育種值分別為3.721 7、4.625 2、5.354 2。到了橫交階段,經過橫交5個世代,這3種性狀遺傳力的第2個背景性狀的平均育種值分別增加到6.234 5、9.549 2、13.181 3。雖然利用MBLUP 對低遺傳力性狀進行選擇獲得的遺傳進展小一些,但是相對常規選擇方法的效率卻會更高(Ruane and Colleau,1996),因此,對于低遺傳力的背景性狀,MBLUP仍是比較理想的背景選擇方法(圖4)。
3結論與討論
[CM(24]由于分子標記不易受性別、年齡等影響,能夠在早期獲[CM)]
[FK(W21][TPBJY4.tif][FK)]
得,因此利用標記進行前景選擇是一個比較好的策略。Visscher等模擬研究了在回交育種中的標記輔助導入,發現經過回交3個世代,兩側標記選擇的個體攜帶了目標等位基因1個拷貝的比例為49%[21]。本研究結果與前人的研究結果基本一致。兩側標記選擇的正確率要比單側標記高,由于兩側標記發生雙交換的概率低,因此雙交換配子的比例很小,絕大部分為親本型配子,所以利用兩側標記對目標基因進行跟蹤選擇,可以大大提高選擇的正確率,不易使目標基因丟失。此外,Hospital等研究指出目標控制率與座位之間的重組率和回交世代數有直接關系,并發現隨回交世代數的增加其目標控制率呈下降趨勢,有兩側標記比單側標記的目標控制率高[22],由此推導出導入QTL的條件概率公式如下,當QTL的真實位置是x已知時,它位于xk和xk+12個標記之間,回交t世代的后代在所有標記上含有導入QTL的條件概率可表示為pQ|W[t]=f(x)=[SX(][1-r(xk,x)]t[1-r(x,xk+1)]t[1-r(xk,xk+1)]t[SX)]。而在實踐中,QTL的位置往往是未知的,當QTL的真實位置未知時,則需要估計QTL的位置,此時導入QTL的條件概率必須綜合在所有QTL的假定位置上才可以計算,此時含有導入QTL的條件概率可表示為pQ|W[t]=∫L0f[x]g[x]dx,其中g[x]是QTL期望位置的概率密度,L是染色體總長度。
隨著基因組研究的飛速發展,家畜中具有重要經濟意義的QTL或主基因被人們相繼發現和定位,因此,在標記輔助導入過程中對目標基因的識別將不成問題。近年來,許多研究者在如何恢復遺傳背景方面做了大量研究,以期望能找到恢復遺傳背景的最有效途徑。結合前人的研究成果,本研究將MBLUP方法引入到標記輔助導入的背景選擇中。利用MBLUP雖然不能使遺傳背景迅速全部恢復,但是能使受體的特定背景性狀迅速恢復,同時使這些背景性狀在導入過程中獲得最大的遺傳進展。在實際育種中,人們關心的主要是那些有重要經濟價值的性狀,在進行標記輔助導入時,并不一定嚴格要求受體的遺傳背景完全恢復,但要求某些特定的背景性狀一定要得到恢復,這時利用與影響這些性狀的QTL連鎖的標記(而不是覆蓋整個基因組的全部標記)和性狀的表型信息進行標記輔助BLUP(MBLUP)背景選擇就可以達到目的,而這種選擇的成本要比利用整個基因組的全部標記進行背景選擇低得多,因而利用MBLUP進行背景選擇可能是一個更實際的選擇方案。
人們對性狀遺傳力的大小對標記輔助導入影響的研究很少,僅見Visscher 等在模擬研究中考慮了性狀遺傳力為0.1和0.4對標記輔助導入的影響,發現經過回交3個世代,利用表型進行背景選擇,0.1和0.4性狀遺傳力的平均遺傳值分別為76.9和85.1,而利用標記進行背景選擇的平均遺傳值分別為94.8和94.7,可見高性狀遺傳力在表型選擇里能獲得更高的育種值,而在標記選擇里性狀遺傳力的大小對育種值基本無影響。本研究考慮了3種不同性狀遺傳力對標記輔助導入的影響,研究結果與前人的研究結果部分一致,由于前景選擇是利用標記進行選擇的,因此,導入QTL頻率和前景性狀遺傳進展在不同性狀遺傳力之間基本無差異,而背景選擇是利用MBLUP選擇,因此,背景QTL頻率和背景性狀遺傳進展卻隨性狀遺傳力的增大而增大。
[HS2][HT8.5H]參考文獻:[HT8.SS]
[1][ZK(#]Hanset R,Dasnoi C,Scalais S,et al. Effets de lintrogression dons le genome Piétrain de lallele normal aux locus de sensibilité a lhalothane[J]. Genet Select Evol,1995,27(1):77-88.
[2]Yancovich A,Levin I,Cahaner A,et al. Introgression of the avian naked neck gene assisted by DNA fingerprints[J]. Anim Genet,1996,27(3):149-155.
[3]Gootwine E,Yossefi S,Zenou A,et al. Marker assisted selection for FecB carriers in Booroola Awassi crosses[C]. Proc 6th World Cong Genet Appl Livest Prod,1998.
[4]Young N D,Tanksley S D. Restriction fragment length polymorphism maps and the concept of graphical genotypes[J]. Theor Appl Genet,1989,77(1):95-101.
[5]Young N D,Tanksley S D. RFLP analysis of the size of chromosomal segments retained around the Tm-2 locus of tomato during backcross breeding[J]. Theor Appl Genet,1989,77(3):353-359.
[6]Hillel J,Schaap T,Haberfeld A,et al. DNA fingerprints applied to gene introgression in breeding programs [J]. Genetics,1990,124(3):783-789.
[7]Hospital F,Chevalet C,Mulsant P. Using markers in gene introgression breeding programs [J]. Genetics,1992,132(4):1199-1210.
[8]Hospital F. Marker-assisted breeding[M]//Newbury H J. Plant molecular breeding. London:Blackwell Scientific Publishers,2003:30-56.
[9]Hospital F. Size of donor chromosome segments around introgressed loci and reduction of linkage drag in marker-assisted backcross programs[J]. Genetics,2001,158(3):1363-1379.
[10][ZK(#]Groen A E,Timmermans M M J. The use of genetic marker increase the efficiency of introgression a simulation study[C]. Proceedings of the Ⅹ[KG-*3]Ⅸ Worlds Poultry Congress,1992:523-527.
[LM]
[11][ZK(#]Groen A F,Smith C. A stochastic simulation study on the efficiency of marker-assisted introgression in livestock[J]. Joumal of Animal Breeding and Genetics,1995,112:161-170.
[12]Visscher P M,Haley C S. On the efficiency of marker-assisted introgression[J]. Animal Science,1999,68(1):59-68.
[13]Visscher P M. Proportion of the variation in genetic composition in backcrossing programs explained by genetic markers[J]. Jourmal of Heredity,1996,87(2):136-138.
[14]van Heelsum A M,Haley C S,Visscher P M. Marker-assisted introgression using non-unique marker alleles:Ⅱ. Selection on probability of presence of the introgressed allele[J]. Animal Genetics,1997,28(3):188-194.
[15][JP3]van Heelsum A M,Visscher P M,Haley C S. Marker-assisted introgression using non-unique marker alleles:Ⅰ. Selection on the presence of linked marker alleles[J]. Animal Genetics,1997,28,181-187.
[16]Frisch M,Bohn M,Melchinger A E. Minimum sample size and optimal positioning of flanking markers in marker-assisted backcrossing for transfer of a target gene[J]. Crop Science,2001,41(6):1716-1725.
[17]Frisch M,Melchinger A E. Marker-assisted backcrossing for simultaneous introgression of two genes[J]. Crop Science,2001,41(6):1716-1725.
[18]Frisch M,Melchinger A E. The length of the intact donor chromosome segment around a target gene in marker-assisted backcrossing[J]. Genetics,2001,157(3):1343-1356.
[19]Chaiwong N,Dekkers J C M,Fernando R L,et al. Introgressing gressing multiple QTL in backcross breeding programs of limited size[C]. Proc 7th World Cong Genet Appl Livest Prod,2002.
[20]Bai J Y,Zhang Q,Jia X P.Comparison of different foreground and background selection methods in marker-assisted introgression[J]. Acta Genetica Sinica,2007,33 (12):1073-1080.
[21]Visscher P M,Haley C S,Thompson R. Marker-assisted introgression in backcross breeding programs[J]. Genetics,1996,144(4):1923-1932.
[22]Hospital F,Charcosset A. Marker-assisted introgression of quantitative trait loci[J]. Genetics,1997,147(3):1469-1485.
