玉米穗行數基因的QTL定位與分析
2016-04-11 18:05:14翟立紅周蘭庭韓鵬騰峰
江蘇農業科學 2016年2期
翟立紅++周蘭庭+++韓鵬++騰峰


摘要:對玉米中控制穗行數的QTL進行定位和分析,為分子標記輔助選擇育種提供理論基礎。在一套以綜3為遺傳背景攜帶衡白522置換片段的染色體片段置換系群體進行QTL初步定位的基礎上,以穗行數明顯減少的置換系SIL8為材料,構建置換片段內的F2次級分離群體和跨疊系進行了穗行數基因的QTL定位。在1.02~1.04 bin區段存在控制玉米穗行數的QTL,命名為qKRN1。2年的QTL定位及跨疊系分析結果將qKRN1鎖定在分子標記HND9-umc1297之間,遺傳距離為2.7cM,該穗行數QTL的鑒定,為進一步精細定位或克隆相應基因奠定了基礎。
關鍵詞:玉米;染色體片段置換系;穗行數;數量性狀
中圖分類號: S513.03文獻標志碼: A文章編號:1002-1302(2016)02-0069-04
收稿日期:2015-09-07
基金項目:國家自然科學基金(編號:31501320);襄陽市研究與開發計劃(編號:[2014]12-33);湖北文理學院博士科研啟動基金(編號:31#);湖北省教育廳科學研究計劃中青年人才項目(編號:Q20152601)。
作者簡介:翟立紅(1983—),女,河北邢臺人,講師,主要從事玉米遺傳育種研究。E-mail:zlh_0302@126.com。
通信作者:騰峰,博士,助理研究員,主要從事玉米育種研究。E-mail:tobenumberone@yeah.net。玉米產量是玉米育種學家和遺傳學家共同關注的性狀,穗行數是一個重要的產量組成因子,與產量顯著正相關,探明穗行數形成的遺傳機理有助于玉米產量性狀形成的遺傳機理的闡述,為開展分子設計育種提供理論指導。玉米穗行數是一個數量性狀,受到多基因的控制。隨著分子標記技術的發展和玉米基因組測序結果的出現,在玉米全基因組內檢測控制產量相關QTL取得了一定進展。在20世紀80—90年代,Edwards等就使用同工酶和RFLP標記開展玉米數量性狀基因的研究[1-2]。隨后Veldboom等報道了利用RFLP標記對玉米形態和產量性狀進行QTL定位,鑒定了23個控制形態性狀、35個控制產量性狀的QTL,其中,在1 S(bnl5.62,1.02 bin)、2 S、4 S和4 L上各鑒定了一個穗行數QTL[3]。bnl5.62附近的QTL也被Austin 等所鑒定到,另外他們在其鄰近的umc157附近還檢測到一個穗粗QTL[4]。楊俊品等以4822×5003 的166 個F2:3 家系作為定位群體,共檢測出 59 個分布于10個連鎖群的QTL,其中第1、3 連鎖群較多;在第一染色體的bnlg1083(1.02 bin)、umc1035(1.06 bin)附近發現了2個穗行數QTL[5],這些QTL也與產量有關[6]。嚴建兵等分別利用綜3×87-1的F2:3家系和RIL群體,發現umc1122~bnlg1558(1.06 bin)為一個產量、行數和行粒數QTL簇,umc1169~bnlg1811(1.04 bin)之間存在一個穗行數QTL,bnlg1811(1.04 bin)~umc1124(1.05 bin)之間存在一個百粒質量QTL[7],這些結果也為Li等在爆裂玉米群體中所檢測[8]。湯繼華等利用綜3×87-1的441個永久F2群體,也在第一染色體的umc1122~bnlg1025(1.06 bin)、umc1774~phi26545(1.10~1.11 bin)區段定位了2個穗行數QTL,bnlg1614~bnlg1083(1.02 bin)檢測到一個百粒質量的QTL[9]。Ma等在標記 bnlg2180(1.03 bin)~umc1169(1.04 bin)之間檢測到了一個產量和穗行數的QTL[10]。Blanc等使用4個自交系雜交組配得到的6個F2群體及F2:3家系,在6個組合中均在bnlg1627(1.02 bin)~bnlg176(1.02 bin)之間檢測到產量的QTL[11]。
白葦等利用玉米自交系綜3為受體,衡白522為供體構建的染色體片段置換系群體,進行了玉米產量性狀的定位研究,其中在第一條染色體的1.02~1.04 bin (bnlg1083~bnlg1811)之間定位到了穗行數、穗粗和單穗產量等性狀,認為該區段所含的基因對玉米產量的形成有重大作用,可能是一個控制產量的基因簇[12]。本研究以白葦等定位到穗行數QTL的染色體置換系SIL8為材料,構建了定位區段內的分離群體和跨疊系進行了進一步的QTL鑒定和分析,為分子標記輔助育種提供重要理論依據。
1材料與方法
1.1材料
SIL8是一個穗行數為8~10行的玉米置換系,選自于本實驗室構建的染色體片段置換系群體Z3HBILs,該群體的詳細構建過程見王立秋等的文獻[13]報道。綜3是中國玉米育種中廣泛使用的自交系。與綜3相比,SIL8的遺傳背景非常相似,僅在第一染色體的1.02~1.04 bin(bnlg1083和bnlg1811之間)具有遺傳差異,在表現型上,SIL8的穗行數比綜 3 減少了6~8行,該置換系作為在穗行數上與受體有顯著差異的純系來進行分離群體的構建和精細定位研究。
1.2群體構建和田間種植
以SIL8和綜3雜交,得到區段內F1自交構建第一染色體的1.02~1.04 bin的F2分離群體,于2008年(253個單株)、2009年(593個單株)在河北農業大學(保定)試驗田進行田間種植,行長3 m,行距60 cm,株距25 cm,單粒點播,每行13株,單株考察玉米果穗的穗行數。這2個群體主要用于遺傳連鎖圖譜的構建、QTL的定位和QTL區段內跨疊系的構建。
1.3多態性標記的鑒定和開發
選擇玉米公共遺傳圖譜 IBM Neighbors 2005上已公布的、位于bnlg1083~bnlg1811之間得的所有SSR標記,篩選在綜3和衡白522間具有多態性的標記。由于公共遺傳圖譜上可供利用的標記有限,且多態性不足,利用已公布的玉米自交系B73基因組序列開發更多的、新的標記。SSR標記開發的具體方法如下:從公共網站上www.maizesequence.org上下載區段內的B73的基因組序列,利用網站www.gramene.org提供的SSRIT工具尋找序列中的簡單重復序列,對所搜尋到的簡單重復序列位點,逐個提取簡單重復序列位點前后約 200 bp 的序列,軟件Primer 5.0用于SSR分子標記的引物設計,選擇擴增產物大小150~250 bp的引物,交由上海生工合成。新開發標記的引物以HND標示。
1.4QTL定位
利用在親本間具有多態性的SSR分子標記鑒定分離群體的基因型,使用軟件MAPMAKERNT、Kosambi函數,構建區段內遺傳連鎖圖譜;將群體的基因型和穗行數的表型數據結合,利用軟件WINQTLCART2.5,設置成排列測驗1 000次,復合區間作圖,進行QTL的定位和效應分析。
2結果與分析
2.1多態性標記的篩選與開發
研究利用的置換系SIL8的導入片段在第一染色體的分子標記bnlg1083和bnlg1811之間,將玉米公共圖譜IBM Neighbors 2005上在這2個標記之間的所有SSR標記于親本間篩選多態性。合成的SSR標記有45個,在親本之間具有多態性的標記有19個,即多態性標記比率為42.20%。隨后,利用玉米公共網站www.maizesequence.org上的序列信息,將初步定位區間的序列下載,其物理距離約為10 Mb,含有86個BACs。開發的標記數為119個,新開發的標記中在親本間有多態性的36個,占總開發標記的30.25%,表明綜3和衡白522這2個玉米自交系間的基因組差異較大,多態性標記的開發效率較高。在群體中有較好分離用于群體基因型檢測的有12個,占總開發標記的10.08%。條帶清晰、有多態性(圖1-A)和無多態性(圖1-B)的SSR分子標記檢測見圖1。
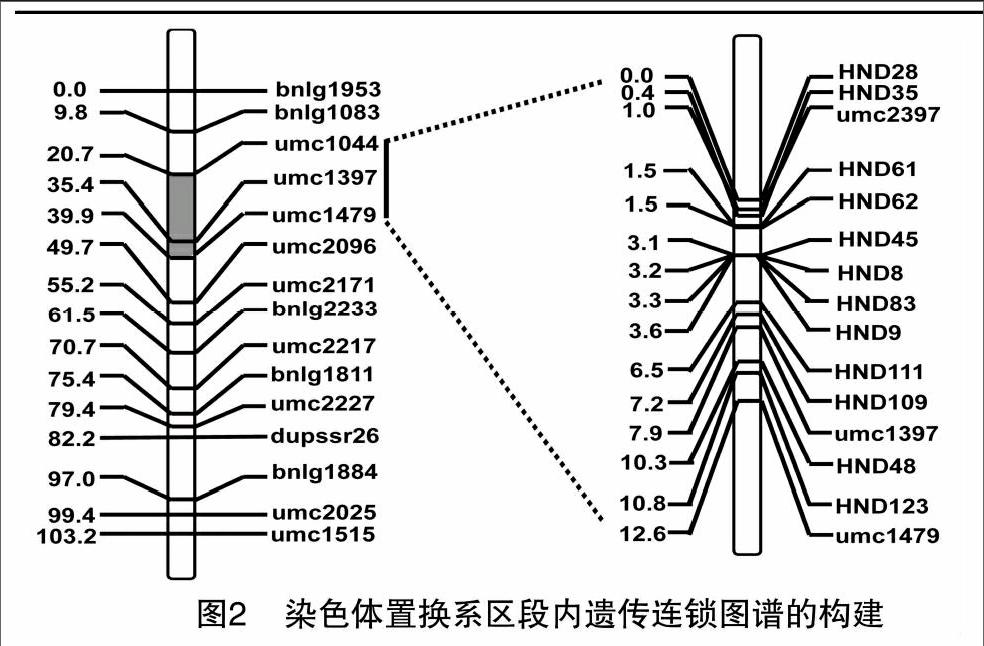
2.2遺傳連鎖圖譜的構建
利用SIL8和綜3雜交構建的區段(bnlg1083~bnlg1811)內F2次級分離群體,分子標記檢測得到的單株基因型進行遺傳連鎖圖譜的構建。2008年,利用保定種植的分離群體(BC4F2)253個單株基因型、15個標記來繪制區段的遺傳連鎖圖譜見圖2(左),遺傳距離總長度為103.2 cM,2個標記最短間距為2.4 cM,最長間距為14.8 cM,平均間距為7.37 cM,標記分布較均勻。2009年,根據2008年初步定位結果,在縮小的區段內進行新的標記的開發和多態性的篩選,利用分離群體(BC5F2)577個單株基因型、15個標記來構建遺傳連鎖圖譜。從圖2(右)可知,遺傳距離總長度為12.6 cM,最短間距達到0.1 cM,最長間距僅2.9 cM,平均間距為0.9 cM,標記密度比較高。
2.3穗行數QTL的定位
2008年,在保定種植分離群體材料,得到180個單株的穗行數表型值,結合群體基因型數據,利用WINQTLCART 2.5軟件設置為步長2 cM,排列測驗1 000次,復合區間作圖法,進行了穗行數QTL的初步定位分析。根據定位結果(圖3-A),在標記umc1044和umc2096之間有2個比較明顯的峰,將目標區段由103.2 cM縮小到29 cM,LOD閾值為3.7,其中
左邊峰的LOD值為4.8,右邊的比較尖一點的峰的LOD值為5.2,貢獻率(R2)達到11%,初步鑒定為主效的穗行數QTL,以加性效應為主。在穗行數QTL初步定位的基礎上,開發定位區間內新的分子標記和篩選其在親本間的多態性并選擇在群體中分離較好的標記。2009年,種植較大的分離群體,最終得到577個單株的穗行數表型值,結合15個標記基因型,開展穗行數QTL的精細定位。定位軟件WINQTLCART 2.5設置為步長0.5 cM,排列測驗1 000次,復合區間作圖法。
從圖3-B看出,2009年穗行數QTL的定位結果在區間內同樣出現明顯的峰,與2008年定位結果相比,位置大致相同,定位區間進一步縮小。經排列測驗得LOD閾值為2.0,第1個峰幾乎與標記HND9所處位置相同,第2個峰在標記 HND109~umc1397之間,遺傳距離為0.7 cM,LOD值為4.9,第3個峰處于標記HND123~umc1479之間,遺傳距離為1.8 cM,LOD值為4.7,貢獻率(R2)為7%,以加性效應為主。
與此同時,利用2008年分離群體中的交換單株,選擇在定位區間內雜合的單株自交,于2009年標記篩選出了11個跨疊系的純系材料(圖4),并對純系材料進行了穗行數的考察。構建的11個跨疊系結合各系穗行數表型值,可以看出,具有HND9~umc1397標記之間片段的純系穗行數為8行或10行,不具有該片段的純系穗行數為14行,該片段與2009年定位結果中的前2個峰所處位置一致。
3討論
玉米產量性狀為多基因控制的數量性狀,基因之間存在著復雜的相互作用,并且受環境的影響較大。穗行數作為產量構成因子之一,相對于復雜的數量性狀而言,穗行數表型數據的獲得相對容易且受環境影響不大,是進行數量性狀定位研究的理想模式性狀之一。本研究在一套染色體片段置換系初步定位的基礎上,選取穗行數顯著減少的置換系SIL8為材料,構建定位區段內的次級分離群體和跨疊系進行穗行數基因的QTL定位與分析,將控制穗行數的qKRN1定位在第1染色體HND9~umc1397之間,2個標記之間的遺傳距離為2.7 cM,在此基礎上,利用玉米基因組序列信息對該區段的候選基因進行了初步預測與分析。本研究定位結果與前人研究結果比較發現,qKRN1與前人利用不同的群體定位到控制穗行數[5,7,10]、穗粗[14]、行粒數[14-15]、百粒質量[9]、穗長[15]、穗粒數[15]、單株籽粒質量[6]、產量[10-11,15]等QTL所在的區間相近,說明此區段可能是一個控制玉米產量及其構成因子的基因簇。然而,利用不同的群體進行產量及其相關性狀QTL定位的結果不盡相同[16-20]。田寶華等對玉米產量相關性狀的QTL進行整合分析,結果發現1號染色體上QTL最多,有174個,其中控制穗行數的有15個[21]。江培順等收集并整合了1994—2012年文獻發表的玉米穗行數、行粒數和粒質量QTL584個,確定了22個穗行數Meta-QTL,得出1.04 bin的粒質量QTL與水稻的GS3基因高度同源,并預測了候選基因[22]。張煥欣等對玉米穗行數進行了全基因組的關聯分析,鑒定了9個與穗行數顯著關聯的SNP,其中1個SNP位于1.02 bin,并將1個含有F-box結構域的生長素受體蛋白為候選基因[23]。fea2編碼一個富亮氨酸的受體蛋白,預測是通過控制花序分生組織來控制玉米穗行數[24]。Wang等對控制玉米68個性狀的1 201個QTL進行了整合,發現了玉米QTL成簇分布的普遍性[25],進一步說明了玉米1號染色體1.02~1.04 bin可能存在著控制玉米產量及其構成因子的重要基因,為開展qKRN1的精細定位和克隆提供了重要的理論支撐。
在利用SIL8材料的同時,對該材料的遺傳背景進行了檢測,發現全基因組內的背景回復率為85.98%。本研究中定位到的穗行數QTL的貢獻率不高,LOD值低,可能的原因:(1)該區段內控制穗行數的QTL可能不止1個且存在互作;(2)背景殘留片段可能與區段內QTL存在互作。因此,為進一步精細定位和克隆qKRN1,需要將SIL8繼續與綜3回交并結合分子標記輔助選擇,在減少背景干擾的基礎上對HND9-umc1397區段的穗行數基因進行深入分析和研究。
參考文獻:
[1]Edwards M D,Helentjaris T,Wright S,et al. Molecular-marker-facilitated investigations of quantitative trait loci in maize:4. Analysis based on genome saturation with isozyme and restriction fragment length polymorphism markers[J]. Theoretical and Applied Genetics,1992,83(6/7):765-774.
[2]Edwards M D,Stuber C W,Wendel J F. Molecular-marker-facilitated investigations of quantitative-trait loci in maize. Ⅰ. Numbers,genomic distribution and types of gene action[J]. Genetics,1987,116(1):113-125.
[3]Veldboom L R,Lee M. Molecular-marker-facilitated studies of morphological traits in maize:Ⅱ. Determination of QTLs for grain yield and yield components[J]. Theoretical and Applied Genetics,1994,89(4):451-458.
[4]Austin D F,lee M,Veldboom L R,et al. Genetic mapping in maize with hybrid progeny across testers and generations:grain yield and grain moisture[J]. Crop Science,2000,40:30-39.
[5]楊俊品,榮廷昭,向道權,等. 玉米數量性狀基因定位[J]. 作物學報,2005,31(2):188-196.
[6]向道權,曹海河,曹永國,等. 玉米SSR遺傳圖譜的構建及產量性狀基因定位[J]. 遺傳學報,2001,28(8):778-784.
[7]嚴建兵,湯華,黃益勤,等. 玉米產量及構成因子主效和上位性QTL的全基因組掃描分析[J]. 科學通報,2006,51(12):1413-1421.
[8]Li Y L,Niu S Z,Dong Y B,et al. Identification of trait-improving quantitative trait loci for grain yield components from a dent corn inbred line in an advanced backcross BC2F2 population and comparison with its F2:3 population in popcorn[J]. Theoretical and Applied Genetics,2007,115(1):129-140.
[9]湯繼華,嚴建兵,馬西青,等. 利用“永久F2”群體剖析玉米產量及其相關性狀的遺傳機制[J]. 作物學報,2007,33(8):1299-1303.
[10]Ma X Q,Tang J H,Teng W T,et al. Epistatic interaction is an important genetic basis of grain yield and its components in maize[J]. Molecular Breeding,2007,20(1):41-51.
[11]Blanc G,Charcosset A,Mangin B,et al. Connected populations for detecting quantitative trait loci and testing for epistasis:an application in maize[J]. Theoretical and Applied Genetics,2006,113(2):206-224.
[12]Bai W,Zhang H,Zhang Z,et al. The evidence for non-additive effect as the main genetic component of plant height and ear height in maize using introgression line populations[J]. Plant Breed,2010,129:376-384.
[13]王立秋,趙永鋒,薛亞東,等. 玉米銜接式單片段導入系群體的構建和評價[J]. 作物學報,2007,33(4):663-668.
[14]李衛華,王洪秋,袁亮,等. 利用單片段代換系群體定位玉米穗部性狀的QTL[J]. 河南農業大學學報,2013,47(2):143-146,181.
[15]Lu M,Xie C X,Li X H,et al. Mapping of quantitative trait loci for kernel row number in maize across seven environments[J]. Molecular Breeding,2011,28(2):143-152.
[16]焦付超,李永祥,陳林,等. 特異玉米種質四路糯的穗行數遺傳解析[J]. 中國農業科學,2014(7):1256-1264.
[17]賈波,管飛翔,謝慶春,等. 玉米產量性狀QTL定位分析[J]. 西南農業學報,2013,26(1):22-25.
[18]彭勃,王陽,李永祥,等. 不同水分環境下玉米產量構成因子及籽粒相關性狀的QTL分析[J]. 作物學報,2010,36(11):1832-1842.
[19]譚巍巍,王陽,李永祥,等. 不同環境下多個玉米穗部性狀的QTL分析[J]. 中國農業科學,2011,44(2):233-244.
[20]譚巍巍,李永祥,王陽,等. 在干旱和正常水分條件下玉米穗部性狀QTL分析[J]. 作物學報,2011,37(2):235-248.
[21]田寶華,王建華. 玉米產量相關性狀QTL的整合及一致性QTL發掘[J]. 分子植物育種,2013(6):752-761.
[22]江培順,張煥欣,呂香玲,等. 玉米產量相關性狀Meta-QTL及候選基因分析[J]. 作物學報,2013,39(6):969-978.
[23]張煥欣,翁建峰,張曉聰,等. 玉米穗行數全基因組關聯分析[J]. 作物學報,2014,40(1):1-6.
[24]Bommert P,Nagasawa N S,Jackson D. Quantitative variation in maize kernel row number is controlled by the FASCIATED EAR2 locus[J]. Nature Genetics,2013,45(3):334-337.
[25]Wang Y,Yao J,Zhang Z F,et al. The comparative analysis based on maize integrated QTL map and meta-analysis of plant height QTLs[J]. Chinese Science Bulletin,2006,51(18):2219-2230.
