離子束誘變雙胚苗水稻的花器性狀及相關(guān)分析
2016-04-11 18:18:40黃雅琴朱慶松李盡哲黃群策
江蘇農(nóng)業(yè)科學(xué) 2016年2期
關(guān)鍵詞:水稻
黃雅琴++朱慶松+++李盡哲++黃群策
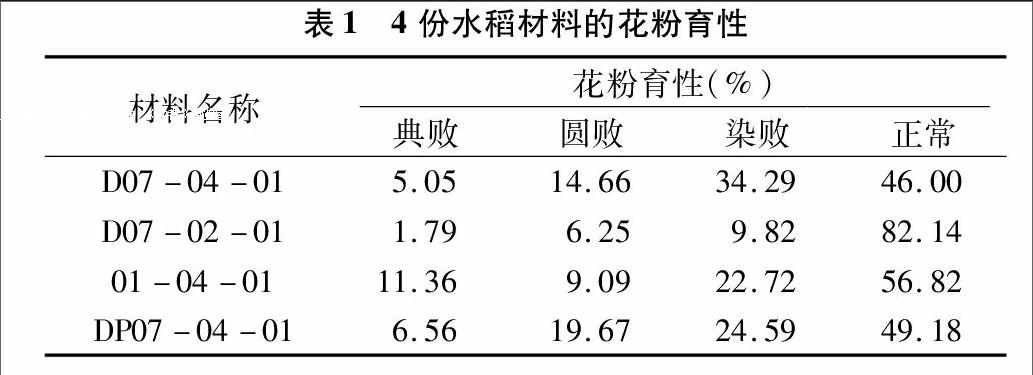

摘要:以離子束誘變所得同源四倍體雙胚苗突變水稻品系、近親系和介導(dǎo)后代為研究材料,對其花粉育性、花器性狀和花器性狀間相關(guān)性進(jìn)行了研究。結(jié)果表明,與四倍體雙胚苗品系相比,二倍體雙胚苗品系的正常花粉率、單穗結(jié)實(shí)率高,花器官偏小,花藥長對單穗結(jié)實(shí)率貢獻(xiàn)大,無芒率高,芒較短;披堿草介導(dǎo)品系的正常花粉率差異不顯著,花器官較大,柱頭長對單穗結(jié)實(shí)率貢獻(xiàn)最大,無芒率最低,芒較長;原始親本在各項(xiàng)指標(biāo)參數(shù)中都差異不顯著。
關(guān)鍵詞:離子束;雙胚苗;水稻;花粉育性;花器性狀;相關(guān)分析;芒
中圖分類號: S511.01文獻(xiàn)標(biāo)志碼: A文章編號:1002-1302(2016)02-0089-03
收稿日期:2015-02-06
基金項(xiàng)目:河南省科技前沿基礎(chǔ)研究項(xiàng)目(編號:82300433202)。
作者簡介:黃雅琴(1986—),女,河南信陽人,碩士,講師,主要從事離子束生物效應(yīng)方面的研究。E-mail:hyaqin88@163.com。
通信作者:李盡哲,碩士,講師,主要從事生物技術(shù)方面的研究。E-mail:xynz1688@163.com。雜交水稻育種從研究方法上可以分為三系法、兩系法和一系法[1]。一系法雜交水稻研究的實(shí)質(zhì)就是挖掘無融合生殖種質(zhì)資源并建立特定的技術(shù)程序,以便固定水稻的雜種優(yōu)勢。前人的研究結(jié)果表明,在多倍體水平挖掘水稻潛在利用價(jià)值,從中篩選多胚苗材料,進(jìn)而尋找無融合生殖種質(zhì)是一條比較有效的途徑[2-4]。20世紀(jì)80年代中期發(fā)展起來的離子束生物技術(shù)具有獨(dú)特的技術(shù)原理和簡單的操作程序,它在作物育種中的誘變效應(yīng)和介導(dǎo)作用已經(jīng)被證實(shí)[5]。筆者所在團(tuán)隊(duì)以低能氮離子束為誘變源對同源四倍體水稻進(jìn)行了誘變,在其后代群體中篩選到具有雙胚苗特征的新材料[6]。同源四倍體水稻具有特殊的生殖發(fā)育現(xiàn)象,在小孢子母細(xì)胞減數(shù)分裂過程的不同階段出現(xiàn)異常現(xiàn)象造成花粉敗育[7],有性繁殖器官花器結(jié)構(gòu)也不同于二倍體水稻[8]。本研究對1個(gè)同源四倍體雙胚苗突變水稻及其2個(gè)近親系、1個(gè)介導(dǎo)品系材料的花粉育性和花器性狀進(jìn)行研究,以同源四倍體雙胚苗水稻為材料開展無融合生殖種質(zhì)挖掘,以推進(jìn)一系法雜交水稻的選育進(jìn)程。
1材料與方法
1.1試驗(yàn)材料
本研究所用材料為同源四倍體雙胚苗突變水稻材料D07-04-01,其近親系D07-02-01和01-04-01,及介導(dǎo)品系DP07-04-01。D07-02-01是以D07-04-01的花粉材料通過花藥培養(yǎng)途徑所獲得的穩(wěn)定二倍體株系;01-04-01是四倍體原始親本,由它通過離子注入后篩選到具有雙胚苗特征的D07-04-01;以D07-04-01為受體,披堿草總DNA為供體在離子束的介導(dǎo)下轉(zhuǎn)基因,在后代群體內(nèi)篩選到四倍體突變單株DP07-04-01。
1.2試驗(yàn)方法
1.2.1I2-KI法觀察成熟花粉粒的育性在抽穗期,每份材料分別隨機(jī)挑選5個(gè)生長良好的未花幼穗,在每穗的上、中、下3個(gè)部位各取3朵穎花分別制成3個(gè)臨時(shí)裝片,用解剖針將每3朵穎花的18個(gè)花藥剝下放在1張載玻片上,滴1滴1%的I2-KI溶液,再用鑷子擠出花粉。在顯微鏡下放大100倍看 30個(gè)標(biāo)準(zhǔn)視野,對成熟花粉粒的育性情況進(jìn)行統(tǒng)計(jì)。根據(jù)I2-KI染色后花粉的表現(xiàn)形態(tài)可分為典敗、圓敗、染敗、正常4種。典敗花粉粒形態(tài)不規(guī)則、透明不染色;圓敗花粉粒圓形,透明不染色;染敗花粉粒圓形,不透明或部分透明,輕度染色;正常花粉粒圓形,不透明,染成較均勻的棕黑色。
1.2.2花器性狀的研究在抽穗期,每份材料分別隨機(jī)挑選30個(gè)生長良好的未花幼穗,在每穗上、中、下3個(gè)部位各取3朵穎花,用游標(biāo)卡尺測量穎花長(x1)、穎花寬(x2),長、寬均測量穎花最長最寬處,精確至0.02 mm,重復(fù)3 次。用圖像掃描法測量柱頭長(x4)、柱頭寬(x6)、子房高(x7)、子房寬(x8)、花藥長(x9)和花藥寬(x10)(單位均為mm) 。計(jì)算2個(gè)2級數(shù)據(jù):穎花長寬比(x3);x4+花柱長=花柱長(x5)。
單穗結(jié)實(shí)率(y):成熟期,每份材料隨機(jī)取生長發(fā)育良好的單穗30個(gè),統(tǒng)計(jì)結(jié)實(shí)數(shù),單穗結(jié)實(shí)率=(單穗的結(jié)實(shí)數(shù)/總穎花數(shù))×100%。用DPS數(shù)據(jù)處理軟件進(jìn)行兩組平均數(shù)t檢驗(yàn),并分析各性狀的差異顯著性;為了比較不同花器性狀對單穗結(jié)實(shí)率的貢獻(xiàn)大小,以單穗結(jié)實(shí)率為依變量,其余10個(gè)花器性狀(x1~x10)為自變量,進(jìn)行逐步回歸分析,分析花器性狀間的相關(guān)性。
隨機(jī)選取1 000粒稻谷進(jìn)行芒長的測定,并根據(jù)所得數(shù)據(jù)實(shí)際情況將其分成10組,計(jì)算每1組的概率。每組的組距為5 mm,組限為0~5 mm、5~10 mm、……、45~50 mm,如芒長恰為5的整數(shù)倍,則將其歸入比其小的組限。
2結(jié)果與分析
2.1花粉育性的比較
同源四倍體水稻與普通的二倍體水稻相比,在小孢子母細(xì)胞減數(shù)分裂時(shí),出現(xiàn)了各種不正常分裂的情況,發(fā)生于單核小孢子期的敗育表現(xiàn)為典敗,發(fā)生于二胞花粉早期的敗育表現(xiàn)為圓敗,發(fā)生于二胞花粉晚期或三胞花粉早期的敗育表現(xiàn)為染敗,這些都最終導(dǎo)致了花粉敗育,因而四倍體水稻的結(jié)實(shí)率要比二倍體水稻低。試驗(yàn)結(jié)果如表1所示,與D07-04-01相比,D07-02-01的典敗、圓敗、染敗分別少3.26百分點(diǎn)、8.41百分點(diǎn)、24.47百分點(diǎn),正常高出36.14百分點(diǎn);01-04-01的典敗和正常分別高出6.31百分點(diǎn)、10.82百分點(diǎn),圓敗和染敗少5.57百分點(diǎn)和11.57百分點(diǎn);DP07-04-01的典敗、圓敗、正常分別高出1.51百分點(diǎn)、5.01百分點(diǎn)、3.18百分點(diǎn),染敗少9.7百分點(diǎn)。由此可見,D07-02-01的正常花粉率最高,D07-04-01、01-04-01和DP07-04-01的敗育類型均主要為染敗。
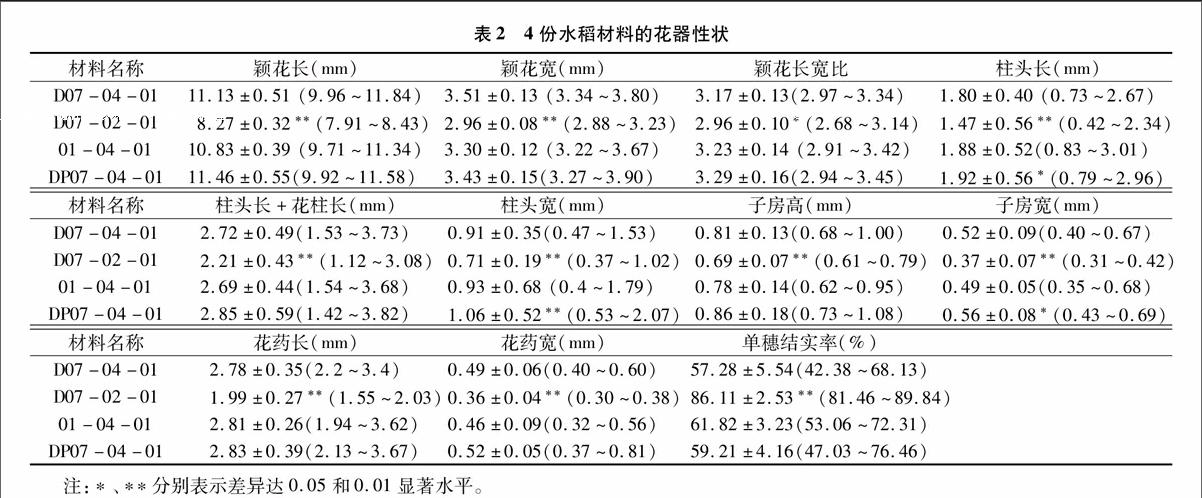
2.2花器性狀的比較
由表2可知,在花器性狀方面,同D07-04-01相比,D07-02-01與其差異顯著或極顯著,其次是DP07-04-01,01-04-01與其差異最小。D07-02-01在研究的11個(gè)花器性狀中,與D07-04-01差異達(dá)極顯著水平的有穎花長、穎花寬、柱頭長、花柱長+柱頭長、柱頭寬、子房高、子房寬、花藥長、花藥寬和單穗結(jié)實(shí)率共10個(gè)參數(shù),其中僅單穗結(jié)實(shí)率高出50.33百分點(diǎn),其他均低;穎花長寬比達(dá)顯著差異(P<0.05),少了6.62%,僅為2.96。DP07-04-01的11個(gè)花器性狀中僅柱頭寬達(dá)到極顯著差異,高出16.48百分點(diǎn);達(dá)到差異顯著水平的有柱頭長和子房寬,分別高出6.67百分點(diǎn)、7.69百分點(diǎn);其他參數(shù)均未達(dá)到差異顯著性水平。01-04-01 的11個(gè)花器性狀均未達(dá)到差異顯著性水平,其中花柱長+柱頭長、子房高、子房寬、花藥寬、穎花長和穎花寬共6個(gè)參數(shù)低于對照,減少最多的是花藥寬,高達(dá)6.12%;柱頭長、柱頭寬、花藥長、穎花長寬比和單穗結(jié)實(shí)率、共5個(gè)參數(shù)高于對照,增加最多的是柱頭長,達(dá)4.44%。
由以上分析可見,在花器性狀上,二倍體雙胚苗材料與四倍體雙胚苗材料之間的差異十分顯著,可見染色體數(shù)目的差異能造成花器性狀的顯著差異;披堿草介導(dǎo)品系由于獲得了披堿草的部分優(yōu)良特性,抗逆能力強(qiáng),花器性狀參數(shù)增大,柱頭寬大,更容易接受花粉而受孕。
2.3花器性狀間的相關(guān)分析
D07-04-01的逐步回歸方程為:
y=0.024 9-0.005 2x2+0.003 3x5-0.007 1x8+0.000 7x9+38.956 2x10(R=0.938 2**,SSE=0.001 1)。
由方程可知,單穗結(jié)實(shí)率與穎花寬、柱頭長+花柱長、子房寬、花藥長、花藥寬相關(guān),其中與柱頭長+花柱長、花藥長、花藥寬呈正相關(guān),與穎花寬、子房寬呈負(fù)相關(guān)。單穗結(jié)實(shí)率與
柱頭長+花柱長、花藥長、花藥寬的偏相關(guān)系數(shù)分別為0.584 1*、0.861 2**、0.830 3**。由此可見,對單穗結(jié)實(shí)率的貢獻(xiàn)上,花藥長>花藥寬>柱頭長+花柱長。
D07-02-01的逐步回歸方程為:
y=-0.013 9+0.003 6x4+0.007 2x6-0.002 2x8+28.764 8x9(R=0.924 3**,SSE=0.000 8)。
由方程可知,單穗結(jié)實(shí)率與柱頭長、柱頭寬、子房寬、花藥長相關(guān),其中與柱頭長、柱頭寬、花藥長呈正相關(guān),與子房寬呈負(fù)相關(guān)。單穗結(jié)實(shí)率與柱頭長、柱頭寬、花藥長的偏相關(guān)系數(shù)分別為0.554 6、0.659 6*、0.692 1*。通過比較偏相關(guān)系數(shù)大小可知,對單穗結(jié)實(shí)率的貢獻(xiàn)上,花藥長>柱頭寬>柱頭長。
01-04-01的逐步回歸方程為:
y=0.021 5-0.005 7x3+0.001 4x5+0.012 9x6+40.560 1x9(R=0.957 4*,SSE=0.000 5)。
由方程可知,單穗結(jié)實(shí)率與穎花長寬比、柱頭長+花柱長、柱頭寬、花藥長相關(guān),其中與柱頭長+花柱長、柱頭寬、花藥長呈正相關(guān),與穎花長寬比呈負(fù)相關(guān)。單穗結(jié)實(shí)率與柱頭長+花柱長、柱頭寬、花藥長的偏相關(guān)系數(shù)分別為0.786 3**、0.742 2*、0.624 5。由此可見,對單穗結(jié)實(shí)率的貢獻(xiàn)上,柱頭長+花柱長>柱頭寬>花藥長。
DP07-04-01的逐步回歸方程為:
y=-0.014 7-0.001 3x3+0.009 3x4-0.005 2x8+0.001 1x9+44.783 4x10(R=0.968 4**,SSE=0.001 0)。
由方程可知,單穗結(jié)實(shí)率與穎花長寬比、柱頭長、子房寬、花藥長、花藥寬相關(guān),其中與柱頭長、花藥長、花藥寬呈正相關(guān),與穎花長寬比、子房寬呈負(fù)相關(guān)。單穗結(jié)實(shí)率與柱頭長、花藥長、花藥寬的偏相關(guān)系數(shù)分別為0.8053**、0.7841**、0.6522。由此可見,對單穗結(jié)實(shí)率的貢獻(xiàn)上,柱頭長>花藥長>花藥寬。
2.4芒的特征比較
觀察每份材料的稻谷后發(fā)現(xiàn)并非每粒都有芒,因此它們都屬于“部分有芒型”,有芒的稻谷,芒的長度也大小不一。D07-04-01無芒率為15.79%,有芒率為84.21%,芒長變幅在0.5~28.8 mm;D07-02-01無芒率為67.96%,有芒率為32.04%,芒長變幅在0.5~11.3 mm;01-04-01無芒率為18.74%,有芒率為81.26%,芒長變幅在0.5~26.3 mm;DP07-04-01無芒率為13.45%,有芒率為86.55%,芒長變幅在0.5~29.3 mm。芒長概率分布如圖1所示,它們的概率密度都整體呈下降趨勢,概率密度最大的都是(0~5 mm)這一組,分別為0.394 5、0.789 5、0.402 0和0.362 7,可見芒長≤5 mm的最多。值得注意的是,唯一的一個(gè)二倍體雙胚苗水稻品系D07-02-01,它的芒長最大為11.3 mm,因此,15~20 mm、20~25 mm、25~30 mm這3個(gè)組限的概率均為零。從整個(gè)稻穗上考察不同長度芒的分布,4個(gè)雙胚苗系列材料都具有這樣的特點(diǎn),即枝梗頂端上部穎花的芒較長,數(shù)量少;枝梗下部穎花芒較短,甚至無芒。由此可見,染色體組減半后,無芒率明顯上升,并且芒長縮短,這是有利變異,有利于稻谷的收割和加工。
3結(jié)論與討論
離子束生物工程是20世紀(jì)80年代在我國興起的,其原理是利用低能離子束注入生物體,質(zhì)量沉積、能量沉積和電荷交換導(dǎo)致生物材料產(chǎn)生一定的生物學(xué)效應(yīng)。離子束生物技術(shù)發(fā)展起來后,在農(nóng)作物育種和遺傳改良上得以廣泛應(yīng)用。我們利用低能氮離子束對同源四倍體水稻進(jìn)行注入處理,從其后代群體內(nèi)中篩選到具有雙胚苗遺傳特性的突變株,通過對D07-04-01、D07-02-01、01-04-01和DP07-04-01這4個(gè)雙胚苗系列材料的花粉育性、花器性狀和芒長分布進(jìn)行比較研究,發(fā)現(xiàn)二倍體雙胚苗品系D07-02-01的正常花粉率最高、花器官偏小、單穗結(jié)實(shí)率最高、花藥長對單穗結(jié)實(shí)率貢獻(xiàn)最大、無芒率最高、芒較短;DP07-04-01這一披堿草介導(dǎo)品系的花器官較大、柱頭長對單穗結(jié)實(shí)率貢獻(xiàn)最大、無芒率最低、芒較長;01-04-01這一原始親本與D07-04-01在各項(xiàng)指標(biāo)參數(shù)中都差異不明顯。已進(jìn)行的苗位和胚位研究發(fā)現(xiàn),D07-02-01的苗位、胚位類型豐富,多胚率高[9]。通過掃描電鏡的觀察發(fā)現(xiàn)D07-02-01的堊白粒率、堊白度和堊白大小均最小,稻米的自然橫斷面中部無明顯輻射狀,裂痕少且淺,“粗短形”的不規(guī)則多邊形胚乳細(xì)胞與淀粉粒分布均勻,多面體狀淀粉粒呈現(xiàn)典型的晶體結(jié)構(gòu),數(shù)目多且排列致密,球形單粒淀粉粒和塊狀不規(guī)則的復(fù)粒淀粉體數(shù)目比較少,相對結(jié)晶度高,直鏈淀粉含量低[10]。綜合以上研究認(rèn)為,二倍體雙胚苗品系D07-02-01雙胚苗頻率高,結(jié)實(shí)率高,加工品質(zhì)優(yōu)良,具有進(jìn)一步挖掘無融合生殖資源的潛在價(jià)值,值得進(jìn)一步研究開發(fā)。
參考文獻(xiàn):
[1]袁隆平. 雜交水稻的育種戰(zhàn)略設(shè)想[J]. 雜交水稻,1987,2(1):1-3.
[2]黃群策. 被子植物的無融合生殖[M]. 福州:福建科學(xué)技術(shù)出版社,2000:16-19.
[3]黃群策,孫梅元,鄧啟云.多倍體水稻及其潛在價(jià)值[J]. 雜交水稻,2001,16(1):4-6.
[4]黃群策. 稻屬植物染色體組多倍化的潛在價(jià)值[J]. 鄭州大學(xué)學(xué)報(bào):理學(xué)版,2008,40(2):108-113.
[5]余增亮. 離子束生物技術(shù)引論[M]. 合肥:安徽科學(xué)技術(shù)出版社,1998:1-3.
[6]黃群策,代西梅. 低能氮離子束對不同倍性水稻的誘變效應(yīng)[J]. 雜交水稻,2004,19(3):60-64.
[7]代西梅,黃群策,李國平,等. 同源四倍體水稻花粉的發(fā)育特征[J]. 中國水稻科學(xué),2006,20(2):165-170.
[8]黃雅琴,黃群策,趙帥鵬. 紫粳4x和紫粳2x水稻品系在3個(gè)植物學(xué)性狀上的差異性比較[J]. 河南農(nóng)業(yè)大學(xué)學(xué)報(bào),2009,43(4):359-363.
[9]黃雅琴,黃群策,燕曉陽. 水稻雙胚苗種子的特征研究[J]. 鄭州大學(xué)學(xué)報(bào):理學(xué)版,2009,41(4):97-102.
[10]黃雅琴,黃群策,燕曉陽,等. 雙胚苗水稻種子的掃描電鏡觀察[J]. 核農(nóng)學(xué)報(bào),2010,24(4):662-667,674.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
