吉林蛟河不同演替階段針闊混交林木本植物幼苗空間分布與年際動態
2016-04-14 03:03:53張新娜張春雨趙秀海
生態學報 2016年23期
關鍵詞:物種
閆 琰,姚 杰,張新娜,張春雨,趙秀海
北京林業大學森林資源與生態系統過程北京市重點實驗室,北京 100083
吉林蛟河不同演替階段針闊混交林木本植物幼苗空間分布與年際動態
閆 琰,姚 杰,張新娜,張春雨,趙秀海*
北京林業大學森林資源與生態系統過程北京市重點實驗室,北京 100083
幼苗的空間分布對群落更新具有重要意義。于2011年夏季在蛟河地區3個不同演替階段針闊混交林樣地內共設置了451個種子雨-幼苗觀測樣站,并于2012—2014年對幼苗樣方內胸徑< 1 cm的木本植物幼苗進行了連續3a的調查。對木本植物幼苗的數量分布以及水曲柳(Fraxinusmandshurica)、五角楓(Acermono)、杉松(Abiesholophylla)和東北楓(Acermandshuricum) 4個主要樹種幼苗與大樹的空間關系及其年際變化進行了分析,并用Syrjala檢驗分析了樣站尺度上幼苗密度、豐富度空間分布在年際間的差異。結果顯示:(1)木本植物幼苗的數量分布在不同群落和年際之間都表現出明顯的差異,樣站尺度上3個樣地內的幼苗密度存在著較大的空間變異性,而幼苗豐富度的空間異質性較低。表明幼苗分布的空間異質性對幼苗密度有著重要影響。(2)樣站尺度上幼苗數量和物種數的空間分布在不同年份之間是存在差異的。在一定程度上證實了種子產量、擴散方式和群落組成對幼苗空間分布的影響。(3))從死亡幼苗與大樹空間關系來看,4個主要樹種的死亡幼苗與成樹表現出相似的分布格局,這既表明成體植株的空間分布特征也能夠影響幼苗的空間分布格局,也從側面說明了負密度制約效應對幼苗空間分布的影響。
幼苗; 空間分布; 年際動態; 演替階段; 蛟河
天然林更新主要是指植物從開花,結實到種子的生產、擴散和萌發,以及幼苗的定居、存活、生長和建成的生態學過程[1-2]。其中,幼苗被認為是森林更新過程對外界環境最敏感的時期[3- 4],其建植成功與否直接關系到未來植物群落的更新[5]。與此同時幼苗定居對植被的更新具有篩選作用[6],通過影響植物種群的數量組成、分布格局來影響森群落動態變化,最終決定了森林的長期演替格局,維系了森林生態系統植被的多樣性[7]。
空間格局與生態學過程之間的相互作用是群落發展的動力[8]。幼苗作為林下生態系統的重要組分,是木本植物生活史中生死過程和動態特性最明顯的時期[9],其空間分布格局會對幼苗的自然更新過程會產生重要的影響。在此過程中,一些機制能夠促進生物多樣性的維持[5]。Swamy等認為,根據Janzen-Connell假說,樹種的更新格局總是偏斜并遠離于母樹[10]。這是由于寄主特異性天敵,如病原體、專性植食者等,容易從成年個體擴散到離它們較近的后代;同時,由于大多數樹種的種子散布在母樹周圍,分布于母樹附近的同種幼苗個體過于密集,病原體和專性植食者的繁殖很可能會加劇,使得幼苗因負密度制約(Negative density dependence)而導致大量死亡[11-12]。這兩個過程使幼苗的存活率隨離母樹或同種成樹距離的增加而升高,在制約同種幼苗增補的同時,為其他物種的幼苗更新提供了空間,從而維持了物種共存。此外,Queenborough等也認為幼苗的空間格局對物種的長期分布格局有著重要影響[13]。
另一方面,森林的更新格局與林分的空間異質性有重要的關系[8]。在不同類型的群落中,地上成體植株的空間異質性為幼苗的增補、定植等過程提供了重要基礎,影響著整個群落的更新特征和演替空間模式[14]。然而,以往對幼苗空間分布格局的研究多集中于特定群落類型[15- 17],對不同演替階段群落中幼苗空間分布格局的研究鮮見報道。
以闊葉紅松(Pinuskoraiensis)林為演替頂級的次生針闊混交林是我國北溫帶地區重要的地帶性植被類型,吉林省蛟河地區是闊葉紅松林的主要分布區之一。早期的人為干擾,如過渡采伐、放牧等活動使大量原始森林遭到破壞,退化成次生林群落。近年來在國家“天然林保護”與“退耕還林”等生態政策的推動下,這些林分形成了針闊混交林次生演替過程中的不同群落類型。盡管如此,森林的天然更新仍然是當地植被恢復過程中面臨的首要難題。因此,本文以吉林省蛟河地區不同演替階段的針闊混交林固定監測樣地為基礎,對2012—2014年連續3 a的木本植物幼苗調查數進行了初分析,旨在:(1)探討木本植物幼苗的空間分布規律及其年際間差異;(2)探討木本植物幼苗和成樹在空間分布上的相關性規律。以揭示溫帶地區不同演替階段森林群落的更新規律,為該地區生態恢復提供理論基礎。
1 研究地概況與研究方法
1.1 研究區概況
蛟河林業試驗區管理局林場位于吉林省蛟河市前進鄉境內(43°51′—44°05′N,127°35′—127°51′E,海拔330—1176 m),屬長白山系張廣才嶺山脈。該地區植被屬于長白山植物區系,天然次生針闊混交林和原始闊葉紅松林是當地的主要植被類型[18]。喬木層樹種主要包括紅松、杉松、東北楓、五角楓、胡桃楸(Juglansmandshurica)、水曲柳和紫椴(Tiliaamurensis)等。林下灌木層樹種主要有暴馬丁香(Syringareticulatavar.mandshurica)、毛榛(Corylusmandshurica)、東北山梅花(Philadelphusschrenkii)和刺五加(Acanthopanaxsenticosus)等。主要草本有美漢草(Meehaniafargesii)、狹葉蕁麻(Urticaangustifolia)、荷青花(Hylomeconjaponica)、華東蹄蓋蕨(Athyriumniponicum)和猴腿蹄蓋蕨(Athyriumbrevifrons)等。該地區處于中緯度北溫帶,屬受季風影響的溫帶大陸性氣候,春季干旱而多大風,夏季炎熱而多雨水,秋季短暫而多晴天,冬季寒冷漫長。年平均氣溫3.8℃,最熱月7月日均溫為21.7℃,最冷月1月份平均溫度-18.6℃。降水多集中在夏季,約為700—800 mm。土壤類型主要為暗棕色森林土[19]。
1.2 樣地建立與調查
根據CTFS(Center for Tropical Forest Science)的技術規范(http://www.ctfs.si.edu/),2010年在研究區內根據林分的發育階段和樹種組成,選取3種不同發育階段的針闊混交林建立固定監測樣地,包括中齡林(21.84 hm2,420 m×520 m)、成熟林(42 hm2,500 m×840 m)和老齡林(30 hm2,500 m×600 m),樣地概況如表1所示。3塊樣地形成了一個由次生林逐漸向頂級植被類型過渡的典型演替系列。調查記錄各樣地內所有胸徑(DBH) ≥1 cm的植株個體物種名稱、胸徑(cm)、樹高(m)、枝下高(m)和冠幅(m),并給每個植株掛牌、定位。
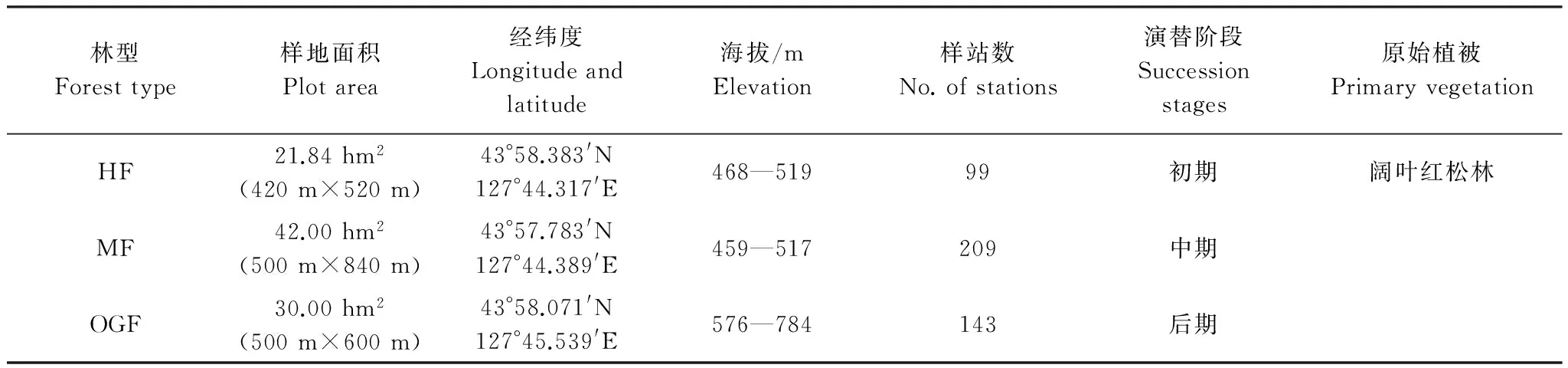
表1 吉林蛟河森林永久監測樣地概況
HF: 中齡林half-mature forest; MF: 成熟林mature forest; OGF: 老齡林old-growth forest
1.3 幼苗樣方設置
2011年7月采用機械布點的方法,在3個樣地內共設置了451個“種子雨-幼苗”觀測樣站,其中中齡林99個、成熟林209個、老齡林143個(圖1)。每個樣站由1個0.8 m×0.8 m的種子雨收集器及其四周距其1 m處的4個1 m×1 m幼苗小樣方組成(圖2)。
圖1 樣站分布示意圖Fig.1 The layout of monitoring stations
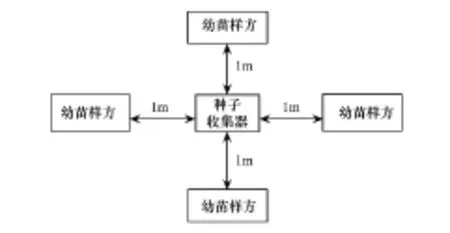
圖2 樣站幼苗小樣方設置示意圖Fig.2 Quadrat layout at each of the sampling station
1.4 幼苗調查
對苗區內胸徑小于1 cm的木本植物幼苗進行調查,記錄每棵幼苗的物種名稱、年齡、高度和基徑,同時對每顆幼苗進行掛牌標記。幼苗的年齡是根據芽鱗痕的個數來確定。幼苗高度為自然狀態下基部到最高頂芽的距離。2012年7—8月進行了第1次幼苗調查工作,以后每年7—8月復查調查1次,并記錄幼苗的出生和死亡狀態[20]。本文選取2012—2014年間的3次幼苗調查數據進行分析。
1.5 數據分析方法
根據各樣站內的幼苗數量(N)劃分為不同的等級:N0=0; 0
選取Syrjala檢驗[21],來分析樣站尺度幼苗密度和豐富度的空間分布在年際間的差異。Syrjala檢驗是通過計算兩個樣本的Cramer-von Mises檢驗來分析兩樣本之間的空間分布差異。Syrjala檢驗的零假設是在整個研究區域內,兩樣本的正態分布是一致的。設在取樣點k的坐標為(xk,yk),di(xk,yk)為第i個種群k點的密度值。構建一個獨立于種群大小的檢驗,所觀測到的密度數據首先被標準化:
gamma.i(xk,yk) =di(xk,yk)/Di
式中,Di是di(xk,yk)觀測值的總和。第i個種群在點(xk,yk)處的累積分布的值,也就是GAMMA.i(xk,yk),是所有正態密度觀察值的總和,gamma.i(xk,yk)。由Syrjala檢驗零假設的統計量為兩個種群之間累積分布的差值的平方,然后所有樣本點求和,即,
psi = sum{GAMMA.1(xk,yk) - GAMMA.2(xk,yk)}2
psi是根據研究區域矩形的角點而改變的,psi被計算4次,每1個角點1次,所得的平均psi值用來做檢驗提供假。觀察到的psi顯著性尺度是由它在所有2K個成對排列所得的檢驗統計值中的序列位置而決定的。
本文中所有數據分析與繪圖均采用國際通用軟件R- 3.2.0(http://www.r-project.org) 和 Microsoft Excel 2010完成。
2 結果與分析
2.1 種子雨組成
2012—2014年間,中齡林共收集到14個物種的種子,隸屬于8科10屬; 成熟林共收集到21個物種的種子,隸屬于11科15屬; 老齡林共收集到20個物種的種子,隸屬于11科14屬20種植物(表2)。3個樣地中,多數物種的種子雨密度都是在2013年達到峰值,但也有少數物種如中齡林中的杉松、胡桃楸,成熟林中的遼椴和老齡林中的千金榆等的種子雨密度卻在2013年最低(表2)。
2.2 幼苗組成的空間分布
研究期間,3個樣地內調查到的木本植物幼苗均為該樣地存在的主要樹種,沒有發現其他樹種的幼苗。樣地內幼苗總數都在2013年達到最大,2014年幼苗數量最少,約為2012年、2013年幼苗數量的一半(表3)。2012—2014年間,中齡林共監測到10科11屬18種木本植物幼苗,其中2012年調查到16種1338株幼苗,沒有發現遼椴(Tiliamandshurica)和千金榆; 2013年調查到18個物種的幼苗,共1942株; 而2014年共調查到724株17種幼苗,沒有發現遼椴(表3)。成熟林共監測到9科11屬17種木本植物幼苗,其中2012年調查到17種幼苗,共4041株; 2013和2014年分別調查到5160和 2379株幼苗,但都沒有發現毛榛(Corylusmandshurica,表3)。老齡林3年內均調查到16個相同樹種的幼苗,隸屬于9科10屬,幼苗株樹分別為2199、2229和917(表3)。
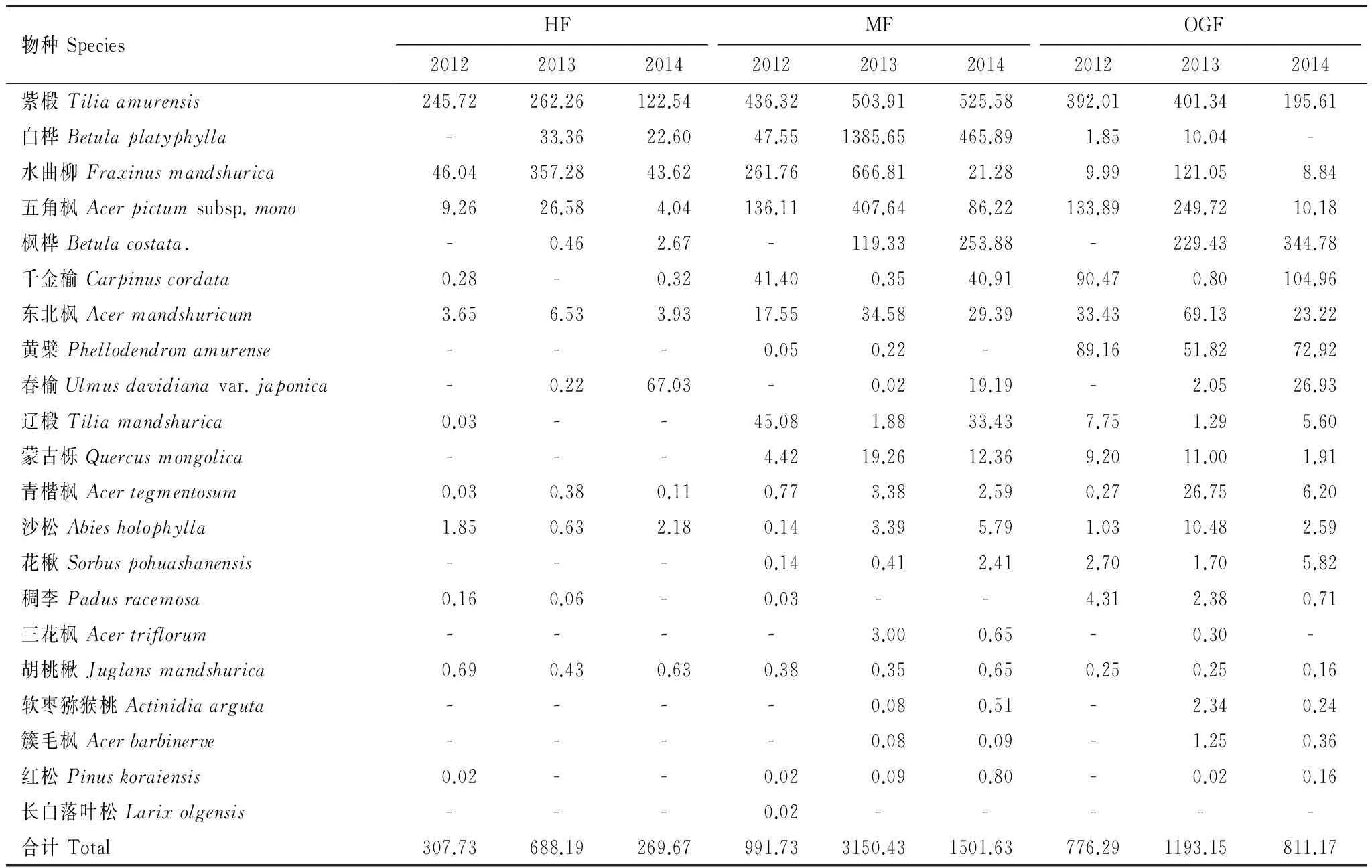
表2 木本植物的種子雨密度/(粒 m-2 a-1)

表3 木本植物幼苗的物種組成、相對多度/%和相對頻度/%
各樣地樹種幼苗數量與出現的樣方數比較結果表明: 水曲柳、五角楓和東北楓等幼苗數量較多的物種,出現的樣方數也相對較多(表3)。3次調查期間,中齡林水曲柳幼苗數量最多,出現樣站的比例均在61.6%以上,其中2012年和2013年出現的樣站數明顯多于2014年; 成熟林水曲柳、五角楓和東北楓出現的樣站數分別達到總樣站數的59.8%、72.7%和37.8%以上; 老齡林中五角楓和東北楓出現的樣站數分別達到總樣站數的62.2% 和48.3% 以上。3個樣地中,盡管2012年的杉松幼苗數量遠高于五角楓,但其出現的樣站數明顯小于五角楓(表3)。
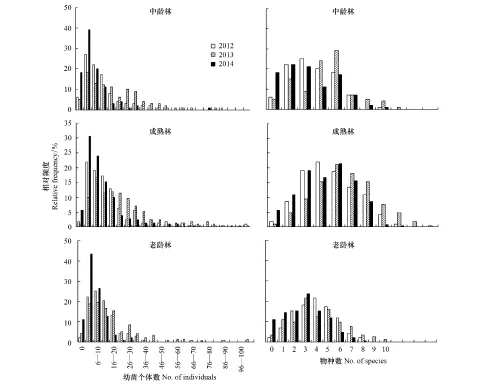
圖3 各樣站出現的幼苗個體數和物種數Fig.3 The number of individuals and species in each of station
根據幼苗出現樣站相對頻率分布可知: 在2012—2014年間,3個樣地中分絕大多數樣站出現幼苗數為1—10株,并且都呈現出偏峰現象; 2014年3個樣地均以幼苗數為1—5株的樣站最多,并且幼苗數為0株和1—5株的樣站數量均明顯高于2012年和2013年; 成熟林在2013年幼苗數為6—10株的樣站數最多,而老齡林在2012年幼苗數為6—10株的樣站數最多(圖3)。
對樣站中出現幼苗的物種數進行統計發現,中齡林在3次調查期間物種的頻率分布均出現偏鋒現象,而且年際間差異明顯: 2012年有6個樣站中沒有發現任何樹種幼苗,85個樣站(占樣站總數的85.9%)中出現的物種數小于5; 2013年出現1—5個幼苗物種的樣站數最多,為29個,遠高于2012年和2014年調查結果; 2014年有18個樣站中沒有出現任何幼苗,61個樣站(樣站總數的61.6%)發現0—2個物種(圖3)。而成熟林和老齡林物種頻率分布的年際差異不明顯,且均為正態分布,樣站數最多的為3—4個樹種(圖3)。
2.3 幼苗空間分布差異
采用Syrjala檢驗分析了同一樣地內幼苗個體總數、物種數以及4個主要物種幼苗多度的年際間空間分布差異(表4)。結果表明: 中齡林幼苗物種數和個體數的空間分布在2013年和2014年存在顯著差異; 水曲柳、杉松和東北楓幼苗數量空間分布在年際間存在顯著差異,而五角楓幼苗數量在年際間差異不顯著,表明五角楓幼苗的產生和分布在中齡林中比較穩定。成熟林幼苗物種數和幼苗個體總數的空間分布存在顯著的年際差異,水曲柳、五角楓和東北楓幼苗數量的空間分布在年際間差異顯著,而杉松幼苗數量在年際間差異不顯著,表明杉松幼苗的產生和分布在成熟林中比較穩定。老齡林中,2012年幼苗個體總數的空間分布與2013年、2014年差異顯著; 2012年杉松幼苗數量空間分布與2013年差異顯著; 而幼苗的物種數以及水曲柳、五角楓和東北楓幼苗數量的空間分布在年際間變化不顯著,表明老齡林林下幼苗的空間分布比中齡林和成熟林相對穩定。
表4 3個樣地內幼苗個體總數、物種數與4個主要物種幼苗多度年際間空間分布差異的Syrjala檢驗結果
Table 4 Syrjala test values for the annual variation of the number of individuals,the number of species and seedling abundance of four major species in the three forest plots
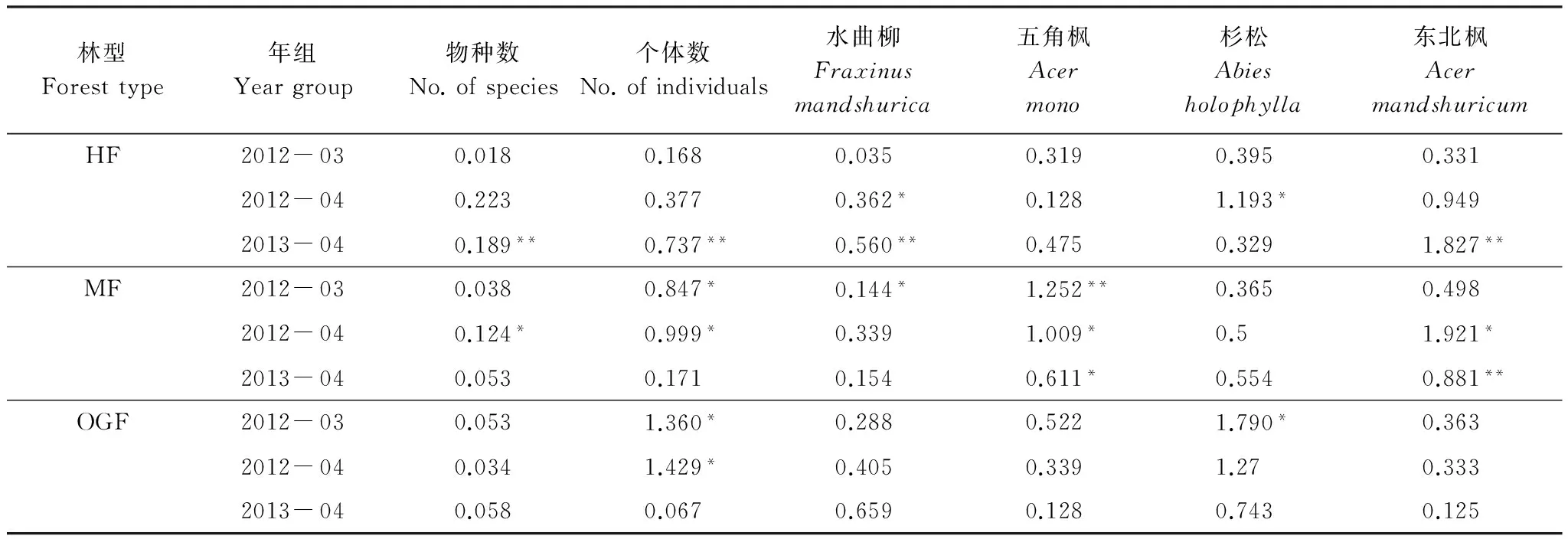
林型Foresttype年組Yeargroup物種數No.ofspecies個體數No.ofindividuals水曲柳Fraxinusmandshurica五角楓Acermono杉松Abiesholophylla東北楓AcermandshuricumHF2012-030.0180.1680.0350.3190.3950.3312012-040.2230.3770.362*0.1281.193*0.9492013-040.189**0.737**0.560**0.4750.3291.827**MF2012-030.0380.847*0.144*1.252**0.3650.4982012-040.124*0.999*0.3391.009*0.51.921*2013-040.0530.1710.1540.611*0.5540.881**OGF2012-030.0531.360*0.2880.5221.790*0.3632012-040.0341.429*0.4050.3391.270.3332013-040.0580.0670.6590.1280.7430.125
2.4 主要樹種死亡幼苗與成樹空間分布
對3個樣地內水曲柳、五角楓、杉松和東北楓的死亡幼苗和成樹空間分布進行比較可以發現: 盡管3個樣地中死亡的幼苗基本分布在母樹附近,并且整體格局與母樹保持一致,但死亡幼苗的空間分布格局在年際間存在一定差異(圖4—圖6); 2013—2014年間五角楓和東北楓的死亡幼苗在3個樣地中分布較為廣泛(圖4,圖5);老齡林中,杉松在2013—2014年間死亡的幼苗在樣地中分布的較少(圖6)。
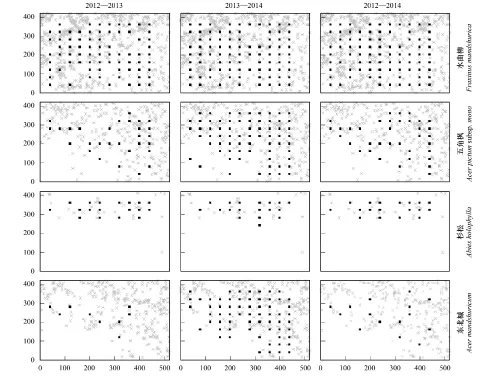
圖4 2012—2013年、2013—2014年和2012—2014年間中齡林4個主要樹種的死亡幼苗及其胸徑大于20 cm母樹的空間分布格局Fig.4 Spatial distribution patterns of the dead seedlings for the four main species during the year of 2012—2013,2013—2014 and 2012—2014,and their parent trees with DBH ≥20 cm in the HF灰色叉號為母樹空間分布,黑色方塊為出現該樹種幼苗死亡的樣站分布
3 討論
3.1 幼苗的數量分布
群落幼苗與地上成體植株的空間分布格局決定了群落更新特征,成體植株的空間分布特征對幼苗建成具有重要影響[14]。因此,幼苗的空間分布對群落更新具有重要意義。本文通過對蛟河地區不同演替階段針闊混交林樣地內木本植物及幼苗的調查分析,發現3個樣地內調查到的木本植物幼苗均為該樣地存在的主要樹種,沒有發現其他樹種的幼苗。木本植物幼苗的數量分布在不同演替階段之間都表現出明顯的差異: 水曲柳在演替初期和中期的林分中出現頻率較大,五角楓在演替中期和后期的林分中中出現頻率最大,東北楓在3個樣地中出現頻率都比較大; 而胡桃楸、杉松和紅松的幼苗在3個樣地中出現的頻率都很低。這一方面可能是由于不同演替階段的林分中,樹種的數量和組成存在差異,使得不同物種的幼苗分布在不同群落間存在差異; 另一方面,嚙齒類動物和人類對紅松、胡桃楸等種子的取食和采摘也會對幼苗的分布造成不同程度的影響。研究期間在3個樣地內水曲柳、五角楓和東北楓的種子數量在所有樹種中均排在前5位,而胡桃楸、杉松和紅松的種子數量較少(未發表數據),這與陳貝貝等部分研究結果一致[22];此外,幼苗的空間分布可能與種子的傳播特性有關,水曲柳、五角楓和東北楓的種子主要通過風力傳播,良好的擴散能力也能使得種子和幼苗在樣地內廣泛分布[23],而胡桃楸、紅松和杉松的種子主要通過重力傳播,擴散能力有限。
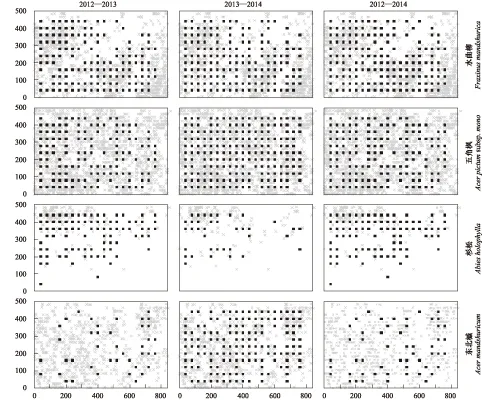
圖5 2012—2013年、2013—2014年和2012—2014年間成熟林4個主要樹種的死亡幼苗及其胸徑大于20 cm母樹的空間分布格局Fig.5 Spatial distribution patterns of the dead seedlings for the four main species during the year of 2012—2013,2013—2014 and 2012—2014,and their parent trees with DBH ≥20 cm in the MF灰色叉號為母樹空間分布,黑色方塊為出現該樹種幼苗死亡的樣站分布
3.2 幼苗密度與豐富度的空間異質性
本研究結果顯示,在4 m2樣站尺度上3個不同演替階段的群落中幼苗密度存在著較大的空間變異性,而幼苗豐富度的空間異質性較低。這與李曉亮等對西雙版納熱帶森林樹種幼苗空間分布研究的部分結果一致[24],他們認為幼苗密度的空間變異是由微生境的空間異質性導致的,微生境的空間變異又會反來影響幼苗的存活[25]。而物種數量由于受群落物種區系控制,空間異質性較低[14]。也有學者認為森林中幼苗空間格局可能比其周圍的環境因素更重要[16],因為幼苗密度的空間格局會影響到幼苗的存活狀態[24]。例如,Janzen和Connell[11- 12]認為分布于母樹附近的幼苗由于受到負密度制約效應會表現出較高的死亡率,而本研究中3個樣地內主要樹種的幼苗都集中于母樹附近(圖3—圖5)。同時,以往的研究已經證實了負密度制約效應是該研究地區幼苗存活的主要驅動機制[20],這在一定程度上可以反映出幼苗分布的空間異質性對幼苗密度的影響。

圖6 2012—2013年、2013—2014年和2012—2014年間老齡林4個主要樹種的死亡幼苗及其胸徑大于20 cm母樹的空間分布格局Fig.6 Spatial distribution patterns of the dead seedlings for the four main species during the year of 2012—2013,2013—2014 and 2012—2014,and their parent trees with DBH ≥20 cm in the OGF灰色叉號為母樹空間分布,黑色方塊為出現該樹種幼苗死亡的樣站分布
3.3 幼苗空間分布的年際差異
不同年份之間幼苗個體數和物種數的空間分布是存在差異的。這主要與種子的產量與擴散有關。首先,種子是幼苗更新的來源,幼苗的數量和分布很大程度上依賴于種子的可獲得性[26],種子數量多的物種跟容易擴散到更多的樣站[27]。另一方面,不同物種的種子擴散方式有很大差異,有些物種的種子在經歷了種子雨擴散之后,還會經歷二次擴散。二次擴散可以降低種子的聚集程度,使種子距離母樹更遠。因此,種子擴散會導致種子萌發位置的改變進而影響幼苗的分布。同時,種子具有休眠的特性,只有光照、溫度等環境因素適宜的時候種子才會萌發,這也影響到幼苗的分布格局。除此之外,不同演替階段群落中的物種組成和林分結構及環境都有很大的差異,因此各林分內幼苗空間分布在年際間的差異也不盡相同。
3.4 死亡幼苗空間分布與成樹關系
除了種子產量和種子擴散方式,成體植株的空間分布特征也能夠影響幼苗的空間分布格局[28]。本研究中,在不同演替階段的群落內,水曲柳、五角楓、杉松和東北楓這4個主要樹種的死亡幼苗在不同年際間的空間分布格局與其母樹分布基本一致。這可能是由于選取的4個物種的母樹在群落中有較多的個體,產生的種子量較多,使母樹附近有足夠的種源,進而使得幼苗與母樹有著相似的空間分布格局[24];另一方面,由于種子的傳播限制,大多數樹種并不能把種子散布到離母樹較遠的地方[29- 30],相反,隨著與母樹距離的增加,種子密度會逐漸降低[31],雖然這4個樹種的種子都依靠風力傳播,但是傳播的距離并不遠,促使幼苗聚集在母樹周圍。因此,在負密度制約效應的作用下,鄰近母樹的幼苗更容易死亡。此外,五角楓、東北楓和沙松的死亡幼苗的空間分布在年際間存在差異,這可能與幼苗的密度有關。3個樣地中,五角楓和東北楓在2013—2014年間死亡幼苗的空間分布要明顯大于2012—2013年,而這兩個物種在2013年的幼苗密度均大于2012年,這從側面進一步證實了負密度制約效應對幼苗空間分布的影響。
[1] 郭秋菊. 擇伐和火干擾對長葉松幼苗更新的影響[D]. 楊凌: 西北農林科技大學,2013.
[2] Rockwood L L. Introduction to Population Ecology. Oxford: Blackwell Publishing,2006
[3] 尹華軍,程新穎,賴挺,林波,劉慶. 川西亞高山65年人工云杉林種子雨、種子庫和幼苗定居研究. 植物生態學報,2011, 35(1): 35- 44.
[4] Du X J,Guo Q F,Gao X M,Ma K P. Seed rain,soil seed bank,seed loss and regeneration ofCastanopsisfargesii(Fagaceae) in a subtropical evergreen broad-leaved forest. Forest Ecology and Management,2007,238(1/3): 212- 219.
[5] 蘇嫄,焦菊英,王志杰. 陜北黃土丘陵溝壑區坡溝立地環境下幼苗的存活特征. 植物生態學報,2014,38(7): 694- 709.
[6] Clark J S,Macklin E,Wood L. Stages and spatial scales of recruitment limitation in southern Appalachian forests. Ecological Monographs,1998,68(2): 213- 235.
[7] 康冰,劉世榮,王得祥,張瑩,劉紅茹,杜焰玲. 秦嶺山地典型次生林木本植物幼苗更新特征. 應用生態學報,2011,22(12): 3123- 3130.
[8] 韓有志,王政權. 天然次生林中水曲柳種子庫的空間格局與過程. 植物生態學報,2002,26(2): 170- 176.
[9] 路興慧. 海南島熱帶低地雨林自然恢復過程中木本植物幼苗功能性狀及增補動態[D]. 北京: 中國林業科學研究院,2012.
[10] Swamy V,Terborgh J,Dexter K G,Best B D,Alvarez P,Cornejo F. Are all seeds equal? Spatially explicit comparisons of seed fall and sapling recruitment in a tropical forest. Ecology Letters,2011,14(2): 195- 201.
[11] Janzen D H. Herbivores and the number of tree species in tropical forests. The American Naturalists,1970,104(940): 501- 528.
[12] Connell J H. On the role of natural enemies in preventing competitive exclusion in some marin animals and rain forest trees//Den Boer P J,Gradwell G R,eds. Dynamics of Population. Wageningen: Center for Agricultural Publishing and Documentation,1971: 289- 312.
[13] Queenborough S A,Burslem D F R P,Garwood N C,Valencia R. Neighborhood and community interactions determine the spatial pattern of tropical tree seedling survival. Ecology,2007,88(9): 2248- 2258.
[14] 尚占環,龍瑞軍,馬玉壽,丁路明. 青藏高原“黑土灘”次生毒雜草群落成體植株與幼苗空間異質性及相似性分析. 植物生態學報,2008,32(5): 1157- 1165.
[15] 韓有志,程志楓,常杰,王永強. 水曲柳人工林下天然更新幼苗的空間格局. 山西農業大學學報,2000,(4): 335- 338.
[16] Armesto J J,Díaz I,Papic C,Willson M F. Seed rain of fleshy and dry propagules in different habitats in the temperate rainforests of Chiloé Island,Chile. Austral Ecology,2001,26(4): 311- 320.
[17] Piao T F,Comita L S,Jin G Z,Kin J H. Density dependence across multiple life stages in a temperate old-growth forest of northeast China. Oecologia,2013,172(1): 207- 217.
[18] Zhao X H,Corral-Rivas J,Zhang C Y,Temesgen H,Gadow K V. Forest observational studies-an essential infrastructure for sustainable use of natural resources. Forest Ecosystems,2014,1: 8.
[19] Zhang C Y,Jin W B,Gao L S,Zhao X H. Scale dependent structuring of spatial diversity in two temperate forest communities. Forest Ecology and Management,2014,316: 110- 116.
[20] Yan Y,Zhang C Y,Wang Y X,Zhao X H,Von Gadow K. Drivers of seedling survival in a temperate forest and their relative importance at three stages of succession. Ecology and Evolution,2015,5(19): 4287- 4299.
[21] Syrjala S E. A statistical test for a difference between the spatial distributions of two populations. Ecology,1996,77(1): 75- 80.
[22] 陳貝貝,趙秀海,倪瑞強,黃珍. 長白山闊葉紅松林喬木樹種幼苗組成與空間分布. 東北林業大學學報,2012,40(10): 39- 42.
[23] 張健,李步杭,白雪嬌,原作強,王緒高,葉吉,郝占慶. 長白山闊葉紅松林喬木樹種幼苗組成及其年際動態. 生物多樣性,2009,17(4): 385- 396.
[24] 李曉亮,王洪,鄭征,林露湘,鄧曉保,曹敏. 西雙版納熱帶森林樹種幼苗的組成、空間分布和旱季存活. 植物生態學報,2009, 33(4): 658- 671.
[25] Baraloto C,Goldberg D E. Microhabitat associations and seedling bank dynamics in a neotropical forest. Oecologia,2004,141(4): 701- 712.
[26] Nathan R,Safriel U N,Noy-Meir S,Schiller G. Spatiotemporal variation in seed dispersal and recruitment near and far from pinus halepensis trees. Ecology 2000,81(8): 2156- 2169.
[27] Chen L,Wang L,Baiketuerhan Y,Zhang C Y,Zhao X H,von Gadow K. Seed dispersal and seedling recruitment of trees at different successional stages in a temperate forest in northeastern China. Journal of Plant Ecology,2013,7(4): 337- 346.
[28] 王蕾. 長白山針闊混交林種子雨動態及擴散限制研究[D]. 北京: 北京林業大學,2010.
[29] 張健,郝占慶,李步杭,葉吉,王緒高,姚曉琳. 長白山闊葉紅松(Pinuskoraiensis)林種子雨組成及其季節動態. 生態學報,2008,28(6): 2445- 2454.
[30] 張玉波,李景文,張昊,鄒大林,武逢平,程春龍,李俊清,李帥英. 胡楊種子散布的時空分布格局. 生態學報,2005,25(8): 1994- 2000.
[31] Willson M F,Whelan C J. Variation of dispersal phenology in a bird-dispersed shrub,cornus drummondii. Ecological Monographs,1993, 63(2): 151- 172.
Spatial distribution and interannual dynamics of tree seedlings at different successional stages in a conifer and broad-leaved mixed forest in Jiaohe,Jilin Province,China
YAN Yan,YAO Jie,Zhang Xinna,ZHANG Chunyu,ZHAO Xiuhai*
KeyLaboratoryforForestResources&EcosystemProcessesofBeijing,BeijingForestryUniversity,Beijing100083,China
The spatial pattern of seedlings plays an important role in community regeneration. Our study was based on 415 seed-seedling census stations established in three large permanent field plots,each representing one of three successional stages in a conifer and broad-leaved mixed forest. We conducted three censuses from 2012 to 2014 for seedlings <1 cm in diameter at breast height and analyzed the quantitative distribution of woody seedlings and the spatial relationship between seedlings and adult trees for four major species:Fraxinusmandshurica,Acermono,Abiesholophylla,andAcermandshuricum. The Syrjala test was used to analyze interannual differences in the spatial pattern of the number of individual seedlings and the number of seedling species at the station level. Our results indicated that(1) there were obvious differences in the quantitative distribution of woody seedlings among communities and years. Seedling density exhibited substantial spatial variability at the station level,whereas spatial heterogeneity of seedling richness was low. These results indicated that spatial heterogeneity of seedling distribution had an important impact on seedling density.(2) Spatial pattern of the number of individual seedlings and the number of seedling species varied among years at the station level. This demonstrated the influence of seed production,dispersal method,and community composition on the spatial distribution of seedlings.(3) Judging from the spatial relationship between dead seedlings and adult trees,dead individuals of the four major seedling species exhibited a similar distribution pattern with that of adult trees,which indicated that the distribution pattern of adult trees might affect the distribution pattern of seedlings,. It further confirmed the negative density dependence of the spatial pattern of seedlings.
seedling; spatial distribution; interannual dynamics; successional stage; Jiaohe
國家“十二五”科技支撐計劃項目(2012BAD22B0203)
2015- 12- 06;
2016- 05- 30
10.5846/stxb201512062442
*通訊作者Corresponding author.E-mail: zhxh@bjfu.edu.cn
閆琰,姚杰,張新娜,張春雨,趙秀海.吉林蛟河不同演替階段針闊混交林木本植物幼苗空間分布與年際動態.生態學報,2016,36(23):7644- 7654.
Yan Y,Yao J,Zhang X N,Zhang C Y, Zhao X H.Spatial distribution and interannual dynamics of tree seedlings at different successional stages in a conifer and broad-leaved mixed forest in Jiaohe,Jilin Province,China.Acta Ecologica Sinica,2016,36(23):7644- 7654.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
