不同放牧強度季節調控下荒漠草原主要植物種群點格局及空間關聯性
2016-04-14 03:03:48孫世賢衛智軍吳新宏郭利彪
生態學報 2016年23期
關鍵詞:關聯
孫世賢,衛智軍,吳新宏,*,姜 超,郭利彪
1 中國農業科學院草原研究所, 呼和浩特 010020 2 內蒙古農業大學生態環境學院, 呼和浩特 010018
不同放牧強度季節調控下荒漠草原主要植物種群點格局及空間關聯性
孫世賢1,衛智軍2,吳新宏1,*,姜 超1,郭利彪1
1 中國農業科學院草原研究所, 呼和浩特 010020 2 內蒙古農業大學生態環境學院, 呼和浩特 010018
種群空間格局是種群自身特性、種間相互關系及環境條件綜合作用的結果。以短花針茅荒漠草原為研究對象,運用Programita軟件,采用Ripley′s K函數和Monte Carlo隨機模擬方法,對短花針茅、無芒影子草和堿韭種群點格局及空間關聯性進行了研究。結果表明:短花針茅在禁牧和過度放牧下集群分布的尺度在增加,禁牧和過度放牧兩種草地利用方式下短花針茅種群的格局具有趨同的趨勢;并且,隨著放牧強度的增大或者是持續放牧的影響,使得隨機分布尺度在逐漸的增大,短花針茅在更大的尺度上才可能為集群分布。春季重牧+夏季重牧+秋季輕牧和全年重度放牧利用下無芒隱子草在較大尺度上才表現為集群分布,并且尺度轉化的臨界點在放牧的影響下有增大的趨勢;重度放牧下堿韭為了適應放牧干擾逐漸向集群分布方向發展,集群分布的尺度在減小以提高種群的穩定性從而抵御過度的干擾。在不同的放牧干擾強度下植物種群具有明顯的響應策略,大致表現為物種的群居性在增強,物種集群分布的尺度在減小以提高種群的穩定性從而抵御過度的干擾。春季休牧+夏季重牧+秋季輕牧處理下種群斑塊化的尺度較大,有利于群落的穩定,因此荒漠草原采用這種利用方法較為合理。
荒漠草原;放牧強度;季節調控;種群點格局;空間關聯性
空間格局是指在環境因子的綜合作用下空間中種群的分布關系。植物在種群、群落和景觀等尺度下均表現出一定的規律性[1]。Watt于1947年發表“植物群落的格局與過程”的開創性論文以來,關于植被格局的研究主要集中在格局與過程的關系方面,如今生態學家們普遍認為空間結構影響著群落動態[2- 3]。種群空間格局的形成與生態過程存在著密切的聯系,種群在空間中分布格局的形成機制可以通過種群空間格局的分析加以解釋[4]。在干擾的影響下,草原植物的生態適應對策因物種類型的差異而不同,進而影響植物對有限資源的競爭,決定了種群不同尺度的空間格局;相反,種群的空間格局及其動態不僅可以反映環境對種群個體的影響,也可指示植物的生態適應對策[5- 7]。目前在放牧干擾下種群格局方面已有一些研究[8- 9],這些研究都在探索過度放牧干擾下種群的衰退機理。隨著人們對草地的過度利用,草地退化已經成為我國北方重要的環境與資源問題之一[10]。
20世紀80年代,任繼周提出“季節畜牧業”,認為中國西部地區草地生長因受氣候影響明顯,必須施行季節放牧[11]。由于不同季節草地生物量和抵抗干擾的能力不同,遵循草地的這種季節性差異規律,研究不同季節性調控利用模式對草地的影響具有理論和現實意義。不同季節調控利用下種群分布格局存在什么樣的差異,種群分布的尺度轉化臨界點有什么不同,那種利用方式下群落更加穩定?這些問題是本研究主要解決的科學問題。荒漠草原生態環境脆弱,且具有較大的波動性,對自然環境和人類生產活動的干擾較敏感[12],目前退化較為嚴重。本文通過對不同放牧強度季節調控利用下對短花針茅、無芒隱子草和堿韭種群點格局變化及種間關聯性進行分析,了解不同放牧強度季節調控下優勢植物種群格局特征及其變化趨勢,以期為進一步揭示荒漠草原退化原因及演替提供理論依據。
1 材料與方法
1.1 實驗區概況
試驗區位于內蒙古錫林郭勒盟蘇尼特右旗朱日和鎮(112°47′16.9″E,42°16′26.2″N)。土壤為淡栗鈣土,有明顯的鈣積層,分布在10—35 cm,腐殖質層厚5—10 cm。試驗區植被以亞洲中部荒漠草原種占主導地位。以短花針茅(Stipabreviflora)為建群種,優勢種為堿韭(Alliumpolyrhizum)和無芒隱子草(Cleistogenessongorica)。主要伴生種有銀灰旋花(Convolvulusammannii)、木地膚(Kochiaprostrata)、細葉韭(Alliumtenuissimum)、阿爾泰狗哇花(Heteropappusaltaicus)、糙隱子草(Cleistogenessquarrosa)、狹葉錦雞兒(Caraganastenophylla)和寸草苔(Carexduriuscula)等。試驗區年均降水量183.0mm,年平均氣溫5.8℃,日照時數3137.3h,年蒸發量2793.4mm,≥5℃的有效積溫3426.0℃,≥10℃的有效積溫2491℃,無霜期177d,屬中溫型氣候。
1.2 主要植物種群的生物生態學特性
短花針茅是多年生密叢植物,四月上旬開始萌動返青,5月下旬至6月中旬抽穗開花進入生長發育盛期。無芒隱子草是強旱生密叢禾草植物,為荒漠草原群落的共建種。無芒隱子草春季返青較晚而秋季枯黃期早,是群落中多年生旱生叢生禾草植物生長期較短的植物。每年4月下旬氣溫上升到10℃時,它才開始萌動。堿韭為典型的旱生植物,鱗莖多枚緊密簇生,圓柱狀;鱗莖外圍包著一層很厚的枯死鱗莖皮,于地表形成保護層,防旱和防熱,減少鱗莖根系暴曬和蒸發水分。鱗莖外面由縱分裂的被膜與葉鞘形成的纖維所緊密包圍。
1.3 實驗設計
放牧試驗于2010開始,設6個處理,每個處理3次重復,共18個放牧試驗小區(表1),采用完全隨機區組設計。放牧試驗地總面積78.5hm2,每個試驗小區面積為5.2 hm2。6個放牧處理分別為春季零放牧(休牧)+夏季重度放牧+秋季適度放牧區(SA1)、春季零放牧(休牧)+夏季適度放牧+秋季重度放牧區(SA2)、放牧季皆為重度放牧區(SA3)、春夏季重度放牧+秋季適度放牧區(SA4)和放牧季皆為適度放牧區(SA5),即每一試驗處理分為春、夏、秋3個放牧季,每一放牧季采用的放牧強度為零放牧、適度放牧和重度放牧中的一種。零放牧、適度放牧和重度放牧區分別放牧蘇尼特羊0只、6只和9只,折算載畜率分別為0、0.87hm2羊-1a-1和0.58hm2羊-1a-1。

表1 試驗設計
1.4 試驗方法
在2013年7月植物生長盛期采集數據,在每個圍欄樣地內隨機設置1個5m×5m的樣方,用1m×1m的樣方框按從左到右,從上到下的順序依次放置(樣方框按正南正北方向放置)25次進行草地群落調查與短花針茅、堿韭和無芒隱子草種群的調查。以樣方框左下角頂點為基點,記錄樣方中每個短花針茅、堿韭和無芒隱子草種群相對位置,以坐標值表示,用坐標值直接表示距離。
1.5 數據分析
Ripley于1977年首先提出點格局分析理論[13],1983年Diggle等人對點格局理論進行了改進與發展[14]。在實踐中,用K(t)函數表征點格局,K(t)主要用下式估計:
式中,A表示樣地面積;n表示群落的個體總數;uij表示第i株植株和第j植株之間的距離,t表示空間尺度,當uij≤t是,It(uij)=1,反之,It(uij)=0;Wij為以點i為圓心、以uij為半徑的圓落在面積A中的弧長與整個圓周的比值,可校正邊界效應引起的誤差。Besag和Kenkel曾建議用L(t)代替K(t),其公式為:
當L(t)=0時,為隨機分布;當L(t)<0時,為均勻分布;當L(t)>0時,為集群分布。
Monte-Carlo擬合檢驗用于計算上下包跡線(Envelopes),即置信區間。假定種群是隨機分布,則用隨機模型擬合一組點的坐標值,對每一t值,計算L(t);同樣用隨機模型再擬合新一組點坐標,分別計算不同尺度t的L(t)。這一過程重復進行直到達到事先確定的次數,L(t)的最大值和最小值分別為上下包跡線的坐標值。
種間格局分析就是2個種的點格局分析,前文中的特定種的格局可以將K(t)寫為K11(t),第2個中的寫為K22(t),2個種個體在尺度t內的數目,以下式求K12(t)和H12(t):
其中,n1,n2分別為物種1和物種2的個體數(點數);i,j代表種群1和種群2的個體;其他符號含義同前文。當H12(t)等于0時,說明2個種群在t尺度下無關聯;H12(t)大于0,表明它們正關聯;H12(t)小于0則為負關聯。所有數據分析過程都通過Programita軟件完成。
2 結果與分析
2.1 短花針茅空間分布格局
短花針茅的點格局在不同放牧干擾下存在著差異(圖1),在CK處理中,0—250cm尺度上短花針茅存在著兩種分布格局,即隨機分布和聚集分布。在0—25、70、110—250cm尺度上都是隨機分布,在30—65、75—105cm尺度上是集群分布,聚集強度在55cm尺度上最大。SA1處理下在0—25cm之間是隨機分布,25—250cm尺度上是集群分布,并且隨著尺度的增加聚集強度波浪式增加。SA2處理0—60cm尺度上是隨機分布,65—250cm尺度上是集群分布,且聚集強度逐漸增加。SA3處理下,隨著尺度的增加分布方式為隨機分布-集群分布-隨機分布方式,在0—95cm尺度上均為隨機分布,200—220cm尺度上為集群分布,225—250cm為隨機分布。SA4處理下,短花針茅先為隨機分布,然后變為集群分布,0—55cm為隨機分布,60—250cm均為集群分布,聚集強度在165cm處達到一個小高峰,然后聚集強度稍微下降,然后再次增加,250cm尺度上聚集強度最大。SA5處理下種群空間格局也是先為隨機分布然后變為集群分布,在0—85cm尺度上為隨機分布,在90—250cm尺度上均為集群分布。
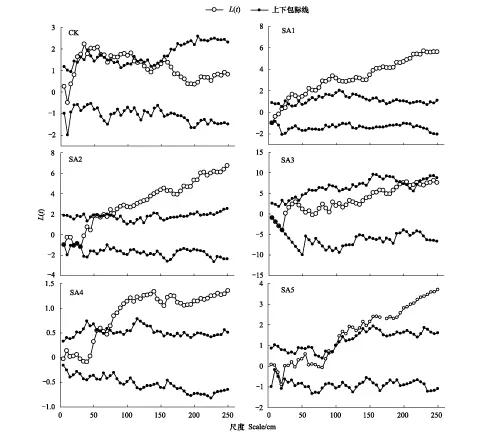
圖1 不同放牧強度季節調控處理下短花針茅點格局分析Fig.1 Point pattern analysis of Stipa breviflora in seasonal regulation under different grazing intensities
2.2 無芒隱子草空間分布格局
無芒隱子草的空間點圖及格局分布見圖2,從表中可以看出無芒隱子草隨著尺度的增加分布模式大部分表現為均勻分布-隨機分布-集群分布模式。CK處理下無芒隱子草在0—50cm尺度上表現為均勻分布,55—135cm尺度上表現為隨機分布,140—225尺度上為集群分布,且聚集強度隨著尺度的增加在逐漸的增加,說明無芒隱子草在較小尺度上為均勻分布,在中等尺度上為隨機分布,在較大尺度上為集群分布。SA1處理下10—70cm為均勻分布,75—235cm為隨機分布,240—245cm為集群分布。SA2處理下0—10cm為隨機分布,15—40cm為均勻分布,45—250cm為集群分布,聚集強度在235cm處達到最大。SA3處理下10—85cm尺度上為隨機分布,90—250cm尺度上均為隨機分布。SA4處理下,10—75cm為均勻分布,80—160cm尺度上為隨機分布,165—185、195—205、215—225cm均為集群分布,190cm和210cm為集群分布。SA5處理下,10—60cm尺度上為均勻分布,65—135cm為隨機分布,135—225cm為集群分布。
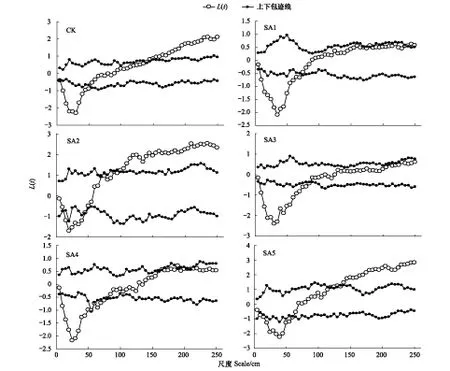
圖2 放牧強度調控處理下無芒隱子草點格局分析Fig.2 Point pattern analysis of Cleistogenes songorica in seasonal regulation under different grazing intensities
2.3 堿韭空間分布格局
堿韭的空間點圖及格局分布圖見圖3,CK處理下,0—70cm尺度上表現為均勻分布,170—250cm尺度上為隨機分布。SA1處理下0—95cm尺度下為均勻分布,100—250cm尺度上為隨機分布。SA2處理下,表現為隨機分布-均勻分布-隨機分布;0—35cm處理下為隨機分布,40—100cm為均勻分布,105—250cm為隨機分布。SA3處理下隨著尺度的增大,為隨機分布-均勻分布-隨機分布-集群分布模式,0—5cm分布方式為隨機分布,20—45cm為均勻分布,50—205cm尺度上為隨機分布,210—250cm尺度上為集群分布。SA4處理下,0—40cm尺度下,堿韭分布方式為隨機分布,45—115cm尺度上表現為均勻分布,120—225cm尺度上表現為隨機分布。SA4處理下,隨著尺度的增加分布方式變的較為復雜,在均勻分布和隨機分布之間來回變動,0—5cm尺度上為隨機分布,10—55cm為均勻分布,60cm為隨機分布,65cm為均勻分布,70—85cm為隨機分布,90—140cm為均勻分布,145—250cm為隨機分布。
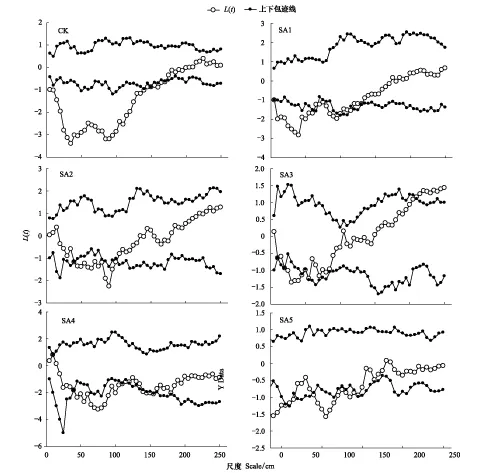
圖3 不同放牧強度季節調控處理下堿韭點格局分析Fig.3 Point pattern analysis of Allium polyrhizum in seasonal regulation under different grazing intensities
2.4 主要植物種群關聯性
主要植物種群關聯性見圖4,從圖中可以看出短花針茅和無芒隱子草在不同放牧處理下關聯性有一定差別,一般小尺度上表現為無關聯或者負關聯,在中等尺度上無關聯或者正關聯。短花針茅和堿韭在小尺度上為正關聯或者無關聯,在較大尺度上表現為負關聯。無芒隱子草和堿韭在較小尺度上為負關聯,較大尺度上無關聯。短花針茅和無芒隱子草在SA3和SA5處理下0—250cm尺度內均無關聯,CK處理下0—20cm尺度上短花針茅和無芒隱子草負關聯,25—70cm尺度上正關聯,75—115cm尺度上無關聯,120cm尺度上負關聯,125—250cm尺度上正關聯;SA1處理下5—10cm尺度上無關聯,15cm尺度上正關聯,20—250cm尺度上正關聯;SA2處理下5—30cm無關聯,40—250cm尺度上正關聯;SA4處理下5cm尺度上無關聯,10—40cm尺度上負關聯,45—250cm尺度上無關聯。短花針茅和堿韭、無芒隱子草和堿韭的關聯性見圖4。
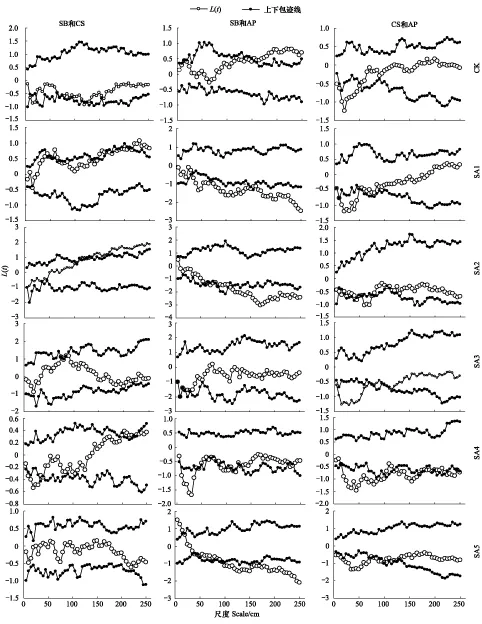
圖4 放牧強度季節調控處理下主要植物種之間關聯性研究(SB:短花針茅;CS:無芒隱子草;AP:堿韭)Fig.4 Spatial association of main plant populations in seasonal regulation under different grazing intensities(SB:Stipa breviflora;CS:Cleistogenes songorica;AP:Allium polyrhizum)
3 討論與結論
3.1 短花針茅、無芒隱子草和堿韭的點格局
種群空間格局是個很復雜的過程,是各種因素共同相互作用的結果[15]。格局的形成過程、種群所處的演替階段、種群在群落中的數量以及人為干擾是造成不同格局類型的重要因素。春季休牧+夏季重牧+秋季輕牧處理下0—25cm之間是隨機分布,春季休牧+夏季適牧+秋季重牧處理0—60cm尺度上是隨機分布,春季重牧+夏季重牧+秋季輕牧處理下0—55cm為隨機分布,全年輕牧處理下在0—85cm尺度上為隨機分布,全年重牧處理下0—195cm尺度上均為隨機分布。這說明隨著放牧強度的增大或者是持續放牧的影響,使得隨機分布尺度在逐漸的增大,物種在更大的尺度上才可能為集群分布,表現為斑塊化現象。大尺度下的斑塊往往表現為相對穩定,而小尺度下的斑塊則相對多變[16- 17],本研究中春季休牧+夏季重牧+秋季輕牧處理下斑塊尺度較大,因此比較穩定,這與吳艷玲的研究結果一致[18]。物種在自然界分布的斑塊性是一種很普遍的現象[19],斑塊在物種分布格局和生態過程中起著物質交換和能量流動載體的作用[20]。造成這種現象的原因可能是物種在重度干擾下種群的斑塊破裂逐漸向小型化發展。趙成章對阿爾泰針茅種群格局的研究表明,斑塊破碎是阿爾泰針茅種群拓殖和衰退的決定因素[9]。通過對短花針茅點格局的分析可以得出家畜的過度采食、踐踏導致種群分布從集群分布向隨機分布過渡,草地禁牧使得植物種內為爭奪各種資源而互相排斥,從而導致植物種群從集群分布逐漸向隨機分布過渡;家畜的過度采食與踐踏雖然能夠降低種群為爭奪資源而互相競爭,但是物理作用使得群落逐漸從大株叢向小株叢過渡。
無芒隱子草和短花針茅的格局分布有很大差別,無芒隱子草分布類型隨著尺度的增大表現為隨機分布-均勻分布-隨機分布-集群分布或者隨機分布,全年重度放牧區均勻分布的尺度最大,其它處理區均勻分布的尺度都小于全年重度放牧處理。Wang對羊草再生性的研究表明,家畜過度的采食及踐踏使羊草形成斑塊的能力減弱[21]。這些研究都說明了放牧通過影響群落物種的生長狀況進而影響種群的生長格局。植物種群空間格局轉化的轉折點分析是尺度分析的目的之一[22]。本實驗得出不同的放牧處理下從集群分布向隨機分布、隨機分布向均勻分布的尺度轉化的臨界點是不同的,從圖中大致可以看出春季重牧+夏季重牧+秋季輕牧和全年重度放牧處理下無芒隱子草在較大尺度上才表現為集群分布,尺度轉化的臨界點在放牧的影響下有增大的趨勢。
堿韭的點格局分析表明,堿韭的分布格局主要表現為隨機分布-均勻分布-隨機分布-隨機分布或者集群分布。全年重度放牧下堿韭在較大尺度上表現為集群分布,而其它放牧處理下250cm尺度以內沒有出現集群分布方式。這可能是因為放牧降低了短花針茅和無芒隱子草對堿韭的抑制作用,導致堿韭向著集群分布方式發展。陳寶瑞對不同干擾下羊草種群點格局分析表明,放牧增加了對羊草的采食及踐踏,從而減弱了羊草形成斑塊化的能力[8],這與本研究的結果不一致。在重度放牧下堿韭為了適應放牧干擾逐漸向集群分布方向發展,提高種群的穩定性抵御過度的干擾。
3.2 空間關聯性分析
空間關聯分析結果是種群空間關系的表象,正負關聯特征可能是物種間相互作用結果,也可能是物種生境趨同或趨異的表現[24]。研究表明試驗區優勢植物種短花針茅、無芒隱子草和堿韭在小尺度上基本為無關聯或負關聯,隨著尺度的增加主要植物種之間無關聯或正關聯。短花針茅和堿韭在春季重牧+夏季重牧+秋季重牧處理下0—250cm尺度上均無關聯性,春季休牧+夏季重牧+秋季適牧和春季休牧+夏季適牧+秋季重牧處理下短花針茅和堿韭隨著尺度的增大表現為正關聯,說明這兩種放牧方式使草地植物種之間相互促進,種間排布在較大尺度上向斑塊化方向發展;過度的放牧釋放了空間資源,植物種之間對資源的競爭減弱,導致種群隨機分布。無芒隱子草和堿韭在較小尺度上表現為負關聯,隨著尺度的增大種間關聯性消失,50—100cm尺度是負關聯向無關聯的臨界點,在放牧的干擾下無芒隱子草和堿韭負關聯的尺度有增大的趨勢。本研究與向春玲的研究結論不太一致[25],這也許是因為不同種群之間的差異或者放牧干擾下種群的關聯性發生變化。因此影響物種之間關聯性的機理需要更進一步研究,以期能夠揭示種間關聯性的影響機理。
綜上所述,表明短花針茅在禁牧和過度放牧下集群分布的尺度在增加,禁牧和過度放牧兩種草地利用方式下短花針茅的格局具有趨同的趨勢;春季重牧+夏季重牧+秋季輕牧和全年重度放牧處理下無芒隱子草在較大尺度上才表現為集群分布,尺度轉化的臨界點在放牧的影響下有增大的趨勢;重度放牧下堿韭為了適應放牧干擾逐漸向集群分布方向發展,集群分布的尺度在減小以提高種群的穩定性抵御過度的干擾。在不同的放牧干擾強度下植物種群具有明顯的響應策略,大致表現為物種的群居性在增強,物種集群分布的尺度在減小以提高種群的穩定性從而抵御過度的干擾。春季休牧+夏季重牧+秋季輕牧處理下種群斑塊化的尺度較大,有利于群落的穩定,因此荒漠草原采用這種利用方法較為合理。
[1] 張金屯. 數量生態學. 北京: 科學出版社, 2004.
[2] Watt A S. Pattern and process in the plant community. Journal of Ecology, 1947, 35(1/2): 1- 22.
[3] Purves D W, Law R. Fine-scale spatial structure in a grassland community: quantifying the plant′s-eye view. Journal of Ecology, 2002, 90(1): 121- 129.
[4] Wiegand T, Gunatilleke S, Gunatilleke N, Okuda T. Analyzing the spatial structure of a Sri Lankan tree species with multiple scales of clustering. Ecology, 2007, 88(12): 3088- 3102.
[5] Brown J R, Stuth J W. How herbivory affects grazing tolerant and sensitive grasses in a central Texas grassland: integrating plant response across hierarchical levels. Oikos, 1993, 67(2): 291- 297.
[6] Dieckmann U, Herben T, Law R. Spatio-temporal processes in plant communities//Lepenies W, ed. Yearbook1995/1996, Institute for Advanced Study Berlin. Berlin, Germany: Nicolaische Verlagsbuchhandlung, 1997: 296- 326.
[7] 劉振國, 李鎮清. 不同放牧強度下冷蒿種群小尺度空間格局. 生態學報, 2004, 24(2): 227- 234.
[8] 陳寶瑞, 楊桂霞, 張宏斌, 段慶偉, 辛曉平. 不同干擾類型下羊草種群的空間格局. 生態學報, 2010, 30(21): 5868- 5874.
[9] 趙成章, 任珩, 盛亞萍, 高福元, 石福習. 不同高寒退化草地阿爾泰針茅種群的小尺度點格局. 生態學報, 2011, 31(21): 6388- 6395.
[10] 李博. 中國北方草地退化及其防治對策. 中國農業科學, 1997, 30(6): 1- 9.
[11] 任繼周, 王欽, 牟新待, 胡自治, 符義坤, 孫吉雄. 草原生產流程及草原季節畜牧業. 中國農業科學, 1987, 11(2): 87- 92.
[12] 衛智軍, 韓國棟, 趙鋼, 李德新. 中國荒漠草原生態系統研究. 北京: 科學出版社, 2013.
[13] Ripley B D. Modelling spatial patterns. Journal of the Royal statistical Society. Series B (Methodological), 1977, 39(2): 172- 212.
[14] Diggle P J. Statistical Analysis of Spatial Point Patterns. New York: Academic Press, 1983.
[15] 楊君瓏, 王輝, 王彬, 孫棟元. 子午嶺油松林灌木層主要樹種的空間分布格局和種間關聯性研究. 西北植物學報, 2007, 27(4): 791- 796.
[16] 鄔建國. 生態學范式變遷綜論. 生態學報, 1996, 16(5): 449- 460.
[17] Levin S A. The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture. Ecology, 1992, 73(6): 1943- 1967.
[18] 吳艷玲, 陳立波, 衛智軍, 劉紅梅, 運向軍, 王穎杰, 展春芳. 不同放牧壓短花針茅荒漠草原群落植物種的空間異質特征. 干旱區資源與環境, 2012, 26(7): 110- 115.
[19] 張衛國, 黃文冰, 楊振宇. 草地微斑塊與草地退化關系的研究. 草業學報, 2003, 12(3): 44- 50.
[20] Gonzalez M, Ladet S, Deconchat M, Cabanettes A, Alard D, Balent G. Relative contribution of edge and interior zones to patch size effect on species richness: an example for woody plants. Forest Ecology and Management, 2010, 259(3): 266- 274.
[21] 王仁忠. 放牧影響下羊草種群生殖生態學的研究. 應用生態學報, 2000, 11(3): 399- 402.
[22] Peterson D L, Parker V T. Ecological Scale: Theory and Applications. New York: Columbia University Press, 1998.
[23] 王伯蓀. 植物種群學. 廣州: 中山大學出版社, 1989: 115- 120.
[24] 向春玲, 張金屯. 東靈山亞高山草甸優勢種的點格局分析. 草業科學, 2013, 30(3): 317- 321.
Point pattern and spatial association of primary plant populations in the seasonal regulation of grazing intensity in desert grassland
SUN Shixian1, WEI Zhijun2, WU Xinhong1,*, JIANG Chao1, GUO Libiao1
1InstituteofGrasslandResearch,ChinaAcademyofAgriculturalSciences,Huhhot010020,China2CollegeofEcologyandEnvironmentalScience,InnerMongoliaAgriculturalUniversity,Huhhot010018,China
The spatial distribution pattern of a population demonstrates a combination of the unique characteristics of the population, and the interrelationship between species and the environment. In the present study,Stipabreviflorawas used to examine the point pattern and spatial correlation of primary plant populations using Programita software, Ripley′s K function, and the Monte Carlo Method. The results showed that the scale of random distribution gradually increased with elevation of grazing intensity, or continuous grazing, and may be the primary explanation for the clustered distribution of populations at large scales. The clustered distribution scale ofS.brevifloraincreased in the “grazing prohibited” and “overgrazed conditions”. These two conditions were also associated with a convergence trend inS.breviflora. Increasing grazing intensity, or continuous grazing, gradually amplified the random clustered distribution scale ofS.brevifloraand may be the primary explanation for this clustered distribution. On a larger scale,Cleistogenessongoricashowed a clustered distribution under “heavy grazing in spring + heavy grazing in summer + light grazing in autumn”, and “heavy annual grazing”. The critical point of the above scale conversion showed an increasing trend with grazing.Alliumpolyrhizumshowed a trend of clustered distribution in response to grazing disturbance. The clustered distribution of this species was reduced to improve the population stability and avoid interference. Plant populations had specific response strategies under different grazing intensities, including enhancement of species sociality, and reduction of the clustered distribution, to improve population stability and avoid excessive disturbance.S.breviflora,C.songorica, andA.polyrhizumhad either no correlation or a negative correlation at the small scale. At larger scales, primary plant species had either no correlation or a positive correlation. At a scale of 0—250 cm,S.brevifloraandA.polyrhizumshowed no correlation in conditions of “heavy grazing in spring + heavy grazing in summer + light grazing in autumn”. However, at larger scales,S.brevifloraandA.polyrhizumwere positively correlated in the conditions of “zero grazing in spring + heavy grazing in summer + light grazing in autumn”, and “zero grazing in spring + light grazing in summer + heavy grazing in autumn”. Conditions of “spring grazing”, “summer grazing”, and “fall grazing” produced relatively lager-scale population patches that benefited the stability of the population. Thus, these methods are more suitable for the desert grassland.
desert grassland; grazing intensities; seasonal regulation of grazing intensity; point pattern of plant population; spatial association
國家重點基礎研究發展計劃(973計劃)(2014CB1388022);科技部“十三五”農村領域國家科技計劃項目(2012BAD13B00);中國農業科學院科技創新工程(CAASASTIP-IGR)
2015- 09- 17;
2016- 05- 10
10.5846/stxb201509171915
*通訊作者Corresponding author.E-mail: wxh1@vip.sina.com
孫世賢,衛智軍,吳新宏,姜超,郭利彪.不同放牧強度季節調控下荒漠草原主要植物種群點格局及空間關聯性.生態學報,2016,36(23):7570- 7579.
Sun S X, Wei Z J, Wu X H, Jiang C, Guo L B.Point pattern and spatial association of primary plant populations in the seasonal regulation of grazing intensity in desert grassland.Acta Ecologica Sinica,2016,36(23):7570- 7579.
猜你喜歡
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:30
當代陜西(2021年17期)2021-11-06 03:21:36
原道(2020年2期)2020-12-21 05:47:06
當代陜西(2019年15期)2019-09-02 01:52:00
中國非營利評論(2018年2期)2018-06-18 10:48:50
學苑創造·A版(2018年11期)2018-02-01 06:29:20
自動化學報(2017年1期)2017-03-11 17:31:17
讀者(2017年5期)2017-02-15 18:04:18
西藏科技(2016年5期)2016-09-26 12:16:39
振動工程學報(2015年1期)2015-03-01 01:15:42
