Pseudomonas putida細胞對微囊藻毒素-LR的脅迫響應
2016-04-16 07:12:32鄧庭進葉錦韶劉芷辰華南理工大學環境與能源學院工業聚集區污染控制與生態修復教育部重點實驗室廣東廣州50006暨南大學環境學院廣東廣州506暨南大學化學系廣東廣州506
中國環境科學 2016年2期
鄧庭進,尹 華*,葉錦韶,彭 輝,劉芷辰(.華南理工大學環境與能源學院,工業聚集區污染控制與生態修復教育部重點實驗室,廣東 廣州 50006;.暨南大學環境學院,廣東 廣州 506;.暨南大學化學系,廣東 廣州 506)
?
Pseudomonas putida細胞對微囊藻毒素-LR的脅迫響應
鄧庭進1,尹 華1*,葉錦韶2,彭 輝3,劉芷辰1(1.華南理工大學環境與能源學院,工業聚集區污染控制與生態修復教育部重點實驗室,廣東 廣州 510006;2.暨南大學環境學院,廣東 廣州 510632;3.暨南大學化學系,廣東 廣州 510632)
摘要:將1.0g/L的微囊藻毒素-LR(MC-LR)降解菌惡臭假單胞菌(Pseudomonas putida)置于含不同濃度MC-LR的體系中,研究了體系中菌體細胞完整性和生物量的變化,考察了MC-LR對細胞的氧化脅迫以及抗氧化酶的響應.結果表明,MC-LR能夠增大P. putida細胞質膜通透性,造成膜損傷,導致胞內物質外流,使細胞完整性遭到破壞;同時,MC-LR能夠引起P. putida細胞的氧化脅迫,隨著毒素暴露時間的延長,活性氧自由基(ROS)和膜脂過氧化產物丙二醛(MDA)含量顯著升高,具有明顯的劑量效應.超氧化物歧化酶(SOD)活性在MC-LR的誘導下有一個先升后降的過程,表現為對低濃度污染物的主動響應,而高濃度(2.5mg/L)MC-LR作用5d后,ROS積累到相當高水平,對細胞代謝功能造成破壞,使SOD活性下降,并加速細胞的死亡,P. putida生物量與對照相比,下降了將近50%.
關鍵詞:微囊藻毒素-LR;惡臭假單胞菌;氧化脅迫;生物量;毒性
* 責任作者, 教授, huayin@scut.edu.cn
微囊藻毒素(MCs)是富營養化污染的次生代謝產物,其結構為環狀七肽,具有很強的生物毒性.研究表明,世界上25%~70%的藍藻水華能夠產生藻毒素[1],對環境危害極大.目前,有關MCs的毒性研究大多集中在動物、高等植物上[2-4],而其對微生物的作用機制研究還很有限,大量細菌、真菌、浮游植物等微生物與藻毒素共存于富營養化水體中,兩者相互接觸,勢必存在相互影響[5-6].楊翠云等[7-8]研究發現MCs能夠促進溶菌酶對大腸桿菌和枯草芽孢桿菌的細胞膜滲透性,增加細胞內可溶性糖和可溶性蛋白的外滲.尹黎燕等[9]研究顯示,MC-LR可對煙草BY-2細胞造成氧化脅迫,并激發細胞內抗氧化系統的響應.另一方面,產毒藻類普遍存在于富營養化水環境中,能夠為微生物的附著生長及生物修復提供條件,然而,MCs的產生與藍藻水華暴發具有同步性,二者呈極顯著正相關[10],MCs對微生物的作用是否會影響到水華污染水域的修復作用,是亟待解決的科學問題.
當前,MCs的微生物降解去除是科學研究的熱點,而關于MCs對菌體的毒性作用研究較少.袁媛等[11]研究發現,MC-LR能抑制降解菌的生長,導致MC-LR的降解率下降.Ding等[12]利用純微囊藻毒素作用于菌體中,發現藻毒素能夠引起菌體細胞的氧化脅迫,并破壞菌體線粒體和電子傳遞鏈.本文選取我國藍藻水華中常見且毒性最強的MC-LR作為MCs代表物,利用本實驗室前期分離、篩選得到的對MC-LR有降解作用的惡臭假單胞菌純菌株(Pseudomonas putida)為研究對象,分析MC-LR對菌體細胞的毒性脅迫影響,并考察細胞抗氧化系統的響應機制,以期為進一步揭示MCs對菌體的作用機制和生理毒性影響,探明藻毒素對生態環境中藻菌之間的作用影響提供一定的理論依據.
1 材料與方法
1.1 實驗試劑與材料
實驗菌種:惡臭假單胞菌(Pseudomonas putida),由本課題組從廣州市某發生水華的人工湖底泥中篩選馴化得到,該菌對MC-LR有良好的降解效果[13].
MC-LR儲備液由實驗室制備,其提取和純化方法參照文獻[14].
營養培養基:牛肉膏 3g,蛋白胨 10g,NaCl 5g,蒸餾水 1000mL,pH為7.4~7.6.
無機鹽基礎培養基(MSM):K2HPO40.02g, NaH2PO40.1g, MgSO4·7H2O 0.05g, NH4NO30.02g, CaCl20.05g,水1000mL.
絕對計數管(TruCOUNT Tubes)購于Becton–Dickinson (SanJose, CA, USA);2',7'-二氯二氫熒光素二乙酯(DCFH-DA)購于Sigma-Aldrich公司.超氧化物歧化酶(SOD)測試試劑盒、丙二醛(MDA)測試試劑盒均購于南京建成生物研究所.
1.2 實驗設備
高速冷凍離心機(KDC-160HR,安徽中科中佳科學儀器有限公司);紫外分光光度計(UVmini-1240,日本島津公司);高效液相色譜(CBM-20A,日本島津公司);流式細胞儀(FACSDiva,美國BD公司);超聲破壁儀(SCIENTZ-ⅡD,寧波新芝科技有限公司).
1.3 P. putida的毒素處理
將活化后的P. putida投加到MC-LR質量濃度分別為0,0.5,2.5mg/L的滅菌無機鹽基礎培養基中,使體系最終為20mL,菌體質量濃度為1.0g/L,于30℃、130r/min搖床中恒溫振蕩培養.以不加MC-LR的MSM體系作為空白對照.
1.4 MC-LR對P. putida細胞完整性的影響
分別取毒素處理0,12,24,48,72,96,120h后的樣品,測定其在260nm處的吸光度值,以表征細胞完整性的變化.每組設置3個平行,進行2次重復實驗.
1.5 MC-LR對P. putida細胞的氧化脅迫
1.5.1 MC-LR作用下P. putida細胞內活性氧自由基(ROS)的測定 分別取毒素處理第0,1,2,3, 4,5d樣品,離心收集菌體,用10mg/L溶菌酶處理后,用雙蒸水清洗2~3遍,用PBS重懸,加入200μL DCFH-DA染液,混勻,37℃避光孵育20min;加入500μL PBS洗滌,置于1000r/min下離心5min,棄上清液,加入200μL PBS,混勻,用流式細胞儀檢測樣品中生成的DCF熒光強度[15].
1.5.2 MC-LR作用下P. putida細胞內丙二醛(MDA)的測定 毒素處理后于0,24,48,72,96, 120h進行取樣,離心收集菌體,用PBS清洗后重懸,置于0℃冰-水混合水浴中,超聲破壁.超聲破壁儀功率為:450W,工作3s,間隙3s,破壁150次.取混合懸液利用丙二醛測試試劑盒測定MDA含量.
1.6 MC-LR作用下P. putida超氧化物歧化酶S0D的響應變化
1.6.1 P. putida細胞酶液的制備 將P.putida菌體培養液離心過濾,收集菌體,用0.9%的生理鹽水清洗3次.洗滌后的菌體收集于離心管中,加入生理鹽水10mL,置于0℃的冰-水混合水浴中,超聲破壁.將破碎后混合懸液離心15min (8000r/ min,4℃),上清液即為所需酶液.參照考馬斯亮藍G-250法測定制備酶液中可溶性蛋白含量,用牛血清蛋白(BSA)作為標準蛋白[16].
1.6.2 超氧化物歧化酶(SOD)活性測定 分別取毒素處理后第0,1,2,3,4,5d樣品,離心收集菌體,根據1.6.1進行酶液的制備,并利用試劑盒測定SOD酶活性.
1.7 MC-LR對P. putida細胞生物量的影響
取毒素處理后第0,2,4,6d樣品,離心收集底部菌體細胞,用PBS洗滌后重懸,稀釋2000倍后, 取300μL加入絕對計數管中,渦旋震蕩使樣品與管中的內參物熒光微球混勻后上機檢測.
每根絕對計數管中含有已知數量的內參物——熒光微球,通過流式細胞儀可以直接檢測到儀器進樣中P. putida細胞的數目和熒光微球的數目,得出數據后,通過公式(1)換算即可直接得出每個樣品中P. putida的細菌數目[17]:

1.8 統計分析
試驗所獲數據利用SPSS 17.0和One-Way ANOVA法進行相關性分析和差異顯著性分析.
2 結果與討論
2.1 MC-LR對P. putida細胞完整性的影響
正常狀態下的細胞,細胞質膜是完整閉合的,胞內物質被封閉起來,只通過主動運輸或離子交換等方式與外界進行物質交換.當細胞質膜被損壞時,細胞質膜通透性會增大,某些胞內大分子物質會流出,這些胞內物質中如DNA、RNA等在波長260nm的發射光照射下能夠被檢出,因此,可以把測得的吸光度值變化情況作為間接判斷細胞質膜完整性是否受到損壞的依據[18].
由圖1可知,在未添加MC-LR的對照體系中,隨著培養時間的延長,吸光度值略微上升,這可能是由于培養到后期,部分細胞逐漸衰老,少量細胞出現死亡破裂現象,胞內物質外流,從而導致所測吸光度值升高,反映出P. putida的完整性受到一定程度的破壞.而對于經不同濃度MC-LR處理的實驗組,測得的吸光度值增加的現象較對照出現得更早,且隨著MC-LR濃度的增加,吸光度值上升幅度更大(P<0.05),這表明MC-LR導致了胞內物質的大量外流,破壞了P. putida細胞的完整性,分析原因,一方面,可能是因為MC-LR能作用于P. putida的細胞質膜,使細胞質膜通透性增大[14],從而使胞內物質更易流出;另一方面,MC-LR可能致使細胞發生極化現象,使細胞遭受損傷,加速細胞的破裂和死亡,最終使得胞內物質大量流出,細胞的完整性遭到破壞.
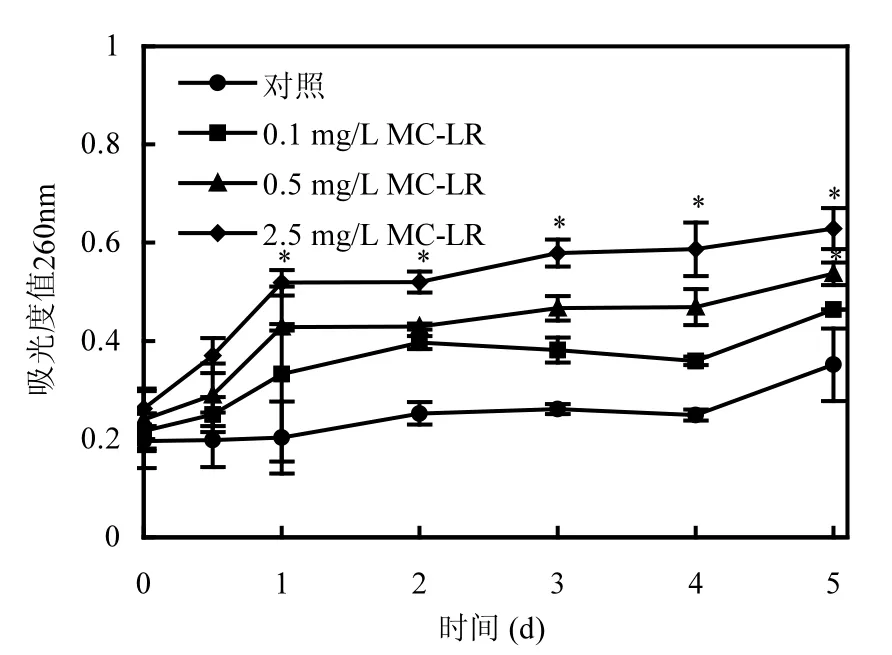
圖1 MC-LR對P. putida細胞完整性的影響Fig.1 Effect of MC-LR on the cell integrity of P. putida *表示與對照組有顯著性差異(P<0.05),下同
2.2 MC-LR對P. putida細胞的氧化脅迫
活性氧自由基ROS(Reactive Oxygen Species)是一類包括超氧陰離子自由基、羥基自由基、過氧化氫等在內的高反應性分子[19].在正常情況下,ROS為細胞提供正常的生理環境,而細胞內過量和長時間的ROS作用能破壞胞內蛋白,引發脂質過氧化作用,并因此形成脂質過氧化產物[20].丙二醛(MDA)是膜脂過氧化最重要的產物之一,它的產生能加劇膜的損傷,因此,可以通過ROS和MDA的含量了解膜脂過氧化的程度,以間接反映P. putida細胞及膜系統受氧化脅迫的損傷程度[21].
圖2和圖3分別為培養體系中不同MC-LR濃度下P. putida細胞內ROS和MDA隨時間的變化情況.由圖中可以看出,MC-LR可引起細胞ROS和MDA含量增多,差異具有顯著性(P<0.05),且表現出一定的劑量效應,即MC-LR濃度越高,ROS和MDA增加越明顯.
圖2中,將P. putida投加至空白對照體系中,由于細胞進入新的培養環境而產生氧化脅迫,而菌體細胞通過自身的調節機制,很快適應了新的環境.因此,表現為ROS含量略微增加,但維持在較低水平.
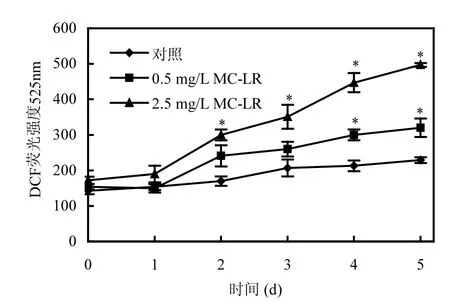
圖2 MC-LR對P. putida細胞活性氧自由基的影響Fig.2 Effect of MC-LR on ROS of P. putida
經MC-LR處理后的實驗組細胞從第2d 起,ROS含量急劇上升,顯著高于對照組,并在整個培養階段都維持在較高水平,表明P. putida細胞受到了新的氧化脅迫,而這種脅迫正是由MC-LR所引起的.低濃度的MC-LR(0.5mg/L)毒性處于一個較低水平,對ROS的激發作用較小,對菌體的毒害作用也有限,但會隨著暴露時間的增加而出現ROS的累積,最終使ROS含量逐漸上升.當把加入體系中的MC-LR濃度提高到2.5mg/L時,菌體ROS水平經過第1d的自動調整期之后,迅速呈現直線增長.Xiong等[22]發現,高濃度的MC-LR毒害性增大,能導致細胞膜系統變性,進而導致細胞的損傷,迅速破壞細胞內的自我防御機制.因此,高濃度MC-LR使P. putida處于不利環境,激發并積累大量活性氧自由基,毒素暴露5d后,ROS含量超過空白對照將近2倍多,對細胞的氧化脅迫程度大大加深.
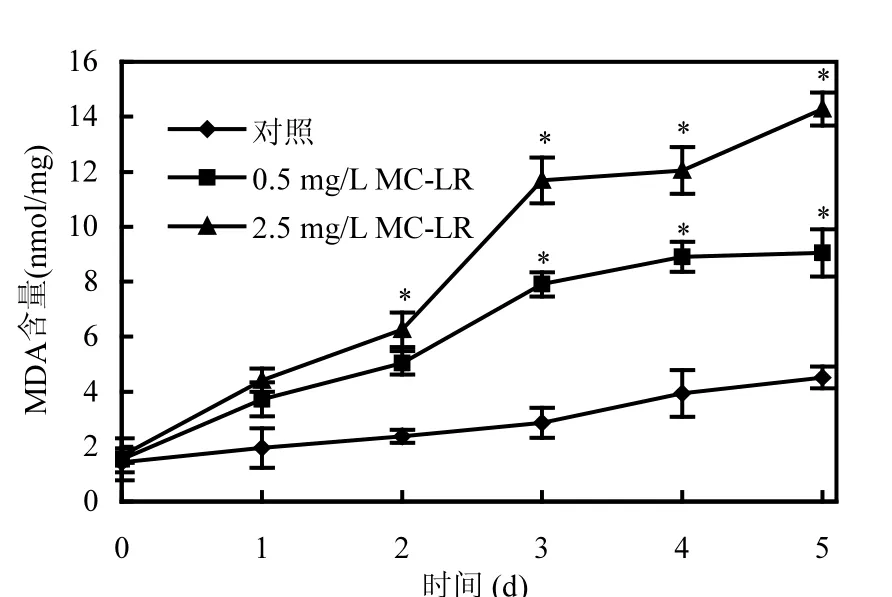
圖3 MC-LR作用下P. putida細胞丙二醛的變化Fig.3 Effect of MC-LR on MDA content of P. putida
膜脂過氧化是藻毒素誘導氧化損傷的另一個表征[23].由圖3可知,對照組的膜脂過氧化程度(以MDA含量表示)在整個實驗階段保持較低水平,這與ROS的形成相似.利用SPSS 17.0統計軟件對毒素處理組MDA含量的變化和ROS含量變化情況作相關性分析,得出其相關性為0.959,在0.01水平上顯著正相關,說明P. putida細胞膜脂過氧化的程度與ROS的變化也存在一致性,均受到MC-LR劑量效應的影響與時間效應的累積作用.這是因為高濃度的MC-LR具有很強的生物毒性,能通過產生活性氧來誘導氧化應激,使得細胞受到氧化脅迫[24],影響細胞的活性,細胞功能遭到破壞,導致活性氧自由基ROS的升高和積累,并發生膜脂過氧化作用,造成細胞膜電位的去極化損傷和通透性的變化,從而使膜脂過氧化產物MDA的含量增加,這些結果與Ding等[12]的研究是一致的.
2.3 MC-LR對P. putida細胞抗氧化性酶活的影響
超氧化物歧化酶(SOD)是普遍存在于生物中的抗膜脂質過氧化的保護性酶.研究表明,高濃度的污染物能夠導致細胞內活性氧自由基的過度積累,并引起氧化脅迫,造成細胞質膜系統脂質和蛋白的過氧化失活,并導致DNA等斷裂,最終使細胞發生死亡[19].而SOD能快速將超氧陰離子自由基氧化還原為H2O2和O2,H2O2則進一步經抗氧化酶分解為H2O和O2,從而限制自由基的過量積累,消除活性氧自由基對細胞的傷害.

圖4 MC-LR對P. putida細胞超氧化物歧化酶的影響Fig.4 Effect of MC-LR on SOD activity of P. putida
如圖4所示,在空白對照體系中,SOD酶活性處于比較平穩的狀態,在0~2d有略微上升,但波動不大,3d以后略有下降,說明在空白體系中SOD酶沒有受到顯著誘導或抑制作用,變化不明顯.而在添加MC-LR的體系中,P. putida受到氧化脅迫,由2.2可知,細胞內氧自由基濃度上升,誘導抗氧化酶系統做出反應,因此,在0~2d SOD酶活性也會隨著上升,以清除胞內自由基,減輕細胞損傷.而隨著藻毒素暴露時間的延長,從第2d開始,SOD酶活性出現明顯的下降(P<0.05).由于細胞內氧自由基濃度超過一定的范圍,抗氧化體系酶活不能及時清除,一方面,SOD與MC-LR相結合,參與了MC-LR的解毒和ROS的清除[25],消耗大量的SOD;另一方面,過量的自由基會導致嚴重的氧化脅迫,破壞細胞,損傷細胞功能,使得抗氧化體系酶活性降低,SOD供應不足[9],最終導致SOD整體活性下降,且MC-LR濃度越高,下降越明顯.
根據本實驗結果及結合前人研究,可以得出,菌體抗氧化酶活可能受到誘導而升高,也有可能受到抑制,對于一定濃度范圍內的MC-LR作用,P. putida細胞表現出主動響應,而當污染物濃度過高時,表現為對毒素的被動脅迫.
2.4 MC-LR對P. putida細胞生物量的影響
本實驗采用流式細胞術(FCM)和絕對計數管(TruCount tube)相結合,對培養體系中的P. putida生物量進行定量檢測.該方法是利用特殊熒光信號對懸液中的單細胞或生物粒子進行標記,再通過流式細胞儀對熒光信號進行檢測和分析,從而實現高速、逐一的細胞定量分析和分選[26],比傳統的細胞計數方法速度更快,精度更高,且更準確無誤.
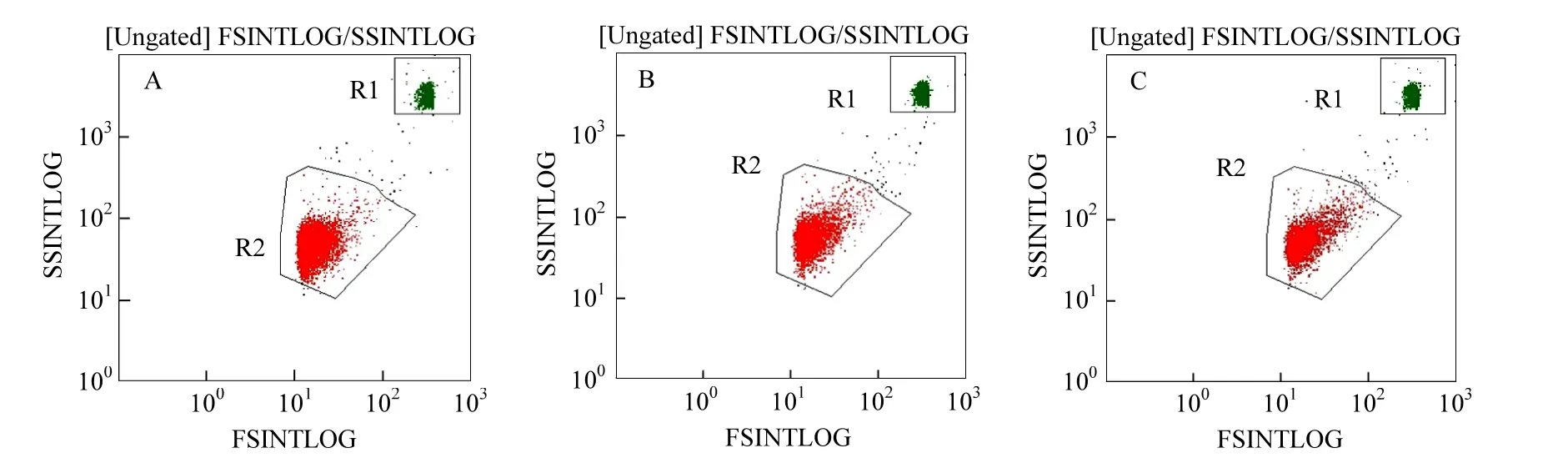
圖5 絕對計數流式檢測(0d)Fig.5 Flow cytometry diagram of P. putida A.對照; B. 0.5mg/L MC-LR; C. 2.5mg/L MC-LR
圖5為流式細胞儀熒光檢測結果圖,圖中熒光信號由兩部分構成,其中R1部分為絕對計數管中微球的熒光信號,R2部分為檢測樣品中P. putida的熒光信號.由于絕對計數管中微球的數量為已知量,因此,通過樣品信號和微球信號的對比,即可計算出樣品中P. putida的數量,再通過樣品的稀釋倍數換算,便可得出培養體系中P. putida的準確生物量.
MC-LR對體系中P. putida細胞生物量的影響如圖6所示,在培養過程中P. putida細胞的生物量變化情況可以反映出菌體對MC-LR作用的響應.由圖可知,體系中P. putida細胞的數目均呈下降趨勢.在空白體系中,當P. putida 從營養培養基轉移到無機鹽培養基中,由于環境的變化及外界營養的突然匱乏,導致菌體短時間內適應較差,從而發生菌體失活或死亡破裂等情況,使得細胞數目略有減少;而隨著培養時間延長,越來越多細胞進入衰老期,死亡破裂的細胞逐漸增多,導致體系中P. putida的生物量進一步減少.
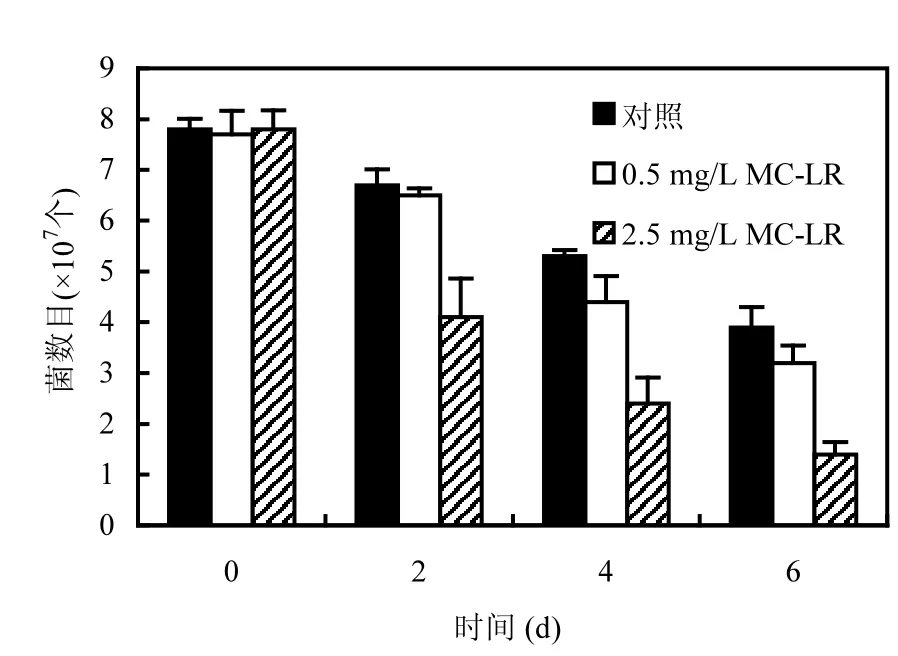
圖6 MC-LR對體系中 P. putida 細胞數目的影響Fig.6 Effect of MC-LR on biomass of P. putida
而MC-LR的加入,使得上述情況變得更加明顯,且加入的MC-LR濃度不同,對P. putida生物量的影響作用也不同,且差異具有顯著性(P< 0.05).當向體系中加入0.5mg/L的MC-LR時,與空白對照相比,菌體生物量有略微的下降,這是由于污染物對P. putida細胞具有脅迫作用,加速了細胞的衰亡,使菌體細胞提前進入衰亡期,但是當污染物濃度處于較低水平時,其影響作用也較小;而當加入體系中的MC-LR濃度提高到2.5mg/L 時,污染物的毒性脅迫作用進一步加強,表現出明顯的劑量效應.P. putida在高濃度MC-LR的毒害下,活性降低并發生細胞凋亡[12],細胞膜通透性逐漸增大,細胞完整性遭到破壞,且隨著暴露時間的延長,污染物作用的時間效應加劇了細菌的死亡和消融,從而使得P. putida生物量迅速減少.
3 結論
3.1 MC-LR作用于P. putida的細胞質膜,使細胞質膜通透性增大,細胞遭受損傷,細胞的完整性遭到破壞.
3.2 MC-LR可引起細胞ROS含量的增多和積累,使P. putida遭受氧化脅迫,繼而發生膜脂過氧化作用,使膜脂過氧化產物MDA的含量增加.P. putida細胞膜脂過氧化的程度與ROS的變化也存在一致性,均受到MC-LR劑量效應的影響與時間效應的累積作用.
3.3 對于低濃度(0.5mg/L)的MC-LR作用,P. putida細胞表現出主動響應,細胞抗氧化酶SOD誘導升高;而當污染物濃度過高時(2.5mg/L),其毒性脅迫作用增強,SOD酶活性受到抑制而降低,且導致P. putida生物量的減少.
參考文獻:
[1] Chen X F, Chuai X M, Yang L Y. Status quo, historical evolution and causes of eutrophication in lakes in typical lake regions of China [J]. Journal of Ecology and Rural Environment, 2014,30(4): 438-443.
[2] 黃文敏,邢 偉,李敦海,等.微囊藻毒素-RR對煙草細胞活力及營養生理的影響 [J]. 中國環境科學, 2007,27(3):382-386.
[3] 王偉琴,金永堂,吳 斌,等.水源水中微囊藻毒素的遺傳毒性與健康風險評價 [J]. 中國環境科學, 2010,30(4):468-476.
[4] Yin L Y, Huang J Q, Huang W M, et al. Microcystin-RR-induced accumulation of reactive oxygen species and alteration of antioxidant systems in tobacco BY-2cells [J]. Toxicon, 2005, 46(5):507-512.
[5] 徐慧敏,閆 海,馬 松,等.鞘氨醇單胞菌USTB-05對微囊藻毒素的生物降解 [J]. 中國環境科學, 2014,34(5):1316-1321.
[6] 張 睿,王廣軍,李志斐,等.枯草芽孢桿菌對銅綠微囊藻抑制效果的研究 [J]. 中國環境科學, 2015,35(6):1814-1821.
[7] 楊翠云,李敦海,劉永定.微囊藻毒素對典型微生物生長及生理生化特性的影響 [J]. 水生生物學報, 2008,32(6):818-823.
[8] 楊翠云,夏傳海,周世偉.微囊藻毒素-RR對大腸桿菌和枯草桿菌的滲透效應 [J]. 科學通報, 2010,55(11):1002-1006.
[9] 尹黎燕,黃家權,沈 強,等.煙草懸浮細胞抗氧化系統對微囊藻毒素-RR的響應 [J]. 中國環境科學, 2005,25(5):576-580.
[10] 鮑忠祥.滇池浮游植物和微囊藻毒素的時空分布及細菌去除毒素研究 [D]. 昆明,云南大學, 2012.
[11] 袁 媛,吳 涓,李玉成,等.活性炭纖維固定化菌對微囊藻毒素MC-LR的去除研究 [J]. 中國環境科學, 2014,34(2):403-409.
[12] Ding W X, Shen H M, Ong C N. Calpain activation after mitochondrial permeability transition in microcystin-induced cell death in rat hepatocytes [J]. Biochemical and Biophysical Research Communications, 2002,291(2):321-331.
[13] 肖巧巧,尹 華,葉錦韶,等.一株微囊藻毒素-LR降解菌的降解特性 [J]. 環境化學, 2014,33(9):1-7.
[14] 鄧庭進,葉錦韶,彭 輝,等.微囊藻毒素-LR對惡臭假單胞菌細胞活性和表面特性的影響 [J]. 環境科學, 2015,36(1):252-258.
[15] He Y Y, H?der D P. UV-B-induced formation of reactive oxygen species and oxidative damage of the cyanobacterium Anabaena sp.: protective effects of ascorbic acid and N-acetyl-L-cysteine [J]. Journal of Photochemistry and Photobiology B: Biology, 2002,66(2):115-124.
[16] Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding [J]. Analytical Biochemistry, 1976,72(1): 248-254.
[17] Khan M M T, Pyle B H, Camper A K. Specific and rapid enumeration of viable but nonculturable and viable-culturable gram-negative bacteria by using flow cytometry [J]. Applied and environmental microbiology, 2010,76(15):5088-5096.
[18] Rurián-Henares J A, Morales F J. Antimicrobial activity of melanoidins against Escherichia coli is mediated by a membranedamage mechanism [J]. Journal of Agricultural and Food Chemistry, 2008,56(7):2357-2362.
[19] Schalk I J, Hannauer M, Braud A. New roles for bacterial siderophores in metal transport and tolerance [J]. Environmental Microbiology, 2011,13(11):2844-2854.
[20] 王 燕,陳永剛,葛鄭增,等.TBT對大鼠肝臟ROS、抗氧化酶和解毒系統酶的影響 [J]. 中國環境科學, 2005,25(4):428-431.
[21] Codreanu S G, Liebler D C. Novel approaches to identify protein adducts produced by lipid peroxidation [J]. Free Radical Research, 2015(0):1-7.
[22] Xiong Q, Xie P, Li H, et al. Acute effects of microcystins exposure on the transcription of antioxidant enzyme genes in three organs (liver, kidney, and testis) of male Wistar rats [J]. Journal of Biochemical and Molecular Toxicology, 2010,24(6): 361-367.
[23] ?egura B, ?traser A, Filipi? M. Genotoxicity and potential carcinogenicity of cyanobacterial toxins–a review [J]. Mutation Research/Reviews in Mutation Research, 2011,727(1):16-41.
[24] Pinho G L L, da Rosa C M, Maciel F E, et al. Antioxidant responses and oxidative stress after microcystin exposure in the hepatopancreas of an estuarine crab species [J]. Ecotoxicology and Environmental Safety, 2005,61(3):353-360.
[25] Pflugmacher S, Wiegand C, Beattie K A, et al. Uptake, effects, and metabolism of cyanobacterial toxins in the emergent reed plant Phragmites australis (cav.) trin. ex steud [J]. Environmental Toxicology and Chemistry, 2001,20(4):846-852.
[26] Wang Y, Hammes F, De Roy K, et al. Past, present and future applications of flow cytometry in aquatic microbiology [J]. Trends in biotechnology, 2010,28(8):416-424.
Response of Pseudomonas putida cells to MC-LR stress.
DENG Ting-jin1, YIN Hua1*, YE Jin-shao2, PENG Hui3, LIU Zhi-chen1(1.Key Laboratory on Pollution Control and Ecosystem Restoration in Industry Clusters, Ministry of Education College of Environment and Energy, South China University of Technology, Guangzhou 510006, China;2.College of Environment, Jinan University, Guangzhou 510632, China;3.Department of Chemistry, Jinan University, Guangzhou 510632, China). China Environment Science, 2016,36(2):603~609
Abstract:The cell integrity and biomass changes of Pseudomonas putida, a MC-LR degrading bacterium, were studied by inoculating 1.0g/L bacterium into systems with different concentrations of MC-LR. The oxidative stress of MC-LR on bacterial cell and the responses of antioxidase were also investigated. The results showed that membrane permeability of P. putida increased under the influence of MC-LR, causing membrane damage, which resulted in the outflow of intracellular substances and the destruction of cell integrity. Also, MC-LR could induce the oxidative stress on the cells of P. putida. With prolonged exposure to MC-LR, reactive oxygen species (ROS) and malondialdehyde (MDA), a product of membrane lipid peroxidation, evidently accumulated in the system, and had obvious dose-effect relationship. Under the effect of MC-LR, the activity of superoxide dismutase (SOD) increased first and then declined, exhibiting an active response to MC-LR of low level. However, after contacting higher concentrations (2.5mg/L) of MC-LR for 5d, ROS accumulation was so high as to cause damage to the metabolism of cells. As a result, SOD activity was suppressed and cells suffered mass mortality, and the biomass decreased by 50% compared with the control.
Key words:microcystin-LR;Pseudomonas putida;oxidative stress;biomass;toxicity
作者簡介:鄧庭進(1989-),男,湖南郴州人,華南理工大學碩士研究生,主要研究方向為水污染的微生物修復.
基金項目:國家自然科學基金委-廣東省聯合基金重點項目(U0933002);廣東省自然科學基金重點項目(S2013020012808)
收稿日期:2015-07-02
中圖分類號:X172
文獻標識碼:A
文章編號:1000-6923(2016)02-0603-07
