金烏賊幼體捕食行為及捕食能力的研究*
2016-05-12 03:01:30雷舒涵張秀梅張沛東覃樂(lè)政
雷舒涵, 張秀梅, 張沛東, 覃樂(lè)政
(中國(guó)海洋大學(xué)海水養(yǎng)殖教育部重點(diǎn)實(shí)驗(yàn)室,山東 青島 266003)
?
金烏賊幼體捕食行為及捕食能力的研究*
雷舒涵, 張秀梅**, 張沛東, 覃樂(lè)政
(中國(guó)海洋大學(xué)海水養(yǎng)殖教育部重點(diǎn)實(shí)驗(yàn)室,山東 青島 266003)
摘要:實(shí)驗(yàn)室條件下,觀(guān)察并記錄胴背長(zhǎng)為8.2~22.9 mm的金烏賊(Sepia esculenta)幼體的捕食行為,通過(guò)分析不同的捕食行為指標(biāo)研究金烏賊幼體的捕食能力。研究發(fā)現(xiàn),金烏賊幼體對(duì)糠蝦的捕食行為分為攻擊和攝食兩部分,其中攻擊行為分為發(fā)現(xiàn)、定位和捕捉3個(gè)階段。金烏賊幼體捕食成功率較高,胴背長(zhǎng)大于10 mm的幼體捕食成功率大于80%。捕食發(fā)生次數(shù)(FT)、攻擊距離(AD)、最大攻擊距離(MAD)和總捕食時(shí)間(TFT)與胴背長(zhǎng)(DML)均呈二次多項(xiàng)式關(guān)系,分別為:FT=0.107DML2-0.170DML+3.157,AD=-0.030DML2-1.641DML+3.150,MAD=-0.067DML2+3.226DML-2.682,TFT=-7.947DML2+404.9DML-1079;總捕食量(TFA)與胴背長(zhǎng)呈指數(shù)關(guān)系(TFA=5.254e(0.133 DML));食餌操縱時(shí)間(HT)與胴背長(zhǎng)呈冪函數(shù)關(guān)系(HT=1587DML(-0.91))。捕食發(fā)生次數(shù)、攻擊距離、最大攻擊距離、總捕食時(shí)間和總捕食量隨生長(zhǎng)而增大,食餌操縱時(shí)間隨生長(zhǎng)而減少,表明金烏賊幼體隨生長(zhǎng)捕食能力迅速增強(qiáng)。本研究結(jié)果豐富了頭足類(lèi)行為生態(tài)學(xué)理論知識(shí)。
關(guān)鍵詞:金烏賊; 幼體; 捕食行為; 捕食能力
引用格式:雷舒涵,張秀梅,張沛東,等. 金烏賊幼體捕食行為及捕食能力的研究[J].中國(guó)海洋大學(xué)學(xué)報(bào)(自然科學(xué)版),2016, 46(4): 37-42.
LEI Shu-Han, ZHANG Xiu-Mei, ZHANG Pei-Dong, et al. Studies on the feeding behavior and ability of juvenileSepiaesculenta(Cephalopoda: Sepiida) [J].Periodical of Ocean University of China, 2016, 46(4):37-42.
金烏賊(Sepiaesculenta)屬軟體動(dòng)物門(mén)(Mollusca)頭足綱(Cephalopoda)鞘亞綱(Coleoidea)烏賊目(Sepiida)烏賊科(Sepiidae)烏賊屬(Sepia),是中國(guó)沿海重要經(jīng)濟(jì)頭足類(lèi)。主要分布在俄羅斯遠(yuǎn)東海海域,日本本州、四國(guó)、九州海域,朝鮮半島西海岸、南海岸,中國(guó)沿海及菲律賓群島海域[1-2]。金烏賊生命周期短,世代更替快,生長(zhǎng)迅速,肉質(zhì)鮮美,蛋白質(zhì)含量高,是一種具有較高經(jīng)濟(jì)價(jià)值的優(yōu)良種類(lèi)[3]。
由于金烏賊特殊的生態(tài)習(xí)性和較高的經(jīng)濟(jì)價(jià)值,國(guó)內(nèi)外學(xué)者在生理、行為、資源增殖等方面展開(kāi)了相關(guān)研究[4-7]。關(guān)于行為生態(tài)學(xué)方面,國(guó)外學(xué)者研究較多,主要集中于繁殖行為[8-9],而對(duì)捕食行為的研究?jī)H見(jiàn)郝振林[3]關(guān)于不同底質(zhì)、光照度、視角對(duì)金烏賊捕食行為影響的報(bào)道。金烏賊幼體階段是一個(gè)快速生長(zhǎng)和高死亡率的發(fā)育階段[10],幼體的捕食能力對(duì)其早期生長(zhǎng)和存活具有重要影響[11]。本文在實(shí)驗(yàn)室條件下,觀(guān)察了胴背長(zhǎng)8.2~22.9mm金烏賊幼體的捕食行為,探討了金烏賊幼體早期發(fā)育階段的捕食能力,以期為頭足類(lèi)的早期培育和人工養(yǎng)殖提供科學(xué)依據(jù)。
1材料與方法
1.1 實(shí)驗(yàn)材料
實(shí)驗(yàn)用金烏賊受精卵于2010年6月取自山東黃島金烏賊繁殖基地。受精卵運(yùn)至中國(guó)海洋大學(xué)魚(yú)類(lèi)行為生態(tài)實(shí)驗(yàn)室后置于4個(gè)35cm×25cm×30cm玻璃水槽(養(yǎng)殖水體15 L)中孵化,孵化水溫22~24℃,鹽度32±0.5,pH7.8~8.6,連續(xù)充氧,日換水量為1/2。孵出后,幼體培育條件與孵化條件相同。0~5日齡幼體投喂鹵蟲(chóng)(Artemiaurmiana)無(wú)節(jié)幼體和活體糠蝦(Neomysisawatschensis),5~30日齡投喂活體糠蝦,從30日齡開(kāi)始投喂全長(zhǎng)為3~4cm的日本沼蝦(Macrobrachiumnipponense)和麥穗魚(yú)(Pseudorasboraparva),每天投餌2次(09:00和18:00),每次投餌充足。
1.2 實(shí)驗(yàn)方法
實(shí)驗(yàn)對(duì)象為胴背長(zhǎng)(Dorsal mantle length,DML)8.2~22.9mm的20尾(10~50日齡)金烏賊幼體。為保證餌料的一致性和可比性,捕食實(shí)驗(yàn)中統(tǒng)一使用糠蝦。實(shí)驗(yàn)前將實(shí)驗(yàn)個(gè)體饑餓1 d,隨機(jī)選取一尾幼體移入60cm×30cm×40cm玻璃水槽(養(yǎng)殖水體50L)中,水槽中糠蝦密度為50ind/L,糠蝦全長(zhǎng)為(8.4 ± 1.1)mm。幼體的捕食行為用掛在水槽正上方的Sony HDR-XR550E攝像機(jī)記錄。攝影從幼體第一次捕食開(kāi)始,隨捕食糠蝦數(shù)量的增加,幼體對(duì)糠蝦的食餌操縱時(shí)間會(huì)逐漸延長(zhǎng),當(dāng)食餌操縱時(shí)間延長(zhǎng)至10min時(shí)(幼體進(jìn)入飽食狀態(tài)),攝影停止。利用該段視頻資料,統(tǒng)計(jì)分析每只幼體在總捕食時(shí)間段內(nèi)捕食發(fā)生次數(shù)、捕食成功率、攻擊距離、最大攻擊距離、總捕食量、食餌操縱時(shí)間和總捕食時(shí)間等指標(biāo)。捕食實(shí)驗(yàn)結(jié)束后立即稱(chēng)取幼體體質(zhì)量(Body weight,BW)。
1.3 數(shù)據(jù)分析
利用MoveTr2D和ImageJ軟件分析捕食錄像,計(jì)算各捕食能力指標(biāo),如下:
總捕食時(shí)間(Total feeding time,TFT):視頻資料中,幼體烏賊開(kāi)始捕食至捕食最后一只糠蝦所用時(shí)間。
捕食發(fā)生次數(shù)(Feeding times,F(xiàn)T):總捕食時(shí)間段內(nèi)幼體完成捕食反應(yīng)的總次數(shù)。
捕食成功率(Successful feeding rate,SFR):總捕食時(shí)間段內(nèi)幼體成功捕食糠蝦的次數(shù)與捕食發(fā)生次數(shù)的比例。
攻擊距離(Attacking distance,AD):幼體發(fā)現(xiàn)餌料并定位后,捕食糠蝦的平均距離。
最大攻擊距離(Maximum of attacking distance,MAD):幼體發(fā)現(xiàn)餌料并定位后,捕食糠蝦的最大距離。
總捕食量(Total feeding amount,TFA):幼體烏賊捕食糠蝦的總質(zhì)量(濕重),計(jì)量方法采用估算法,由于同一批糠蝦的個(gè)體質(zhì)量相差不大,用精密電子天平(Sartorius)稱(chēng)取100尾糠蝦的體質(zhì)量后,估算出每一尾糠蝦的平均體質(zhì)量,再乘以幼體烏賊捕食糠蝦的總個(gè)數(shù)即為總捕食量。
食餌操縱時(shí)間(Handling time,HT):幼體襲擊、捕捉并撕咬餌料生物的耗時(shí)以及開(kāi)始重新搜尋餌料的停留時(shí)間的和。
最優(yōu)食物供給量(Optimal food supply,OFS):總捕食量占烏賊幼體體質(zhì)量的百分比。
2結(jié)果
2.1 捕食行為的觀(guān)察
金烏賊對(duì)糠蝦的捕食行為分為攻擊和攝食兩個(gè)部分,而攻擊又主要分為3個(gè)階段:發(fā)現(xiàn)、定位和捕捉(見(jiàn)圖1)。
發(fā)現(xiàn)(Attention,見(jiàn)圖1a,1b):當(dāng)糠蝦進(jìn)入到幼體的視野內(nèi),即刻發(fā)生一系列的行為反應(yīng):幼體眼睛隨著糠蝦位置發(fā)生轉(zhuǎn)動(dòng),腕、頭、胴體的顏色加深;頭和胴體轉(zhuǎn)向糠蝦方向,第一對(duì)腕豎直向上延伸,左右擺動(dòng)。發(fā)現(xiàn)的過(guò)程通常時(shí)間很短,一般小于1 s。
定位(Positioning,見(jiàn)圖1b):當(dāng)烏賊面向糠蝦后,或原地不動(dòng),或按著食餌—身體軸線(xiàn)的方向前進(jìn)或者后退。當(dāng)距糠蝦較遠(yuǎn)時(shí),烏賊會(huì)前進(jìn)以便接近糠蝦;當(dāng)距糠蝦較近時(shí),烏賊則會(huì)向后游泳,與糠蝦保持一定距離。這個(gè)距離被稱(chēng)為攻擊距離(Attacking distance),攻擊距離并不恒定。
捕捉(Seizure,見(jiàn)圖1c):當(dāng)烏賊調(diào)整好攻擊距離后,第一對(duì)腕對(duì)著食物的方向部分展開(kāi)并持續(xù)幾秒鐘,隨后持續(xù)左右擺動(dòng)。腕、頭部、胴體表面顏色變化迅速,似黑色波浪閃動(dòng),此時(shí),鰭波動(dòng)的速率加快,猶如懸浮在水中。隨后烏賊突然躍起迅速伸出觸腕捕食糠蝦,從觸腕伸出到捕到糠蝦用時(shí)僅為30 ms。捕到食物后,幼體胴體呈現(xiàn)斑塊狀花紋。
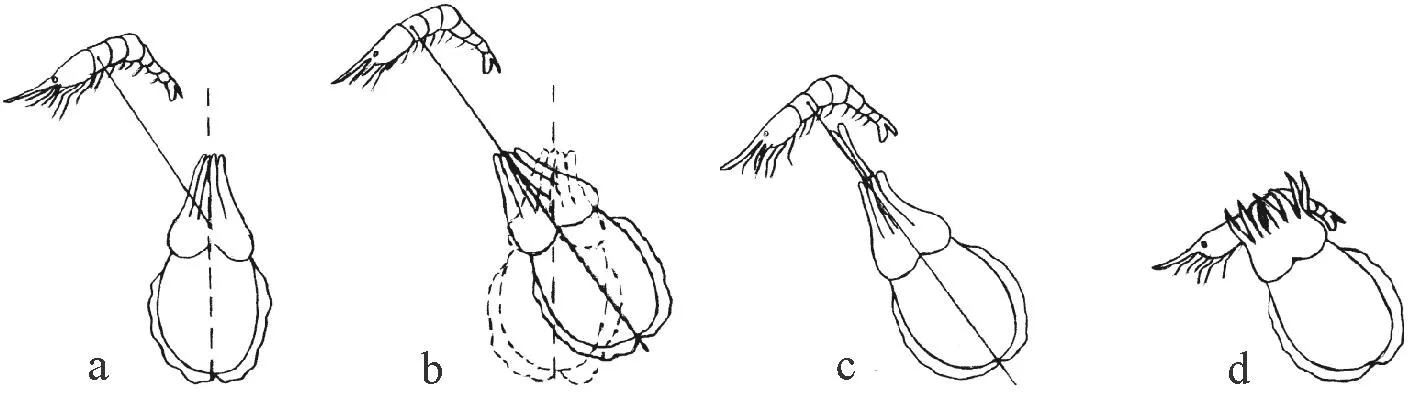
(a: 糠蝦進(jìn)入烏賊視野范圍; b: 頭和身體轉(zhuǎn)向糠蝦的方向; c: 烏賊伸出觸腕捕食糠蝦; d; 烏賊用喙撕咬,并食掉全部糠蝦。a: The shrimp swum into cuttlefish’s field of vision; b: The head-body axis turned to the shrimp; c: Tentacles ejected quickly to prey on the shrimp; d: The cuttlefish ate the whole shrimp by the beaks.)
圖1金烏賊捕食行為示意圖
Fig.1Sketch of the feeding behavior of golden cuttlefishSepiaesculenta
攝食(Eating,見(jiàn)圖1d):當(dāng)金烏賊捕到食物后,觸腕收回到觸腕腔中,其他腕迅速抱住食物,用喙撕咬,并食掉全部糠蝦,水槽中未見(jiàn)頭胸甲殘余。
2.2 捕食能力
2.2.1 捕食發(fā)生次數(shù)及捕食成功率分析錄像得出,DML為8.2mm的金烏賊幼體在觀(guān)測(cè)時(shí)間內(nèi)共發(fā)生捕食行為9次,而DML為21.8mm的幼體發(fā)生捕食行為57次(見(jiàn)圖2)。胴背長(zhǎng)和捕食發(fā)生次數(shù)的關(guān)系可以用二次多項(xiàng)式函數(shù)表示:FT=0.107DML2-0.170DML+3.157(R2= 0.834)。DML為8.8mm的金烏賊幼體捕食成功率最低,為58.3%,而DML>10mm的幼體捕食成功率較高,均大于80%(見(jiàn)圖3)。
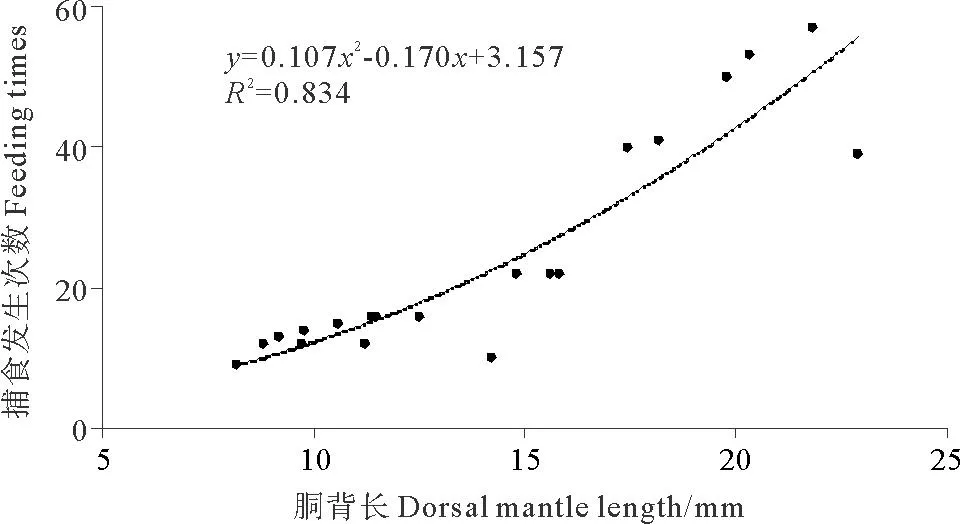
圖2 金烏賊幼體捕食發(fā)生次數(shù)與胴背長(zhǎng)的關(guān)系
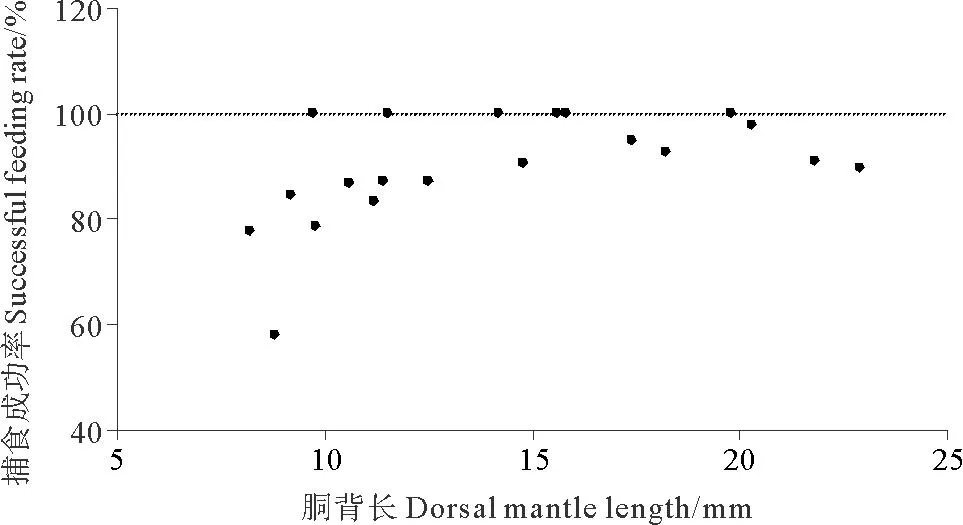
圖3 金烏賊幼體的捕食成功率
2.2.2 攻擊距離金烏賊幼體的攻擊距離隨著胴背長(zhǎng)的生長(zhǎng)而發(fā)生變化,攻擊距離為胴背長(zhǎng)的1~2倍,且胴背長(zhǎng)和攻擊距離的關(guān)系可以用二次多項(xiàng)式函數(shù)表示:AD=-0.030DML2-1.641DML+3.150(R2=0.865)(見(jiàn)圖4)。
金烏賊幼體的最大攻擊距離為胴背長(zhǎng)的1.5~3倍(見(jiàn)圖4)。胴背長(zhǎng)和最大攻擊距離的關(guān)系可以用二次多項(xiàng)式函數(shù)表示:MAD=-0.067DML2+3.226DML-2.682(R2=0.879)。
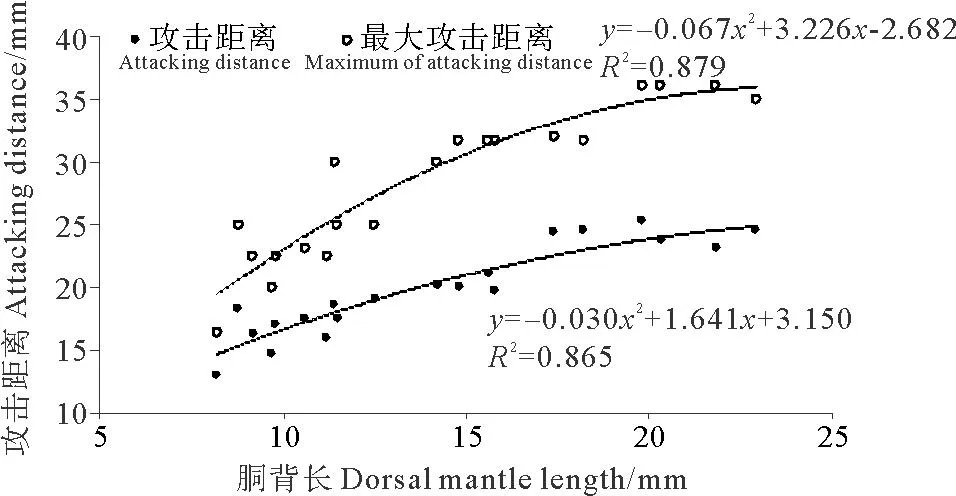
圖4 金烏賊幼體攻擊距離與胴背長(zhǎng)的關(guān)系
2.2.3 總捕食量在一次捕食全過(guò)程中,DML為8.2mm的金烏賊幼體捕食糠蝦的總濕重為13.27mg,而DML為21.8mm的幼體捕食糠蝦的總濕重為98.54mg(見(jiàn)圖5)。胴背長(zhǎng)和總捕食量的關(guān)系可以用指數(shù)函數(shù)表示:TFA=5.254e0.133 DML(R2=0.861)。
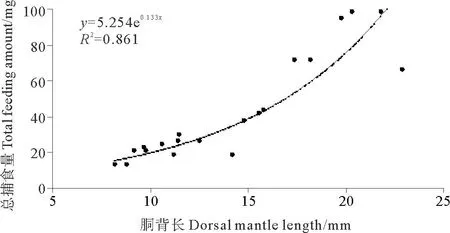
圖5 金烏賊幼體總捕食量與胴背長(zhǎng)的關(guān)系
金烏賊幼體總捕食量與體質(zhì)量的關(guān)系如圖6所示,其關(guān)系可以用多項(xiàng)式函數(shù)表示:TFA=-0.0085BW2+0.0705BW+0.0019(R2=0.972)。
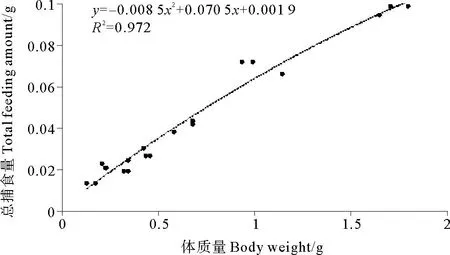
圖6 金烏賊幼體總捕食量與體質(zhì)量的關(guān)系
2.2.4 食餌操縱時(shí)間及總捕食時(shí)間金烏賊幼體食餌操縱時(shí)間隨著胴背長(zhǎng)的增大有減小的趨勢(shì)(見(jiàn)圖7), DML為8.2mm幼體的食餌操縱時(shí)間最長(zhǎng),為265s,而DML為21.8mm個(gè)體的食餌操縱時(shí)間最短,為75s。胴背長(zhǎng)和食餌操縱時(shí)間的關(guān)系可以用冪函數(shù)表示:HT=1587DML-0.91(R2=0.629)。
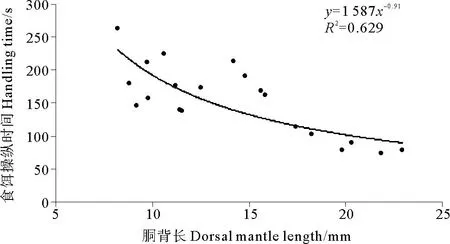
圖7 金烏賊幼體食餌操縱時(shí)間與胴背長(zhǎng)的關(guān)系
金烏賊幼體總捕食時(shí)間隨著胴背長(zhǎng)的增大有增加的趨勢(shì)(見(jiàn)圖8)。DML為9.2mm個(gè)體的總捕食時(shí)間最短,為1768s;捕食時(shí)間最長(zhǎng)(4712s)的個(gè)體胴背長(zhǎng)為20.3mm。胴背長(zhǎng)和總捕食時(shí)間的關(guān)系可以用二次多項(xiàng)式函數(shù)表示:TFT=-7.947DML2+404.9DML-1079(R2=0.569)。

圖8 金烏賊幼體總捕食時(shí)間與胴背長(zhǎng)的關(guān)系
2.2.5 最優(yōu)食物供給量金烏賊幼體最優(yōu)食物供給量隨著體質(zhì)量的增大有減小的趨勢(shì)(見(jiàn)圖9),體質(zhì)量為0.20g個(gè)體的最優(yōu)食物供給量最高,為金烏賊幼體體質(zhì)量的11.19%;體質(zhì)量為1.79g個(gè)體的最優(yōu)食物供給量最低,為金烏賊幼體體質(zhì)量的5.49%。最優(yōu)食物供給量和體質(zhì)量的關(guān)系可以用對(duì)數(shù)函數(shù)表示:OFS = -1.487ln (BW)+6.1108(R2=0.479)。
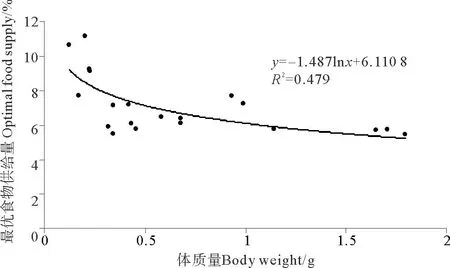
圖9 金烏賊幼體最優(yōu)食物供給量與體質(zhì)量的關(guān)系
3討論
烏賊類(lèi)的捕食行為較為相似,Messerger[12]和Kasugai[13]分別研究了歐洲烏賊(Sepiaofficinalis)和微鰭烏賊(Idiosepiusparadoxus)的捕食行為,發(fā)現(xiàn)攻擊行為均分為發(fā)現(xiàn)、定位和捕捉3個(gè)階段。但是Kasugai[13]發(fā)現(xiàn)微鰭烏賊的攝食行為比較特殊,它捕到食物后將口球(Buccal mass)伸入到甲殼動(dòng)物的外殼中,口球可以伸長(zhǎng)到與第一對(duì)腕相同的長(zhǎng)度,并可以伸向任意方向,只食用軟體組織,丟棄的外殼猶如剛剛蛻皮的完整外殼。而本實(shí)驗(yàn)中,由于糠蝦個(gè)體較小,金烏賊將糠蝦全部食掉,包括糠蝦最硬的頭胸甲。
本研究發(fā)現(xiàn)金烏賊幼體捕食成功率普遍較高,相同的結(jié)果在其它種類(lèi)的烏賊中亦有報(bào)道。Duval等[14]在研究歐洲烏賊的捕食行為中發(fā)現(xiàn),白色水槽中歐洲烏賊捕食成功率為87.7%,在灰色的水槽中更高達(dá)91%。烏賊捕食成功率較高的原因可能有:(1)金烏賊具有發(fā)達(dá)的視覺(jué)系統(tǒng)。個(gè)體在孵化初期,眼已經(jīng)發(fā)育完全,具有與成體相似的結(jié)構(gòu)[15],因此它們可以準(zhǔn)確地判斷餌料生物的方向和距離。(2)金烏賊具有發(fā)達(dá)的觸腕。觸腕是金烏賊的捕食器官,觸腕的頂端為觸腕穗,具有10余行吸盤(pán)[2],吸盤(pán)可以緊緊吸住食物,防止餌料生物逃脫。(3)金烏賊具有快速的觸腕發(fā)射能力,觸腕的發(fā)射極為迅速(約為22ms),這一點(diǎn)在Messerger[12]的實(shí)驗(yàn)中也被證實(shí):在水溫為25℃的條件下,歐洲烏賊觸腕從發(fā)射到捕捉到餌料生物的用時(shí)僅為15ms,如此之快的捕捉速度要?dú)w功于觸腕高度發(fā)達(dá)的橫紋肌的快速收縮[16]。
本研究發(fā)現(xiàn),金烏賊個(gè)體捕食發(fā)生次數(shù)、總捕食量和總捕食時(shí)間隨幼體生長(zhǎng)而增加,表明個(gè)體隨生長(zhǎng)需要更多的食物來(lái)滿(mǎn)足生長(zhǎng)能量的需求(見(jiàn)圖1、5、8)。目前研究最優(yōu)食物供給量的計(jì)算方法為食物占生物體重的百分比[17-18]。Koueta and Boucaud-Camou[18]在實(shí)驗(yàn)室條件下研究歐洲烏賊幼體生長(zhǎng)時(shí)發(fā)現(xiàn),烏賊隨生長(zhǎng)發(fā)育所需要的最優(yōu)食物供給量是隨生長(zhǎng)逐漸降低的。本研究中,將總捕食量換算為最優(yōu)食物供給量后也得出相同的結(jié)論,雖然烏賊幼體捕食餌料增多,但隨著生長(zhǎng)體重逐漸增大,其相對(duì)捕食量有降低的趨勢(shì)。本研究發(fā)現(xiàn),金烏賊個(gè)體的攻擊距離和最大攻擊距離隨著生長(zhǎng)發(fā)育有增大的趨勢(shì),這不僅增加了金烏賊的有效捕食范圍,還可以有效防止因距離過(guò)近可能會(huì)導(dǎo)致餌料生物對(duì)金烏賊產(chǎn)生躲避行為。然而Chen等[19]在研究乳光槍烏賊(Loligoopalescens)對(duì)橈足類(lèi)的捕食行為中發(fā)現(xiàn),與槍烏賊的發(fā)育階段和攻擊速度變化無(wú)明顯相關(guān),當(dāng)攻擊距離大于胴背長(zhǎng)時(shí),攻擊行為極少成功,攻擊距離并沒(méi)有隨生長(zhǎng)而增大,反而最小攻擊距離隨生長(zhǎng)減小。這可能是因?yàn)槿楣鈽尀踬\的捕食方式異于金烏賊。當(dāng)乳光槍烏賊發(fā)現(xiàn)食餌后,它會(huì)將頭和身體轉(zhuǎn)向食餌,張開(kāi)腕并前躍抱住食餌[19]。這樣的捕食方式導(dǎo)致遠(yuǎn)距離捕食成功率大大降低。而金烏賊捕食用觸腕吸住食物后回縮,同時(shí)游向食物,用口腕抱住進(jìn)行攝食,從而顯著擴(kuò)大了捕食范圍。
隨生長(zhǎng)發(fā)育,金烏賊幼體的食餌操縱時(shí)間明顯減少。食餌操縱時(shí)間為個(gè)體發(fā)現(xiàn)、捕捉和吞食餌料生物的時(shí)間以及開(kāi)始重新搜尋餌料的停留時(shí)間的總和。隨著幼體生長(zhǎng),視覺(jué)敏感度(Visual sensitivity)逐步提高[15],個(gè)體搜尋生物餌料的能力也逐步提升;與此同時(shí),幼體的消化能力也得到增強(qiáng)[20],因此,食餌操縱時(shí)間隨幼體生長(zhǎng)逐漸縮短。除此之外,在真鯛(Pagrosomusmajor)仔魚(yú)早期捕食能力的研究中也發(fā)現(xiàn)相同的結(jié)果[21],干重為50μg的仔魚(yú)在捕食褶皺臂尾輪蟲(chóng)(Brachionusplicatilis)時(shí)的食餌操縱時(shí)間為0.90h/ind,而干重為100μg仔魚(yú)的食餌操縱時(shí)間下降為0.04h/ind,并且餌料密度對(duì)食餌操縱時(shí)間的影響隨著仔魚(yú)個(gè)體的增大而逐漸減小。
綜上所述,金烏賊幼體對(duì)糠蝦的捕食行為分為攻擊和攝食兩個(gè)部分,其中攻擊行為又主要分為發(fā)現(xiàn)、定位和捕捉3個(gè)階段;金烏賊幼體捕食成功率較高,并且捕食能力隨生長(zhǎng)而增強(qiáng)。自然環(huán)境中的餌料密度較低,金烏賊幼體高效的捕食能力對(duì)其生長(zhǎng)和存活具有重要的生物學(xué)意義。研究確定的金烏賊胴背長(zhǎng)和總捕食量的關(guān)系曲線(xiàn),為其幼體人工培育過(guò)程中的適量投餌,節(jié)約養(yǎng)殖成本、避免水質(zhì)惡化和餌料浪費(fèi)提供了重要參考依據(jù)。
參考文獻(xiàn):
[1]齊鐘彥. 中國(guó)經(jīng)濟(jì)軟體動(dòng)物 [M]. 北京: 中國(guó)農(nóng)業(yè)出版社, 1998: 293-294.
Qi Zhongyan. Economic mollusca of China [M]. Beijing: China Agriculture Press, 1998: 293-294.
[2]鄭元甲, 陳雪忠, 程家驊, 等. 東海大陸架生物資源與環(huán)境 [M]. 上海: 上海科學(xué)技術(shù)出版社, 2003: 722-727.
Zheng Yuanjia, Chen Xuezhong, Cheng Jiahua, et al. The continental shelf, biological resources and the environment of east China sea [M]. Shanghai: Shanghai Science and Technology Press, 2003: 722-727.
[3]郝振林. 金烏賊的行為習(xí)性及標(biāo)志技術(shù)的研究 [D]. 青島: 中國(guó)海洋大學(xué), 2007.
Hao Zhenlin. Studies on the behavior and marking technology ofSepiaesculenta[D]. Qingdao: Ocean University of China, 2007.
[4]Watanuki N, Kawamura G, Kaneuchi S, et al. Role of vision in behavior, visual field, and visual acuity of cuttlefishSepiaesculenta[J]. Fisheries Science, 2000, 66: 417-423.
[5]郝振林, 張秀梅, 張沛東. 金烏賊的生物學(xué)特性及增殖技術(shù) [J]. 生態(tài)學(xué)雜志, 2007, 26(4): 601-606.
Hao Zhenlin, Zhang Xiumei, Zhang Peidong. Biological characteristics and multiplication techniques ofSepiaesculenta[J]. Chinese Journal of Ecology, 2007, 26(4): 601-606.
[6]雷舒涵, 吳常文, 高天翔, 等. 金烏賊和曼氏無(wú)針烏賊胚胎發(fā)育及其鹽度耐受能力的比較研究[J]. 中國(guó)水產(chǎn)科學(xué), 2011, 18(2): 350-359.
Lei Shuhan, Wu Changwen, Gao Tianxiang, et al. A comparative study ofSepiaesculentaandSepiellamaindronion embryonic development and ability of salinity tolerance [J]. Journal of Fishery Sciences of China, 2011, 18(2): 350-359.
[7]Lei S, Zhang X, Liu S, et al. Effects of temperature fluctuations on cuttlebone formation of cuttlefishSepiaesculenta[J]. Chinese Journal of Oceanology and Limnology, 2012, 30: 547-553.
[8]Fujita T, Hirayama I, Matsuoka T, et al. Spawning behavior and selection of spawning substrate by cuttlefishSepiaesculenta[J]. Nippon Suisan Gakkaishi, 1997, 63: 145-151.
[9]Wada T, Takegaki T, Mori T, et al. Sperm displacement behavior of the cuttlefishSepiaesculenta(Cephalopoda: Sepiidae) [J]. Journal of Ethology, 2005, 23: 85-92.
[10]陳四清, 劉長(zhǎng)琳, 莊志猛, 等. 餌料、鹽度對(duì)金烏賊幼體生長(zhǎng)的影響 [J]. 漁業(yè)現(xiàn)代化, 2008, 35(6): 23-25.
Chen Siqing, Liu Changlin, Zhuang Zhimeng, et el. Effect of feeds and water salinity on growth of golden sepia (Sepiaesculenta) [J]. Fishery Modernization, 2008, 35(6): 23-25.
[11]范帆, 尹飛, 彭士明, 等. 饑餓脅迫對(duì)曼氏無(wú)針烏賊幼體的影響 [J]. 生態(tài)學(xué)雜志, 2011, 30(10): 2262-2268.
Fan Fan, Yin Fei, Peng Shiming, et al. Impacts of starvation on biochemical indices and behaviors ofSepiellamaindronijuveniles [J]. Chinese Journal of Ecology, 2011, 30(10): 2262-2268.
[12]Messenger J B. The visual attack of the cuttlefish,Sepiaofficinalis[J]. Animal Behaviour, 1968, 16: 342-357.
[13]Kasugai T. Feeding behaviour of the Japanese pygmy cuttlefishIdiosepiusparadoxus(Cephalopoda: Idiosepiidae) in captivity: evidence for external digestion [J]. Journal of the Marine Biological Association of the United Kingdom, 2001, 81: 979-981.
[14]Duval P, Chichery M P, Chichery R. Prey capture by the cuttlefish (SepiaofficinalisL): An experimental study of two strategies [J]. Behavioural Processes, 1984, 9: 13-21.
[15]Hao Z L, Zhang X M, Kudo H, et al. Development of the retina in cuttlefishSepiaesculenta[J]. Journal of Shellfish Research, 2010, 29(2): 463-470.
[16]Kier W M. Squid cross-striated muscle: the evolution of a specialized muscle fiber type [J]. Bulletin of Marine Science, 1991, 49: 389-403.
[17]Koueta N, Boucaud-Camou E. Food intake and growth in reared early juvenile cuttlefishSepiaofficinalisL. (Mollusca Cephalopoda) [J]. Journal of Experimental Marine Biology and Ecology, 1999, 240: 93-109.
[18]Koueta N, Boucaud-Camou E. Basic growth relations in experimental rearing of early juvenile cuttlefishSepiaofficinalisL. (Mollusca: Cephalopoda) [J]. Journal of Experimental Marine Biology and Ecology, 2001, 265: 75-87.
[19]Chen D S, Van Dykhuizen G, Hodge J, et al. Ontogeny of copepod predation in juvenile squid (Loligoopalescens) [J]. Biological Bulletin, 1996, 190: 69-81.
[20]Moguel C, Mascaró M, Avila-Poveda O H, et al. Morphological, physiological and behavioural changes during post-hatching development ofOctopusmaya(Mollusca: Cephalopoda) with special focus on the digestive system [J]. Aquatic Biology, 2010, 9:35-48.
[21]殷名稱(chēng), 鮑寶龍, 蘇錦祥. 真鯛仔魚(yú)早期階段的攝食能力——發(fā)育反應(yīng)和功能反應(yīng) [J]. 海洋與湖沼, 1999, 30(6): 591-596.
Yin Mingcheng, Bao Baolong, Su Jinxiang. Feeding ability ofPagrosomusMajorlarvae during early edvelopment - ITS developmental and functional responses [J]. Oceanologia ET Limnologia Sinica, 1999, 30(6): 591-596.
責(zé)任編輯朱寶象
Studies on the Feeding Behavior and Ability of JuvenileSepiaesculenta(Cephalopoda: Sepiida)
LEI Shu-Han, ZHANG Xiu-Mei, ZHANG Pei-Dong, QIN Le-Zheng
(The Key Laboratory of Mariculture, Ministry of Education, Ocean University of China, Qingdao 266003, China)
Abstract:Cuttlefish is an active predator. Like all known coleoid cephalopods, it feeds on small crusta-ceans, fish, and shrimps. The feeding ability is vital for cuttlefish to grow and survive, especially for the newly hatched larvae. Aiming to gain the knowledge on the feeding ability of early developing cuttlefish, the feeding behavior of juvenile Sepia esculenta was observed under laboratory conditions. The mantle length of the experimental cuttlefish was 8.2~22.9 mm in length. The video camera was used to record the feeding behavior and the indicators of feeding ability. The feeding behavior on prawns consisted of two phases, namely attacking and eating. The sequence of attacking behavior comprised three phases, attention, positioning and seizure. The successful feeding rate of the juvenile cuttlefish with>10mm dorsal mantle length (DML) was relatively high, no less than 80%. The relationship between DML and feeding time (FT), attacking distance (AD), maximum of attacking distance (MAD), total feeding time (TFT) could be described with a quadratic model expressed as FT=0.107DML2-0.170DML+3.157; AD=-0.030DML2-1.641DML+3.150; MAD=-0.067DML2+3.226DML-2.682; and TFT=-7.947DML2+404.9DML-1079, respectively, while the relationship between DML and total feeding amount (TFA), handling time (HT) with an exponential model expressed as TFA=5.254e(0.133 DML) and HT=1587DML(-0.91), respectively. Feeding time, attacking distance, maximum of attacking distance, total feeding time and total feeding amount increased with the growth while handling time decreased with the growth. In conclusion, the successful feeding rate of juvenile cuttlefish was relatively high and the feeding ability rapidly increased with the growth. Because low prey density, high feeding rate and efficient feeding ability is extremely vital for cuttlefish to survive wild, the relationship between DML and TFA provides the evidence for suitable feeding. The results obtained in present study would enrich the knowledge of the ethology and ecology of cephalopods.
Key words:Sepia esculenta; juvenile; feeding behavior; feeding ability
DOI:10.16441/j.cnki.hdxb. 20130194
中圖法分類(lèi)號(hào):S917.4
文獻(xiàn)標(biāo)志碼:A
文章編號(hào):1672-5174(2016)04-037-06
作者簡(jiǎn)介:雷舒涵(1987-), 女, 博士生, 研究方向:金烏賊資源增殖學(xué)研究。 E-mail: shuhanlei@163.com**通訊作者: E-mail: gaozhang@ouc.edu.cn
收稿日期:2014-11-06;
修訂日期:2015-09-07
*基金項(xiàng)目:國(guó)家海洋公益性行業(yè)科研專(zhuān)項(xiàng)經(jīng)費(fèi)項(xiàng)目(201405010;201305043)資助
Supported by National Marine public welfare industry research special funds project (201405010;201305043)
