草魚-鰱-鯉混養生態系統的EwE模型分析*
2016-05-12 03:03:39董雙林
中國海洋大學學報(自然科學版) 2016年4期
關鍵詞:生態系統
周 波, 董雙林, 王 芳
(1.中國海洋大學海水養殖教育部重點實驗室,山東 青島 266003; 2.四川省農業科學院水產研究所,四川 宜賓 644003)
?
草魚-鰱-鯉混養生態系統的EwE模型分析*
周波1, 2, 董雙林1**, 王芳1
(1.中國海洋大學海水養殖教育部重點實驗室,山東 青島 266003; 2.四川省農業科學院水產研究所,四川 宜賓 644003)
摘要:采用EwE模型軟件,構建了一個具有14個生物功能組的草魚、鰱和鯉混養生態系統EwE模型,對草魚、鰱和鯉混養生態系統的結構和功能進行綜合量化分析。研究表明,草魚、鰱和鯉混養生態系統主要由3個營養級構成。從營養物質流量看,營養級Ⅰ流量最大,占系統總流量(TST)的56.90%;營養級Ⅱ、Ⅲ、Ⅳ、Ⅴ的流量隨營養級的增加而遞減,分別占總流量的34.45%、8.20%、0.44%和0.003%。食物網和營養級之間營養流動分析表明,系統營養流通的主要途徑為從浮游植物開始的牧食鏈、從碎屑開始的腐食鏈和從飼料開始的飼料鏈。從生態營養學效率(EE值)看,除螺類的EE值為零外,大部分功能組的EE值都較高,表明系統中大部分功能組都得到了較好的利用。碎屑在草魚、鰱和鯉混養生態系統中具有十分重要的作用,其主要來源是細菌、原生動物和浮游植物,且碎屑的EE值較高(水中為0.903,底泥為0.551),表明大部分碎屑重新進入食物鏈循環,碎屑得到了再利用。研究結果表明,草魚、鰱和鯉混養的模式可以進一步優化,建議增加放養魚類的密度,同時引進一些其他魚類(如青魚和鳙魚),以提高系統的綜合效益。
關鍵詞:生態系統; 草魚; 鰱; 鯉; EwE模型; 混養
引用格式:周波,董雙林,王芳. 草魚-鰱-鯉混養生態系統的EwE模型分析[J].中國海洋大學學報(自然科學版),2016, 46(4): 28-36.
ZHOU Bo, DONG Shuang-Lin, WANG Fang. Modeling analysis of the strueture of grass carp-silver carp-common carp polycultured pond ecosystem [J].Periodical of Ocean University of China, 2016, 46(4): 28-36.
混養(Polyculture or co-culture)是根據水生生物的棲息水層和食性等把不同種類的水生生物放在同一水體進行養殖,在中國有著悠久的歷史。混養的理念符合現代提倡的綜合水產養殖(Integrated aquaculture)概念,是綜合水產養殖中最重要的一類[1]。有關混養的種類和比例搭配研究(即養殖結構優化研究)是目前水產養殖生態系統研究中最活躍的研究領域之一。在早期的研究中,研究人員多用一些產量、經濟效益、對投入物質的利用率、水質等指標對系統進行評判[2-5],近些年人們開始利用養殖系統的N、P收支、光合作用效率等一些生態學指標來評判、優化養殖結構[6-13]。但總的來說,以往的大部分研究還只是就一些經濟和宏觀生態指標進行比較分析,尚缺乏綜合環境、經濟、能量利用等方面因素系統而全面的比較分析,特別是對生態系統內部的物質流動和能量傳遞效率等的分析十分缺乏。
EwE(Ecopath with Ecosim)模型是根據營養動力學原理,在生態系統食物網結構基礎上對能量流動進行描述的生態系統營養平衡模型。EwE模型的基本功能是對生態系統的結構、營養流動過程、營養動力學特征進行量化綜合分析。基于EwE模型的強大功能,目前已成為漁業生態系統研究中最廣泛使用的方法,但目前國內外對EwE模型的研究多用于海洋和大型湖泊等自然生態系統[14-16],在養殖生態系統尚鮮有報道。為此,本文以草魚、鰱和鯉混養生態系統為對象,用EwE模型方法對系統的結構和功能特征進行了量化和分析,為養殖系統的評價和進一步結構優化提供科學依據。
1材料和方法
1.1 研究對象
本實驗采用陸基圍隔實驗法在山東省淡水水產研究所進行(36°40′57.46″N, 116°51′9.04″E),選取3口大小相同、處理完全一致的草魚、鰱和鯉混養生態系統為研究對象。圍隔建于面積0.27hm2(90m×30m),平均水深(1.50±0.12)m 的淡水池塘中,每個面積為64m2(8m×8m)。時間單位為一個完整的養殖周期(從2010年5月17日—10月14日,150d)。模型所采用的數據如無特殊說明均來自于本系統的現場實驗和測定,所有數據均采用平均值。
1.2 模型構建
1.2.1 基本方程組EwE模型采用一系列線性方程描述生態系統各個功能組的能量流動或者物質流動平衡,并通過建立食物網絡使得整個生態系統達到平衡,從而得到一個生態系統特定時間的靜態平衡模型。用于平衡每一個功能組的基本方程為[17-18]:
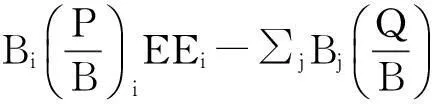
1.2.2 功能組設置根據模型的功能組設置原則,本研究將草魚、鰱和鯉混養生態系統劃分為14個生態功能組,即:草魚(Grass carp)、鰱(Silver carp)、鯉(Common carp)、橈足類(Copepoda)、枝角類(Cladocera)、輪蟲(Rotifera)、原生動物(Protozoa)、螺(Gastropoda)、水中細菌(Bacteria in water)、底泥細菌(Bacteria in sediment)、浮游植物(Phytoplankton)、人工飼料(Artificial feed)、水中碎屑(Detritus in water)和底泥碎屑(Detritus in sediment)。其中,由于細菌和碎屑在水體和底泥中的利用存在一定差別,因此分別分為水中細菌和底泥細菌,以及水中碎屑和底泥碎屑。人工投喂飼料為非生命組分,僅為系統提供能量來源,因此作為又一碎屑組處理。
1.2.3 數據來源對每一功能組,要求必須輸入的參數包括生物量(B)、P/B系數、Q/B系數以及食物組成矩陣等。生物量(B)通過現場實驗或調查獲得。草魚、鰱和鯉的B,P/B和Q/B值是通過放養量、收獲量、餌料系數以及每月的平均采樣重量計算而來。浮游植物的B,P/B和Q/B值則是通過黑白瓶法現場實驗計算,細菌的B和P/B值是通過平板計數法和16S rDNA序列分析法,但是其Q/B值是參考文獻[20]。對于其他功能組,生物量(B)通過每月的平均采樣密度計算而來,而P/B和Q/B值則是參考文獻[21-27],各功能組的基本輸入參數值詳見表1。此外,利用同位素分析技術對鰱的食物來源進行了分析,而其他功能組的食物來源則主要參考文獻[20,28-30],具體的食物組成如表2所示。
1.3 模型平衡
在模型平衡調試過程中,我們首先將EE<1作為其基本限制條件。如果某一功能組的EE>1,則逐步調整其食物組成(每次±5%),直到所有功能組的EE<1。然后檢查所有的功能組的呼吸值(R)是否為負值,如果存在這種情況,則按照活動能力越強,R/B值越高的原則,通過調整其P/B或Q/B值,使模型達到平衡并進行一致性檢驗。
1.4 模型可信度的評價
在EwE模型中采用Pedigree指數對模型質量(可信度)進行評價,Pedigree指數值越高,表明模型可信度越大,越接近于系統的真實狀況,其評價標準詳見Christensen等[31]的描述。
2結果
經過參數的輸入、平衡和估計,構建了草魚、鰱和鯉魚養殖生態系統的EwE模型,模型的Pedigree指數值為0.810,遠高于湖泊生態系統如太湖0.514[32]、海水養殖生態系統0.664[33]以及其他現有的研究報道,表明模型具有較高的可信度。
2.1 生態營養學效率(EE)
草魚、鰱和鯉混養生態系統EwE模型的基本參數見表1,其中生態營養學效率(EE)值為模型估計值。從結果可以看出,大部分功能組的EE值都較高。其中,放養魚類的EE值都大于0.95。人工飼料組EE值高達0.952,可能是由于大部分人工飼料被養殖動物直接攝食,同時溶失于水中部分又通過碎屑或細菌組重新在系統中再利用,而目前的投飼率又相對較低的結果。浮游植物組的EE值為0.609,表明系統中有將近40%的初級生產未得到充分利用。浮游動物組的EE值從原生動物的0.123到輪蟲組的0.730不等,其中原生動物組的利用率最低,僅為0.123,而其他浮游動物組(包括橈足類、枝角類和輪蟲)的EE值都在0.5以上(0.577~0.85)。系統中細菌組的EE值也較高(>0.85),表明以細菌組為媒介的微型食物鏈在系統中占據比較重要的地位,其中水中細菌組的利用率(0.914)較底泥細菌組(0.884)略高。水中碎屑組的EE值為0.903,表明大部分水體中的碎屑經過循環再利用重新進入了食物網。而底泥碎屑組的EE值僅為0.551,雖然高于太湖生態系統的0.379[32],但其循環再利用率較水中碎屑組要低得多。此外,值得注意的是,系統中螺類組的EE值為0,表明該功能組在生態系統中沒有得到利用。

表1 草魚-鰱-鯉混養生態系統EwE模型的基本輸入(黑體為模型估計值)

表2、表3、表4和圖1、圖3參見表1的表注。Table 2, table 3, table 4 and figure 1, figure 3 were in accordance with table 1.
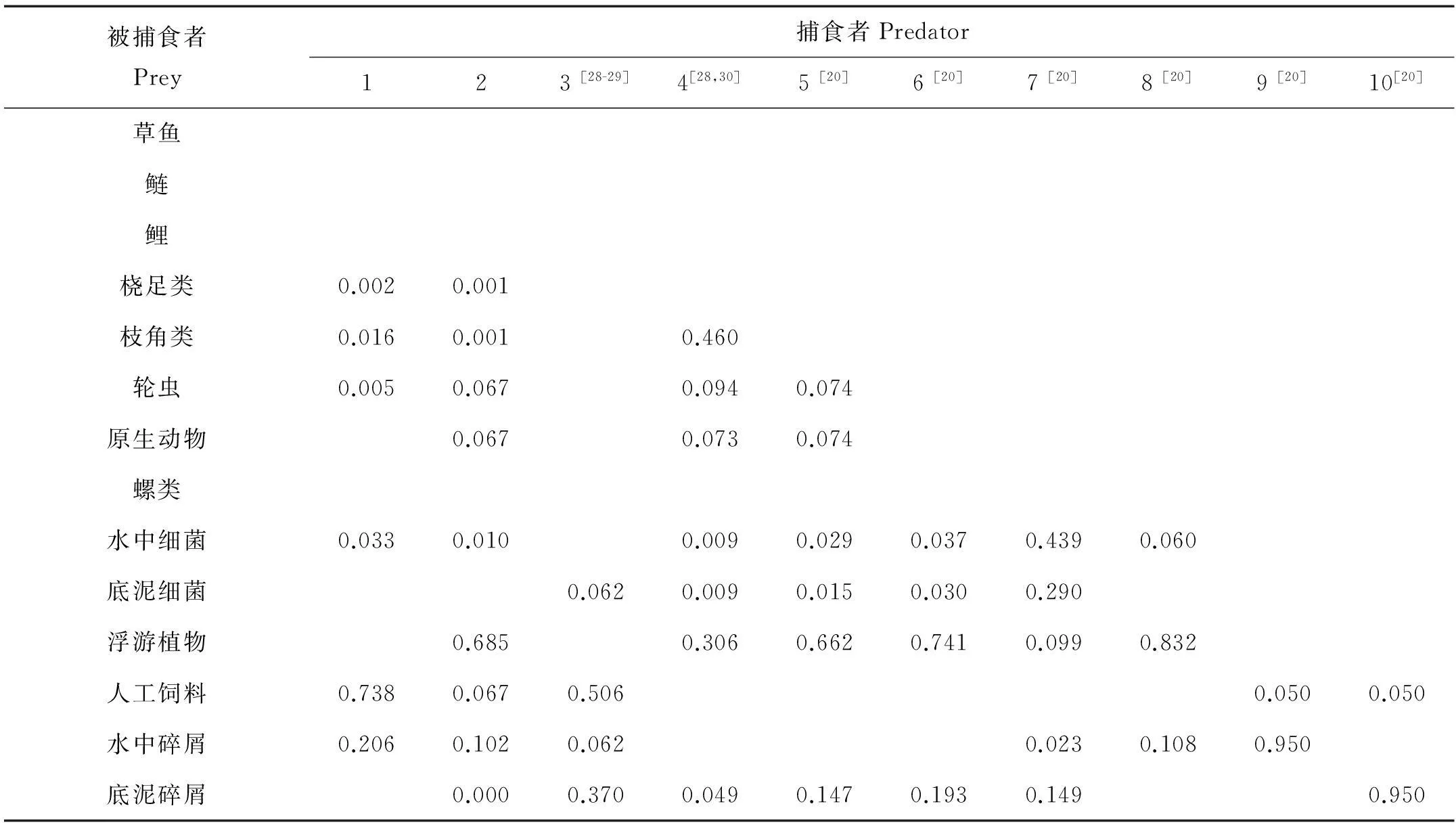
表2 草魚-鰱-鯉混養生態系統EwE模型的食物組成矩陣
2.2 營養級結構
模型將系統的營養物質流合并為6個整合營養級,但從營養級Ⅳ到營養級Ⅵ的流量、生產量和生物量都非常低。因此,草魚、鰱和鯉混養生態系統實際上主要由3個營養級構成,其流量、生物量和生產量的分布均呈金字塔型(見圖1)。營養級Ⅰ由初級生產者(浮游植物)和碎屑(包括人工飼料、底泥碎屑和水中碎屑功能組)占據,橈足類和原生動物可歸并為營養級Ⅲ,其他功能組則屬于營養級Ⅱ。
由于整合營養級是生態系統內復雜食物網絡關系的簡化,不能反映各功能組營養物質流在各營養級間的分布情況,目前較傾向于使用有效營養級(Effective trophic level)的概念。模型計算的草魚、鰱和鯉混養生態系統各功能組的有效營養級如表3 所示。從表上可以看出,有效營養級最高的為橈足類,值為2.82。
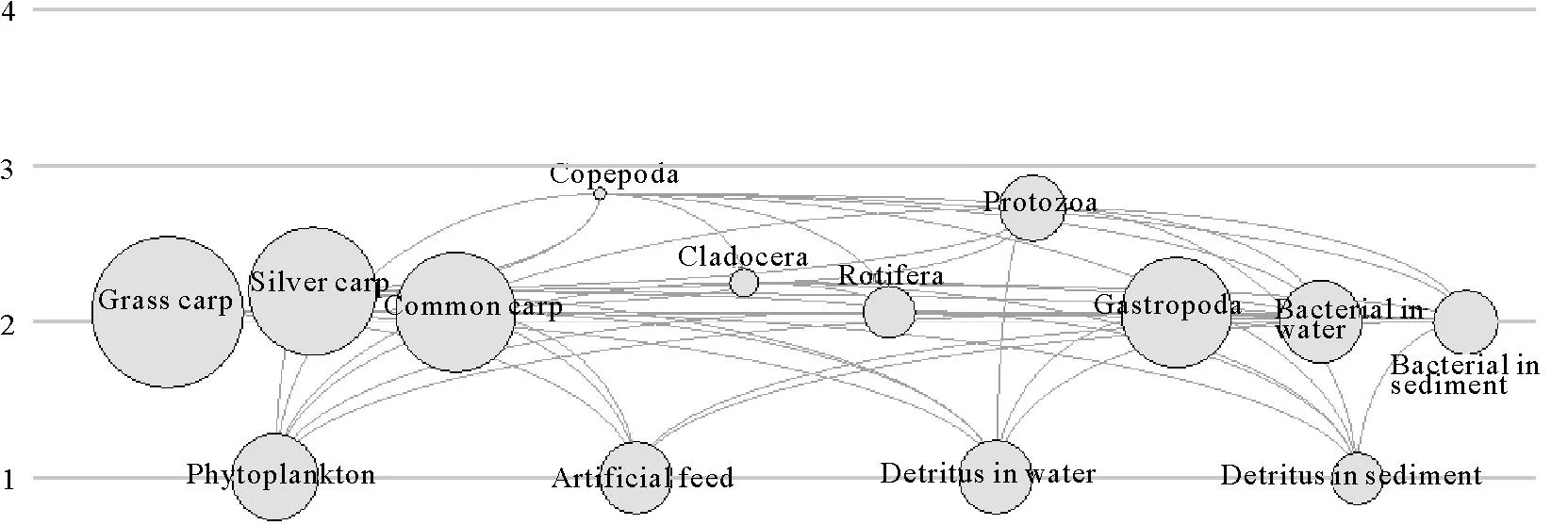
圖1 草魚、鰱和鯉混養生態系統各功能組的營養級與能流
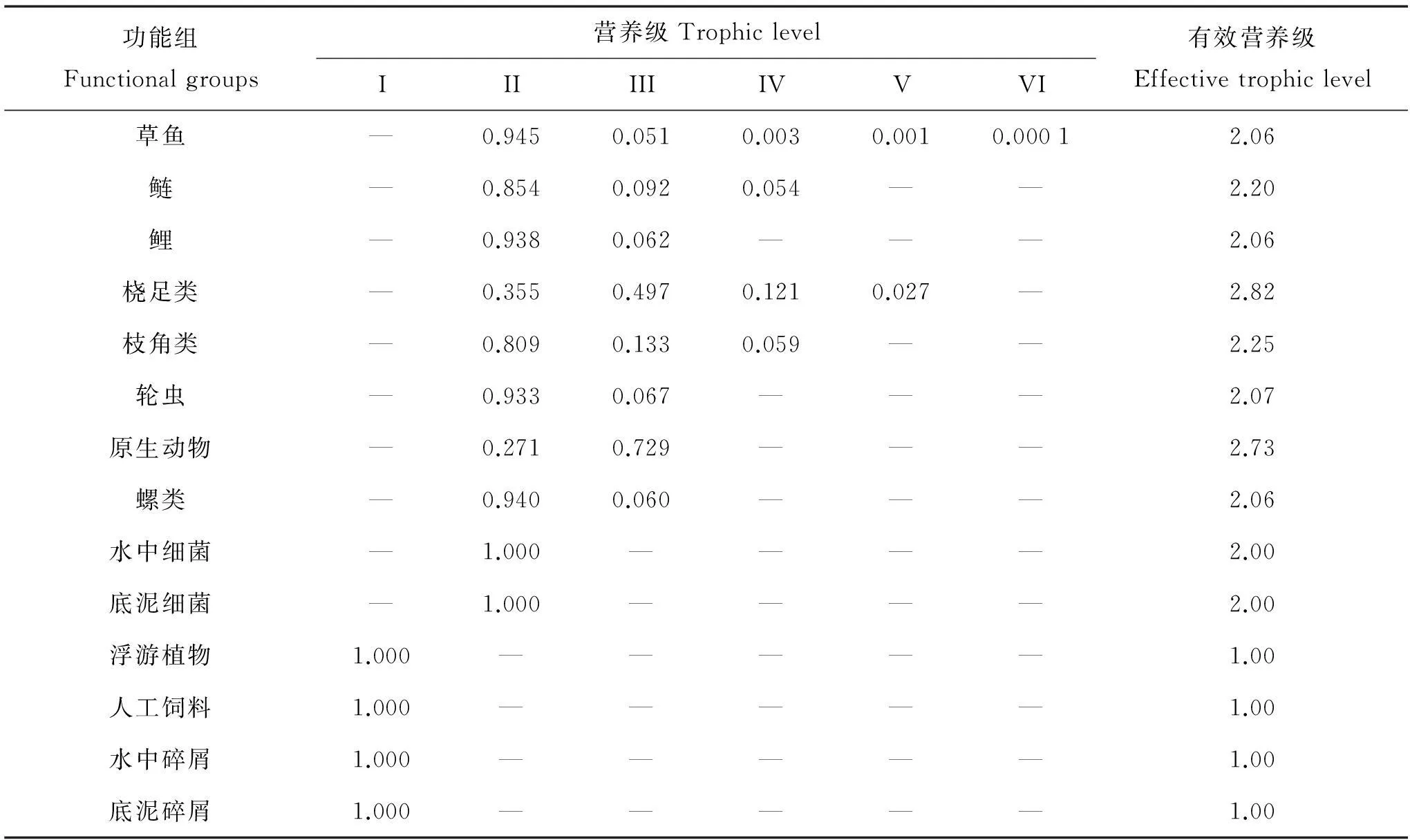
功能組Functionalgroups營養級TrophiclevelIIIIIIIVVVI有效營養級Effectivetrophiclevel草魚—0.9450.0510.0030.0010.00012.06鰱—0.8540.0920.054——2.20鯉—0.9380.062———2.06橈足類—0.3550.4970.1210.027—2.82枝角類—0.8090.1330.059——2.25輪蟲—0.9330.067———2.07原生動物—0.2710.729———2.73螺類—0.9400.060———2.06水中細菌—1.000————2.00底泥細菌—1.000————2.00浮游植物1.000—————1.00人工飼料1.000—————1.00水中碎屑1.000—————1.00底泥碎屑1.000—————1.00
2.3 營養級間的物質流動
草魚、鰱和鯉混養生態系統各營養級間的物質流動模型計算結果如圖2所示。從圖2可以看出,初級生產者的生產量為3298g·m-2·a-1,其營養物質流量約占系統總流量(Total system throughput, TST)的17.68%。其中1288 g·m-2·a-1流向了碎屑組,2010 g·m-2·a-1被營養級Ⅱ所攝食(占初級生產的60.9%)。各營養級流向碎屑組(包括水中碎屑和底泥碎屑組)的量總計6653g·m-2·a-1,約占系統TST的39.22%。其中1288g·m-2·a-1來自初級生產者,其他營養級5365g·m-2·a-1。雖然碎屑組流量較大,但大部分(4983g·m-2·a-1)可在系統中重新進入食物鏈循環得到再利用,只有1670 g·m-2·a-1的碎屑沉積于池底。總的來說,營養級Ⅰ(包括初級生產者和碎屑組)的流量最大,占系統TST的56.90%,營養級Ⅱ、Ⅲ、Ⅳ、Ⅴ的總流量隨其營養級的增加而遞減,分別為34.45%、8.20%、0.44%和0.003%。其中,營養級Ⅳ和Ⅴ的總流量都小于0.5%,可以忽略。此外,營養級Ⅱ的傳輸效率(TE)達到了24.4%,遠高于太湖生態系統[32],而與對蝦高位池塘精養模式生態系統相當[34]。

圖2 草魚、鰱和鯉魚混養生態系統各營養級之間的物質流動
2.4 混合營養效應分析
混合營養效應分析(Mixed trophic impact, MTI)反映的是系統各功能組間直接或間接的相互關系(有利或有害),因此也可以作為系統敏感性分析的一種方式。其取值范圍在-1和1之間,有利取正值,有害取負值。此外通過對食物網的綜合分析,MTI還可以用來估計某個功能組生物量變化時對其他功能組生物量的影響[31]。
圖3為草魚、鰱和鯉混養生態系統的混合營養效應分析結果。從圖3可以看出,所有的功能組都對自身有一定的負面影響。草魚對橈足類和人工飼料有一定的負面影響。鰱除對細菌組(包括水中細菌和底泥細菌)有一些有利影響外,對其他各功能組均存在一定的不利影響,其中影響最大的是輪蟲。而鯉對其他功能組幾乎沒有影響。浮游植物對大部分的捕食者(除原生動物組)都存在一定的正面影響,而對營養級Ⅰ的其他各功能組均存在一定的不利影響。細菌組對自身和碎屑組(水中細菌對水中碎屑、底泥細菌對底泥碎屑)具有較大的不利影響,表明在細菌組存在較強烈的密度制約,并且較依賴于碎屑組。人工飼料組對草魚和鯉都有一定的正面影響,表明增加人工飼料的投喂,對草魚和鯉的產量增加具有一定的正效應,但對橈足類和枝角類會產生一定的間接負效應。
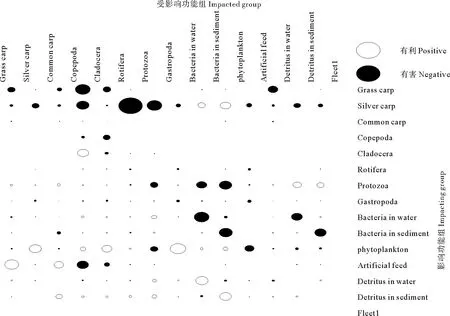
圖3 草魚、鰱和鯉混養生態系統混合營養效應分析
3討論
EwE模型目前主要應用于海洋和湖泊等自然生態系統,與這些生態系統相比,池塘養殖生態系統相對較小,生物組成相對簡單,對小微生物種群之間的關系也更為關注。為此,本文在對草魚、鰱和鯉混養生態系統進行模型構建和分析時,在更小尺度上對功能組進行了劃分,如浮游動物分成了橈足類、枝角類和輪蟲3個功能組。而在其他生態系統的研究中[35-36],這些功能組通常都是被歸并為一個功能組。從另一方面來說,功能組的細分也更有助于更好地理解系統內各種復雜的關系。模型的Pedigree指數值為0.810,表明模型具有較高的可靠性。同時模型模擬結果也與我們的專業判斷相符,為此,我們認為模型比較好的反映了系統的真實狀況,其結果是可信的。
3.1 食物鏈
通過模型構建和分析,可以大致概括出草魚、鰱和鯉混養生態系統營養流通的主要途徑:即從浮游植物開始的牧食鏈、從碎屑開始的腐食鏈和從飼料開始的飼料鏈。從營養物質流量看,幾條主要營養物質流的關系為:腐食鏈>牧食鏈>飼料鏈。
與千島湖生態系統相似,食物鏈短是草魚、鰱和鯉混養生態系統最顯著的特征[35]。系統中最高營養級僅為2.82(見表3),遠低于已報道的其他自然水域生態系統的研究結果[35-36]。人工飼料的投喂可能是造成食物鏈短的重要原因。由于人工飼料成為養殖魚類的主要食物來源,與自然生態系統相比,養殖魚類及其相關功能組的連接指數(Connectance index)和雜食指數(Omnivory index)大大降低,整個生態系統的雜食指數和連接指數也隨之降低[33]。此外,頂級捕食者的缺乏可能也是造成食物鏈短的一個原因,因為頂級捕食者的缺失將會縮短系統的食物鏈[35]。
3.2 碎屑流
營養物質流分析結果顯示,碎屑組的營養物質流量總計為6653g·m-2·a-1,約占系統TST的39.22%(見圖2),表明碎屑組在草、鰱和鯉混養生態系統中具有十分重要的作用。為了對碎屑的作用作進一步的分析,本文將碎屑組細分為水中碎屑和底泥碎屑2個功能組,同時對2個碎屑組的來源組成進行了分析(見表4)。結果表明,浮游植物和水中細菌是水中碎屑的主要來源,二者的貢獻基本相當,分別占34.12% 和31.74%。同時,底泥碎屑和原生動物也是水中碎屑的重要來源,分別占22.89%和5.76%。值得注意的是,系統中放養魚類僅占水中碎屑來源的1.74% (其中草魚0.44% ,鰱魚1.22% 和鯉魚0.08%)。人工飼料組更低,僅占0.28%。對于底泥碎屑,原生動物為其主要來源,占40.67%,其次是底泥細菌和浮游植物,分別占18.39% 和11.48%。相比于水中碎屑組,放養魚類對底泥碎屑的貢獻有所上升,占底泥碎屑來源的12.28% (其中草魚3.09% ,鰱魚 8.64% 和鯉魚0.55%)。人工飼料組仍然較低,為0.89%。總的來說,草魚、鰱和鯉混養生態系統中碎屑的主要來源是細菌、原生動物和浮游植物,而放養魚類和人工投喂飼料所占比重較低,不是其主要來源。
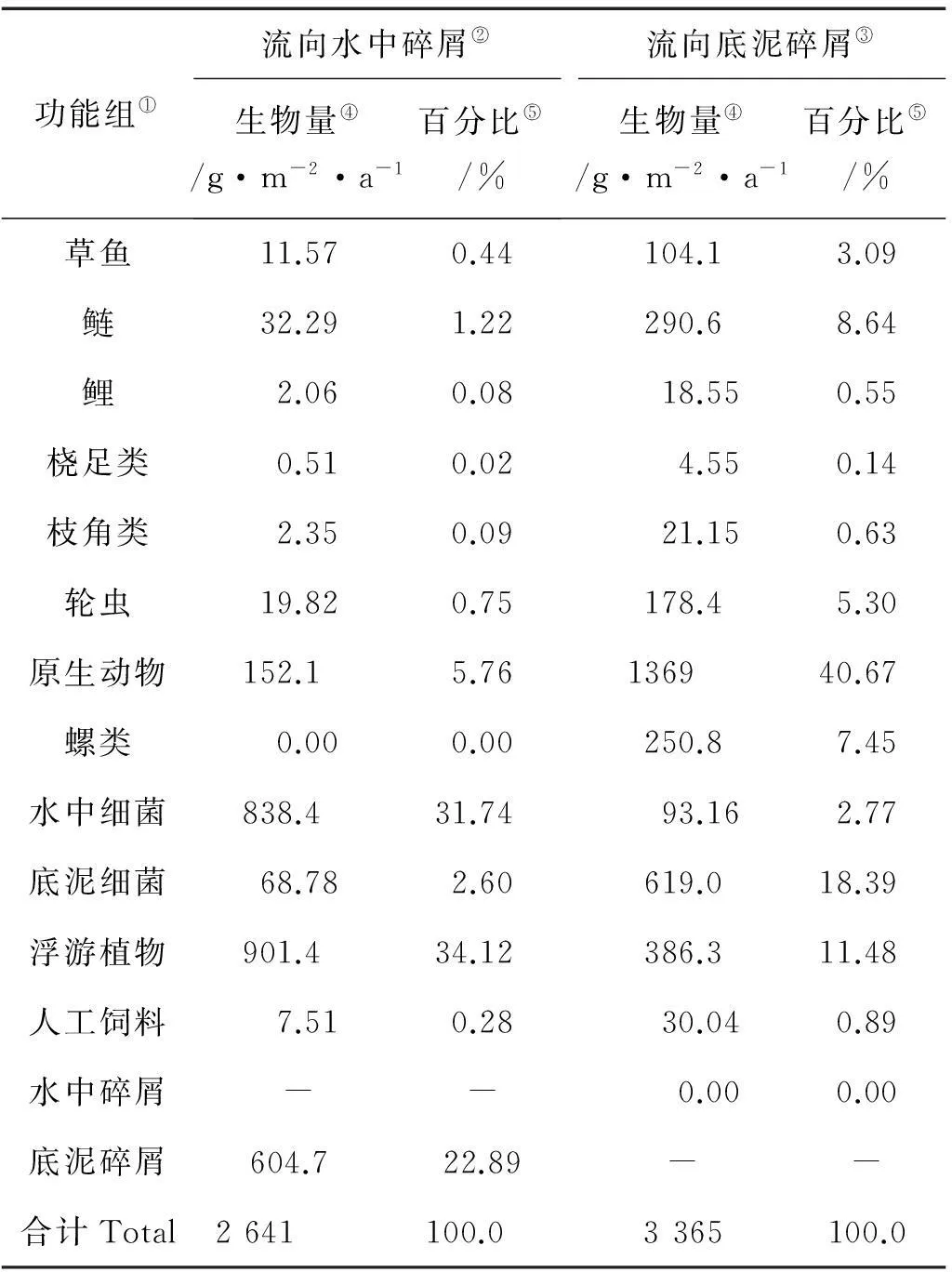
表4 草魚、鰱和鯉混養生態系統碎屑來源分析
Note:①Functional groups;②Flow to detritus in water;③Flow to detritus in sediment;④Biomass;⑤Percentage
此外,2個碎屑組的EE值都較高(其中:水中碎屑為0.903,底泥碎屑為0.551,見表1),表明大部分的碎屑重新進入食物鏈循環得到了再利用。特別是水中碎屑,高達90%重新進入了食物鏈循環,而底泥碎屑也有超過一半得到了重新再利用。
3.3 養殖結構優化
模型分析結果顯示,在草魚、鰱和鯉混養生態系統中大量的生產力(包括初級生產力和次級生產力)未得到充分利用,還可以進一步的優化。根據模型結果,有1288g·m-2·a-1的初級生產在系統中沒有得到利用,最終流向碎屑組。如果以10%的轉化率來計算,那么系統每年將增加128.8g·m-2·a-1的產品輸出。同樣地,浮游動物有226.8g·m-2·a-1沒有得到利用(其中橈足類占1.26%,枝角類占11.33%,輪蟲占87.41%),如果浮游動物食性魚類如鳙魚(Aristichthysnobilis)被引入系統,同樣以10%的轉化率計算,那么每年又可增加22.68g·m-2·a-1的產品輸出。螺類對系統產出也沒有貢獻,如果把一些可以攝食螺類的魚類如青魚(Mylopharyngodonpiceus)引入系統,那么系統的產出還將得到提高。
此外,混合營養效應分析表明人工飼料組對草魚和鯉都有一定的正面影響,如果增加人工飼料的投喂,對草魚和鯉的產量增加具有一定的正效應,這意味著目前的放養密度過低,對系統沒有產生壓力。為此,建議在目前的養殖模式下,增加放養魚類的密度,同時引進一些其他魚類(如青魚和鳙魚),以充分挖掘系統的生產潛力,大力提高系統的綜合效益。
4結論
(1)草魚、鰱和鯉混養生態系統主要由3個營養級構成。有效營養級最高的為橈足類,值為2.82。從營養物質流量看,營養級Ⅰ流量最大,占系統總流量(TST)的56.90%(其中初級生產者占17.68%,碎屑占39.22%),營養級Ⅱ、Ⅲ、Ⅳ、Ⅴ的總流量隨其營養級的增加而遞減,分別為34.45%、8.20%、0.44%和0.003%。
(2)草魚、鰱和鯉混養生態系統營養流通的主要途徑包括牧食鏈、腐食鏈和飼料鏈。且幾條主要營養物質流的關系為:腐食鏈>牧食鏈>飼料鏈。
(3)碎屑在草、鰱和鯉混養生態系統中具有十分重要的作用,其主要來源是細菌、原生動物和浮游植物,而放養魚類和人工投喂飼料所占比重較低。而且碎屑的EE值較高(水中碎屑組為0.903,底泥碎屑組為0.551),表明大部分的碎屑重新進入食物鏈循環得到了再利用。
(4)目前的養殖模式還可進一步優化。建議在目前的養殖模式下,增加放養魚類的密度,同時引進一些其他魚類(如青魚和鳙魚),以提高系統的綜合效益。
致謝:在野外實驗和現場調查中得到了山東省淡水研究所的大力支持,本研究團隊成員研究生張振東、宋頎、夏斌、柳炳俊、趙蕾、楊建雷、郭永堅等也做了大量的工作,在此一并致以誠摯地謝意!
參考文獻:
[1]董雙林. 中國綜合水產養殖的歷史、原理和分類[J]. 中國水產科學, 2011, 18(5): 1202-1209.
Dong S L. History, principles, and classification of integrated aquaculture in China[J]. Journal of Fishery Sciences of China, 2011, 18(5): 1202-1209.
[2]熊邦喜, 李德尚, 李琪, 等. 配養濾食性魚對投餌網箱養魚負荷力的影響[J]. 水生生物學報, 1993, 17(2): 131-144.
Xiong B X, Li D S, Li Q, et al. The impact of filter-feeding fish on the carrying capacity of reservoirs for feeding-cage-culture of fish[J]. Acta Hydrobiologica Sinica, 1993,17(2): 131-144.
[3]Wang J Q, Li D S, Dong S L, et al. Experimental studies on polyculture in closed shrimp ponds 1. Intensive polyculture of Chinese shrimp (Penaeuschinensis) with tilapia hybrid[J]. Aquaculture, 1998, 163: 11-27.
[4]Wang J Q, Li D S, Dong S L, et al. Experimental studies on intensive polyculture of Chinese shrimp with bay scallop[J]. Journal of Fishery Sciences of China, 1999, 6(1): 97-102.
[5]Tian X L, Li D S, Dong S L, et al. An experimental study on closed-polyculture of penaeid shrimp with tilapia and constricted tagelus[J]. Aquaculture, 2000, 202(3/4): 57-77.
[6]Frei M, Razzak M A, Hossain M M, et al. Performance of common carp,CyprinuscarpioL. andNiletilapia,Oreochromisniloticus(L.) in integrated rice-fish culture in Bangladesh[J]. Aquaculture, 2007, 262(2-4): 250-259.
[7]Jena J, Kar S, Das P C, et al. Olive barb, Puntius sarana (Hamilton) is a potential candidate species for introduction into the grow-out carp polyculture system[J]. Aquaculture, 2008, 280(1-4): 154-157.
[8]Rahman M M, Leopold A J, Nagelkerke, et al. Relationships among water quality, food resources, fish diet and fish growth in polyculture ponds: A multivariate approach[J]. Aquaculture, 2008, 275(1-4): 108-115.
[9]Rahman M M,Jo Q, Gong Y G, et al. A comparative study of common carp (CyprinuscarpioL.) and calbasu (LabeocalbasuHamilton) on bottom soil resuspension, water quality, nutrient accumulations, food intake and growth of fish in simulated rohu (LabeorohitaHamilton) ponds[J].Aquaculture, 2008, 285(1-4): 78-83.
[10]包杰, 田相利, 董雙林, 等. 對蝦、青蛤和江蘺混養的能量收支及轉化效率研究[J]. 中國海洋大學學報(自然科學版), 2006,36: 27-32.
Bao J, Tian X L, Dong S L,et al. An experiment study on the energy budget and conversion efficiency in ployculture of shrimp, bivalve and seaweed. [J].Periodical of Ocean University of China,2006, 36: 27-32.
[11]常杰, 田相利, 董雙林, 等. 對蝦、青蛤和江蘺混養系統氮磷收支的實驗研究[J]. 中國海洋大學學報(自然科學版), 2006, 36: 33-39.
Chang J, Tian X L, Dong S L, et al. An experimental study on nitrogen and phosphorus budgets in polyculture of shrimp, bivalve and seaweed [J]. Periodical of Ocean University of China,2006, 36: 33-39.
[12]宋頎, 田相利, 董雙林, 等. 草魚混養生態系統能量收支的研究[J]. 中國海洋大學學報(自然科學版), 2011, 41(10): 45-51.
Song Q, Tian X L, Dong S L, et al. An experiment study on the energy budget and conversion efficiency of ecosystem in ployculture of ctenopharyngodon idellus[J].Periodical of Ocean University of China, 2011, 41(10): 45-51.
[13]張振東, 王芳, 董雙林, 等.草魚、鰱魚和凡納濱對蝦多元化養殖系統結構優化的研究[J]. 中國海洋大學學報(自然科學版), 2011, 41(7/8): 60-66.
Zhang Z D, Wang F, Dong S L, et al. A preliminary study on structural optimization in polycultural systems of ctenopharyngodon idellus with hypophthalmichthys molitrix and litopenaeus vannamei[J]. Periodical of Ocean University of China, 2011, 41(7/8): 60-66.
[14]Christensen V, Walters C J. Ecopath with Ecosim: methods, capabilities and limitations[J]. Ecological Modelling, 2004, 172(2-4): 109-139.
[15]吳忠鑫, 張秀梅, 張磊, 等. 基于Ecopath模型的榮成俚島人工魚礁區生態系統結構和功能評價[J]. 應用生態學報, 2012, 23(10): 2878-2886.
Wu Z L, Zhang X M, Zhang L, et al. Structure and function of Lidao artificial reef ecosystem in Rongcheng of Shandong Province, East China: An evaluation based on Ecopath model[J].Chinese Journal of Applied Ecology, 2012, 23(10): 2878-2886.
[16]林群, 金顯仕, 張波, 等. 基于營養通道模型的渤海生態系統結構十年變化比較[J]. 生態學報, 2009, 29(7):3613-3620.
Lin Q, Jin X S, Zhang B, et al. Comparative study on the changes of the Bohai Sea ecosystem structure based on Ecopath model between ten years[J]. Acta ecologicasinica, 2009, 29(7):3613-3620.
[17]Christensen V, Pauly D. A guide to the Ecopath II software systems (version 2.1), ICLARM software 6[M]. Manila: International Center for Living Aquatic Resources Management (ICLARM), 1992: 72.
[18]Christensen V, Pauly D. Ecopath II: a software for balancing steady-state ecosystem models and calculating network characteristics[J]. Ecological Modelling, 1992, 61(3-4): 169-186.
[19]Christensen V, Walters C J, Pauly D. Ecopath with Ecosim: A User’s Guide (Version5.1) [M]. Vancouver, Canada: Fisheries Centre, University of British Columbia, 2005: 55-70,154.
[20]劉健康. 高級水生生物學[M]. 北京: 科學出版社, 1999: 154-248.
Liu J K .Advanced hydrobiology[M]. Beijing: Science Press, 1999: 154-248.
[21]Schwinghamer P, Hargrave B, Peer D, et al. Partitioning of production and respiration among size groups of organisms in an intertidal benthic community[J]. Marine Ecology Progress Series, 1986, 31: 131-142.
[22]Halfon E, Schito N, Ulanowicz R. Energy flow through the Lake Ontario food web: Conceptual model and an attempt at mass balance[J]. Ecol Model, 1996, 86: 1-36.
[23]Riddle M J, Alongi D M, Dayton P K, et al. Detrital pathways in a coral reef lagoon[J].Marine Biology, 1990, 104: 109-118.
[24]Vega-Cendejas M E, Arreguin-Sanchez F, Herandez M. Trophic fluxes on the Campeche Bank, Mexico[C]∥ Christensen V, Pauly D. Trophic Models of Aquatic Ecosystems. Manila Philippines: ICLARM Conference Proceedings, 1993: 206-213.
[25]張覺民, 何志輝. 內陸水域漁業自然資源調查手冊[M]. 北京: 農業出版社, 1991: 72-74.
Zhang J M, He Z H . Investigation handbook of fishery resources in inland waters[M]. Beijing: Agriculture Press, 1999: 72-74.
[26]Opitz S. Torphic interactions in Caribbean coral reefs[C]. Rome: ICLARM Technical Report, 1996: 43.
[27]閆云軍, 梁彥齡. 扁擔塘底棲動物群落的能量流動[J].生態學報, 2003, 23(3): 527-538.
Yan Y J, Liang Y L. Energy flow of macrozoobenthic community in a macrophytic lake, Biandantang Lake[J]. Acta Ecol Sin, 2003, 23(3): 527-538.
[28]Lin H J, Dai X X, Shao K T, et al. Trophic structure and functioning in a eutrophic and poorly-flushed lagoon in southern Taiwan[J]. Mar Environ Res, 2006, 62: 61-82.
[29]劉健康, 何碧梧. 中國淡水魚類養殖學[M].北京: 科學出版社, 1992: 427.
Liu J K, He B W . Pisciculture of Chinese freshwater fishes[M]. Beijing: Science Press, 1992: 427.
[30]施煒綱, 嚴小梅, 邴旭文.太湖秀麗白蝦生物學與食性的研究[J].湖泊科學, 1995, 7(1): 69-76.
Shi W G, YAN X M, Bin X W. Biology and feeding habit of Palaemon modestus (Heller) in TaiHu Lake[J]. Journal of Lake Science, 1995, 7(1):69-76.
[31]Christensen V, Walters C J, Pauly D. Ecopath with Ecosim: a user’s guide [R]. Vancouver: Fisheries Centre, University of British Columbia, 2005: 46-47.
[32]宋兵. 太湖漁業和環境的生態系統模型研究[D]. 上海: 華東師范大學, 2004.
Song B. EcoSystem modeling study on the fishery and environment of Taihu lake[D]. Shanghai: East China Normal University, 2004.
[33]Bayle-Sempere J T, Arreguín-Sánchez F, Sanchez-Jerez P, et al. Trophic structure and energy fluxes around a Mediterranean fish farm[J]. Ecol Model, 2013,248: 135-147.
[34]張天文. 對蝦高位池精養模式和生態養殖模式中碳流通特征的解析一Ecopath with Ecosim在養殖生態系統中應用的初步研究[D]. 青島: 中國海洋大學, 2011.
Zhang T W. The model of Carbon flux in intensive penaeid shrimp culture and ecological cultivation[D]. Qingdao: Ocean University of China, 2011.
[35]Liu Q G, Chen Y, Li J L, et al. The food web structure and ecosystem properties of a filter-feeding carps dominated deep reservoir ecosystem[J]. Ecol Model, 2007, 203: 279-289.
[36]Lin H J, Shao K T, Jan R Q, et al. A trophic model for the Danshuei River Estuary, a hypoxic estuary in northern Taiwan[J]. Mar Pollut Bull, 2007, 54: 1789-1800.
責任編輯朱寶象
Modeling Analysis of the Strueture of Grass Carp-Silver Carp-Common Carp Polycultured Pond Ecosystem
ZHOU Bo1,2, DONG Shuang-Lin1, WANG Fang1
(1.The Key Laboratory of Mariculture, Ministry of Education, Ocean University of China, Qingdao 266003, China; 2.Fishery Institute of Sichuan Academy of Agricultural Sciences, Yibin 644003, China)
Abstract:A grass carp, silver carp and common carp polyculture pond ecosystem was described using an Ecopath with Ecosim (EwE) model, aiming to characterize trophic structure, the interaction among ecological groups and assess the efficiency of pond ecosystem. Fourteen functional groups were incorporated into the model. The results indicated that this polyculture pond ecosystem was mainly composed of 3 aggregated trophic levels. The trophic flow analysis suggested that the trophic flow of trophic level Ⅰ was up to 56.90% of the total system throughput (TST), and the trophic flow from trophic level Ⅱ to trophic level Ⅴ decreased with the increase of the trophic level, 34.45% for trophic level Ⅱ, 8.20% for trophic level Ⅲ, 0.44% for trophic level Ⅳ and 0.003% for trophic level Ⅴ. The main way of the trophic flow in grass carp-silver carp-common carp polyculture pond ecosystem included pastoral food chain (starting from phytoplankton), rotten food chain (starting from detritus) and feed chain (starting from feed). The ecotrophic efficiency (EE) of most functional groups were relatively high (exception for gastropoda, the EE was nil), implying a high efficiency of the ecosystem. Detritus plays an important role in the ecosystem; its main source was bacteria, protozoa and phytoplankton. The EE of detritus was relatively high (0.903 for detritus in water, 0.551 for detritus in sediment), indicating most of detritus consumed and passed up the food web. Therefore, we propose to increase the density of culture species in and introduce other fish species (such as bighead carp and black carp) into the pondfor optimizing the structure in order to exploit the potential productivity of the culture pond system and increase overall efficiency.
Key words:ecosystem; grass carp; silver carp; common carp; Ecopath with Ecosim model; polyculture
DOI:10.16441/j.cnki.hdxb. 20150097
中圖法分類號:S964.3
文獻標志碼:A
文章編號:1672-5174(2016)04-028-09
作者簡介:周波(1981-),男,博士生,主要從事水產養殖生態學研究。E-mail:zhoubo2008@163.com**通訊作者: E-mail: dongsl@ouc.edu.cn
收稿日期:2015-03-27;
修訂日期:2015-06-12
*基金項目:國家重點基礎研究計劃項目(2009CB118706);山東省自然科學基金項目(ZR2014CM042)資助
Supported by National Basic Research Program of China (2009CB118706);Natural Science Fund Project of Shandong Province (ZR2014CM042)
猜你喜歡
綠色科技(2016年21期)2016-12-27 16:05:03
現代管理科學(2017年1期)2016-12-26 17:16:55
考試周刊(2016年21期)2016-12-16 10:37:45
當代經濟管理(2016年11期)2016-12-14 17:47:10
出版廣角(2016年14期)2016-12-13 01:49:53
新課程·中旬(2016年9期)2016-12-01 12:56:28
商情(2016年40期)2016-11-28 10:54:26
青年文學家(2016年30期)2016-11-22 19:02:32
科教導刊(2016年27期)2016-11-15 22:36:28
科技視界(2016年18期)2016-11-03 22:02:50
