氮、磷添加對亞高寒草甸地上生物量的影響
2016-05-17 11:24:29張仁懿史小明李文金王剛郭睿
生態(tài)科學(xué) 2016年5期
張仁懿, 史小明, 李文金, 王剛, 郭睿
氮、磷添加對亞高寒草甸地上生物量的影響
張仁懿1*, 史小明1, 李文金1, 王剛1, 郭睿2
1. 蘭州大學(xué)生命科學(xué)學(xué)院/草地與農(nóng)業(yè)生態(tài)系統(tǒng)國家重點實驗室, 甘肅 蘭州 730000
2. 蘭州城市學(xué)院城市發(fā)展研究院, 甘肅, 蘭州 730070
通過生長季后期及初期分別設(shè)置N-P養(yǎng)分添加實驗, 探討甘南亞高寒草甸群落及不同功能群生物量對養(yǎng)分添加的響應(yīng), 并確定提高生產(chǎn)力的最佳養(yǎng)分添加量。結(jié)果顯示, 隨著養(yǎng)分添加量的增加, 群落生物量表現(xiàn)出先升后降的趨勢。二次項擬合亞高寒草甸最佳N添加量為16.58 g·m–2, 最佳P添加量為4.50 g·m–2, 最佳N-P共添加量為N = 18.47 g·m–2、P = 7.39 g·m–2。N、P養(yǎng)分添加存在交互作用, 對群落生物量的促進作用N-P共添加 > N添加 > P添加。各添加處理下禾草均能做出積極的響應(yīng), 其中N-P共添加對禾草類生物量的促進作用最大; 雜草生物量隨生長季后期養(yǎng)分添加量下降, 而隨生長季初期養(yǎng)分添加量表現(xiàn)為先升后降的趨勢; 莎草生物量在單一N添加處理下隨添加量上升; P添加促進、而N添加及N-P共添加抑制豆科生物量的積累。
亞高寒草甸; 生物量; 養(yǎng)分添加; 功能群
1 前言
作為陸地植物的基本營養(yǎng)元素, N和P的供應(yīng)制約著陸地生態(tài)系統(tǒng)的大多數(shù)過程, 為陸地生態(tài)系統(tǒng)生產(chǎn)力的主要限制因素[1–2]。N、P養(yǎng)分的添加, 已成為提高植被生產(chǎn)力的最有效而快速的重要途徑[3]。養(yǎng)分添加及沉積實驗的研究表明, 增加N、P等養(yǎng)分的供應(yīng)(一種或多種)能夠增加植被的初級生產(chǎn)力, 但將同時改變?nèi)郝浣Y(jié)構(gòu)和功能, 往往伴隨著物種的喪失[4]。過量的養(yǎng)分添加甚至能對植物生長產(chǎn)生毒害作用, 抑制群落生物量的積累[5]。
自然群落中往往包括多個物種或功能群的植物,養(yǎng)分添加對它們的影響并非均等。不同功能群植物對土壤養(yǎng)分條件的適應(yīng)過程中采取不同策略, 對養(yǎng)分添加的響應(yīng)也存在差異。例如, 固氮植物有額外的N素來源, 不易受到土壤N素可用性的限制, 易受P素供應(yīng)的影響; 相反, 非固氮植物保持較高的N利用率以充分利用體內(nèi)低水平的N素[6]。全球尺度[5,7]和青藏高原高寒草甸[8]的研究均表明, 禾本科植物具有較高的N、P養(yǎng)分利用效率及N:P比值, 可能與其對養(yǎng)分添加作出快速的響應(yīng)相關(guān)[9–10]。不同功能群植物對土壤養(yǎng)分變化的響應(yīng)差異, 是群落中物種生態(tài)位互補與分化的結(jié)果, 也是實現(xiàn)物種多樣性、生產(chǎn)力最大化的前提[10]。
由于寒冷的氣候條件以及長期的過度放牧等因素的共同作用, 青藏高原高寒、亞高寒草甸土壤有效養(yǎng)分的供給不足, 成為限制植物生長的重要因子[3]。為了提高亞高寒草甸的初級生產(chǎn)力, 需找到最佳的N、P添加量。現(xiàn)有研究表明, 該地區(qū)的物種多樣性較高[8], 且不同功能群植物對養(yǎng)分添加的響應(yīng)存在差異, 然而其變化模式尚存在爭議[11–13]。本研究以甘南州合作市的亞高寒草甸為研究對象, 擬就以下問題展開研究: a.探討不同功能群植物對養(yǎng)分添加的響應(yīng)差異; b.尋找提高生產(chǎn)力的最佳養(yǎng)分添加量。
2 材料與方法
2.1研究區(qū)概況
研究地點位于甘南藏族自治州合作市的蘭州大學(xué)高寒草甸與濕地生態(tài)系統(tǒng)定位研究站合作站(N 34°55′, E 102°53′), 海拔約為3000 m。該樣地地處青藏高原東北部, 為寒溫濕潤的高原氣候, 根據(jù)合作氣象站的資料顯示, 該地年平均氣溫2.4 ℃,年平均降雨量530 mm, 年平均蒸發(fā)量1200 mm; 植被類型屬于多年生草本植物占優(yōu)勢的亞高寒草甸。
2.2實驗設(shè)計
2008年生長季后期(8月下旬), 選取實驗站禁牧區(qū)內(nèi)面積53 m×35 m的樣地, 劃分為54個5 m×5 m的區(qū)塊, 相鄰區(qū)塊間隔1 m避免邊緣效應(yīng)及作為過道。選取其中50個區(qū)塊進行隨機區(qū)組的養(yǎng)分添加處理, 分別為N添加、P添加、N-P共添加(質(zhì)量比1:1);養(yǎng)分添加量(以N、P計)為0、5、10、15 g·m–2。共10個處理, 每個處理設(shè)計5個重復(fù)。添加的養(yǎng)分均為速效肥, 其中N為尿素[CO(NH2)2, N% = 46%], P為磷酸二氫鈉(NaH2PO4, P% = 26%)。當(dāng)年及之后09-11年養(yǎng)分添加處理時間均為8月中旬。
為了探討N、P最佳添加量及添加時間的影響, 2012年生長季初期, 設(shè)置5 m×5 m的區(qū)塊90個, 區(qū)塊間距1 m, 進行養(yǎng)分添加實驗(尿素及磷酸二氫鈉)。添加組合為添加N、添加P、N-P共添加(比例2.5︰1); N添加量為0、2.5、5、10、20、30 g·m–2, P添加量為0、1、2、4、8、12 g·m–2, 添加梯度分別標(biāo)記為CK、1、2、3、4、5, 每個處理5個重復(fù)。
2.3生物量測定
2012年8月中旬進行生物量調(diào)查。各區(qū)塊選擇兩個0.5 m × 0.5 m的樣方, 分功能群(莎草、禾草、豆科及雜草)齊地剪下生物量, 70 ℃烘干至恒重。
2.4數(shù)據(jù)分析
同一處理下不同添加量的生物量差異用最小顯著性差異法(LSD)分析, N、P養(yǎng)分對生物量的影響采用多因素方差分析(General Linear Model Multivariate, GLM)處理。以折線圖表示養(yǎng)分添加量/添加序列與各功能群及群落生物量之間的關(guān)系, 其中生長季后期實驗以直線、生長季初期實驗以二項式進行擬合。分析及作圖功能分別在SPSS13.0(IBM Corp.)和Origin 8.0(OriginLab Corp.)上實現(xiàn)。
3 結(jié)果與分析
3.1群落生物量變化模式
生長季后期養(yǎng)分添加結(jié)果顯示, 群落生物量隨P的添加量顯著降低, 斜率為–3.99(P < 0.05), 而隨N及N-P共添加的添加量顯著上升, 斜率分別為9.53(P < 0.001)和17.32(P < 0.05), 其中N-P共添加對生物量的影響存在交互作用(圖 1, 表1)。
生長季初期N添加、P添加以及N-P共添加下, 群落生物量均表現(xiàn)出先上升后下降的趨勢。N添加、P添加以及N-P共添加對生物量最大增產(chǎn)量分別為47%、24%以及59%(圖 2)。N添加量及N-P共添加量與群落生物量呈顯著地拋物線關(guān)系(顯著性分別為P < 0.01, P < 0.001), 最佳N添加量擬合結(jié)果為16.58 g·m–2, 最佳N-P共添加量為N = 18.47 g·m–2, P = 7.39 g·m–2; 雖然二項式擬合結(jié)果不顯著(P > 0.05), 最佳P添加量擬合為4.50 g·m–2(未列出)。
3.2功能群生物量變化模式
生長季后期養(yǎng)分添加的結(jié)果顯示, 雜草生物量與養(yǎng)分添加量正相關(guān), 養(yǎng)分添加的抑制作用順序為: N添加> P添加>N-P共添加, 斜率分別為–4.55(P <0.05)、–3.73(P < 0.05)和–2.94(P < 0.05)(圖3a); N添加促進莎草生物量, 而P添加、N-P共添加抑制其生物量的積累(斜率分別為2.84、–1.00和–0.31, 顯著性均P > 0.05)(圖3b); 禾本科生物量與養(yǎng)分添加量正相關(guān), 養(yǎng)分添加的促進作用順序為: N-P共添加 >N添加 > P添加, 斜率分別為22.81(P < 0.05)、7.74 (P < 0.05)和3.94(P > 0.05)(圖3c); 養(yǎng)分添加抑制豆科生物量, 抑制作用N添加 > N-P添加> P添加, 斜率分別為–0.77、–0.38和–0.17, 顯著性均P > 0.05 (圖3d)。
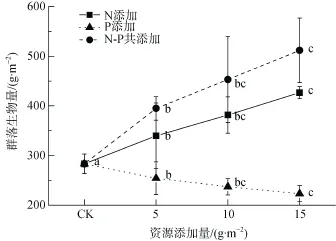
圖1 群落生物量與生長季后期養(yǎng)分添加量之間的關(guān)系。不同小寫字母表示不同添加量之間生物量差異顯著(P<0.05,最小顯著性差異法)Fig. 1 Relationship between the community biomass and the addition amount of late growing season. Different lowercase letters indicate significant differences among addition amounts (P < 0.05, LSD)
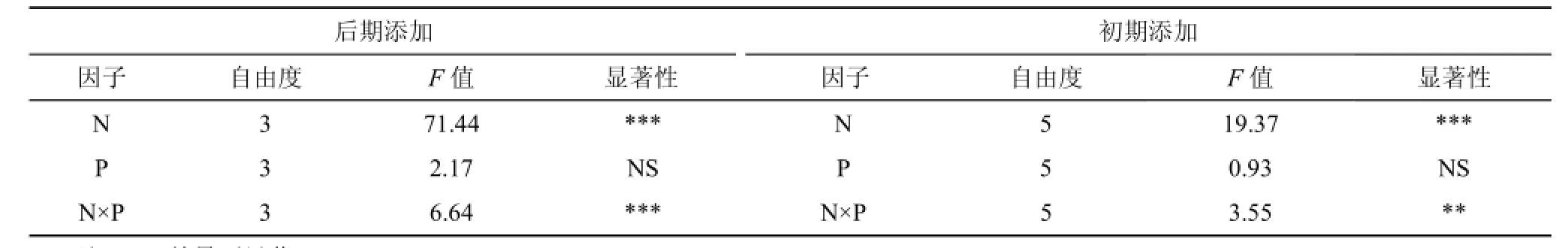
表1 生長季后期及初期N、P添加對生物量的GLM分析結(jié)果Tab. 1 F values and significance levels of general linear model multivariate (GLM) analysis for the effect of addition treatments on biomass
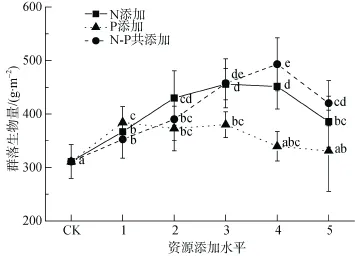
圖2 群落生物量與生長季初期養(yǎng)分添加量之間的關(guān)系。不同小寫字母表示不同添加量之間生物量差異顯著(P<0.05,最小顯著性差異法)Fig. 2 Relationship between the community biomass and the addition amount of early growing season. Different lowercase letters indicate significant differences among addition amounts (P < 0.05, LSD)

圖3 a)雜草、b)莎草、c)禾草以及d)豆科的生物量與生長季后期養(yǎng)分添加量之間的關(guān)系Fig. 3 Relationship between the a) forb, b) sedge, c) graminoid and d) legume biomass and the addition amount of late growing season
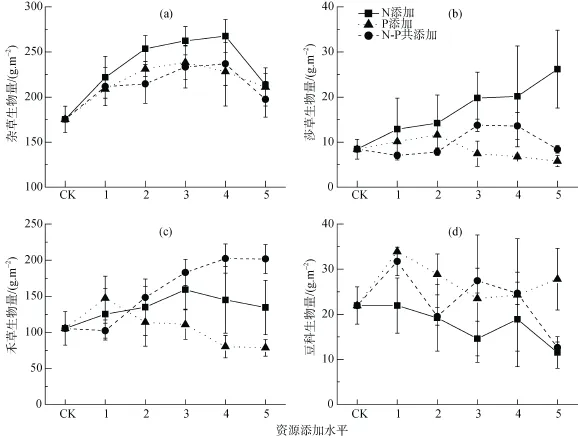
圖4 a)雜草、b)莎草、c)禾草以及d)豆科生物量與生長季初期養(yǎng)分添加量之間的關(guān)系Fig. 4 Relationship between the a) forb, b) sedge, c) graminoid and d) legume biomass and the addition amount of early growing season
生長季初期添加結(jié)果顯示, N添加對雜草生物量、莎草生物量的促進作用高于其它處理, 而對豆科生物量的積累起抑制作用; N-P共添加對禾草生物量的促進作用高于其它處理(圖4)。雜草生物量與養(yǎng)分添加量呈顯著地拋物線關(guān)系(顯著性均P <0.05), 最佳N添加量擬合為16.13 g·m–2, 最佳P添加量擬合為6.47 g·m–2, 最佳N-P共添加量擬合為N=16.07 g·m–2、P=6.43 g·m–2(圖4a); 莎草生物量與養(yǎng)分添加量二項式回歸僅在N添加下顯著(P < 0.05),最佳N添加量回歸為26.71 g·m–2, P添加及N-P共添加下實際最大生物量分別出現(xiàn)在P2: P = 2 g·m–2及NP3: N = 10 g·m–2、P = 3 g·m–2(圖4b); 禾草生物量與N添加量及N-P共添加量二項式回歸顯著(均為P < 0.05), 最佳N添加量擬合為17.03 g·m–2, 最佳共添加量為N = 22.33 g·m–2, P = 8.93 g·m–2, P添加下實際最大生物量出現(xiàn)在P1 = 2.5 g·m–2(圖4c); 除了N添加的抑制作用, 豆科生物量與養(yǎng)分添加量之間未發(fā)現(xiàn)一般性規(guī)律(圖4d)。
4 討論
青藏高原亞高寒草甸低溫作用的結(jié)果, 限制了土壤中礦質(zhì)養(yǎng)分的可利用性[6]。土壤N礦化過程是一個遵循Q10法則的生物過程, 而P的風(fēng)化屬于化學(xué)過程, 其溫度敏感性遠(yuǎn)低于N礦化過程[14]。因此, 該地區(qū)N可利用性比P更易受到低溫的抑制, 植被生產(chǎn)力主要受N供應(yīng)的影響[6,8,15]。在討論限制類型的研究中, 生態(tài)學(xué)者常常采用農(nóng)學(xué)中的標(biāo)準(zhǔn), 只考慮群落生產(chǎn)力是否受到限制[16]。不管是生長季后期還是生長季初期對亞高寒草甸進行的養(yǎng)分添加, N添加對地上生物量的促進作用均大于P(圖1, 圖2), 驗證了該地區(qū)植被主要養(yǎng)分限制類型為N限制。
植物具有調(diào)整體內(nèi)養(yǎng)分含量, 以適應(yīng)環(huán)境養(yǎng)分供應(yīng)條件變化的能力, 即使外界環(huán)境不斷變化,其組成也不會發(fā)生很大變化, 這種有機體與其環(huán)境保持一種相對穩(wěn)定的狀態(tài)就被稱為“內(nèi)穩(wěn)態(tài)(homeostasis )”[17]。然而, 對自然植被過量的養(yǎng)分添加量超出植物的內(nèi)穩(wěn)態(tài)調(diào)節(jié)能力時, 機體內(nèi)養(yǎng)分的過量積累反而對其產(chǎn)生毒害作用[5]。隨著養(yǎng)分添加量的上升, 亞高寒草甸群落生物量表現(xiàn)出先升后降的變化趨勢(圖2)。二項式回歸結(jié)果顯示, 增加該地區(qū)植被生產(chǎn)力的最佳N添加量為16.58 g·m–2, 最佳N-P共添加量為N = 18.47 g·m–2, P = 7.39 g·m–2, 這與沈振西等對高寒草甸的最佳N添加量的結(jié)果一致[3]; 而獲得最大生物量的實際P添加量為2.5 g·m–2,因此, 生長季后期養(yǎng)分添加實驗中, 5 g·m–2的最低P添加量可能已過量供應(yīng), 抑制了群落生物量的積累(圖1)。亞高寒草甸最佳共添加量總是高于單一養(yǎng)分的最佳添加量(圖2), 意味著多種類養(yǎng)分的添加,能夠減輕單一養(yǎng)分過量供應(yīng)對植物造成的毒害。
高寒草甸多種養(yǎng)分添加實驗表明, 添加養(yǎng)分種類越多, 群落地上生物量越高[15]。因此, 青藏高原高寒、亞高寒草甸生產(chǎn)力雖然主要受到N供應(yīng)的限制,但其它元素(如P、K、Si等)均對群落生物量產(chǎn)生影響[12;15;18]。從生態(tài)學(xué)角度嚴(yán)格的來看, 由于群落中物種所受限制因子多樣性, 植物群落并沒有限制因子的概念, 養(yǎng)分供應(yīng)變化對植物選擇的結(jié)果,只是引起物種的擴張與排除, 導(dǎo)致新舊群落的替代與生物量的變化[6]。亞高寒草甸N添加與P添加對群落生物量存在交互作用, 相對于單種養(yǎng)分的添加, N-P共添加對地上生物量的增產(chǎn)效應(yīng)較強(圖1, 圖2)。
不同功能群植物之間資源獲取方式、利用方式和對擾動響應(yīng)存在差異[5], 而這些差異是它們對養(yǎng)分條件的適應(yīng)過程中采取不同策略的前提或結(jié)果。結(jié)果顯示, 無論是生長季初期或后期的養(yǎng)分添加,各添加組合下禾草類植物都能做出積極的響應(yīng), 其中N-P共添加對禾草類生物量的促進作用最大(圖3c, 圖4c), 與其它高寒草甸進行的養(yǎng)分添加實驗結(jié)果一致[3,11,13,19], 這與其發(fā)達(dá)的根系及根系形態(tài)密不可分[9–10,20]。不同養(yǎng)分添加時間, 雜草生物量的變化模式也不同: 生長季后期養(yǎng)分添加處理隨添加量下降(圖3a), 而生長季初期養(yǎng)分添加實驗隨添加量表現(xiàn)出先升后降的趨勢(圖4a), 這可能與雜草植物中一年生植物占有一定比例[21], 其獲取的養(yǎng)分不能用于第二年有關(guān)。莎草生物量在單一N添加處理下隨添加量上升(圖4b), 這與韓潼等[22]的高寒草甸養(yǎng)分添加實驗的結(jié)果相近; 豆科植物的固氮作用使其具有額外的氮素來源, 其生長主要受P的限制, 因此, P添加促進、而N添加及N-P共添加抑制其生物量的積累(圖3d, 圖4d)。
通過選擇養(yǎng)分添加時間及添加量, 可以選擇性地提高特定功能群的生物量。例如, 生長季初期16.13 g·m–2的N添加量, 可以獲得最大的雜草生物量; 由于禾本科植物多為多年生, 添加時間應(yīng)為生長季后期, 最佳養(yǎng)分添加組合為22.33 g·m–2、4.46 g·m–2,以獲得最高的次年生物量; 莎草科植物最佳生長季初N添加量擬合為26.71 g·m–2, 但該添加量已超出本研究的梯度范圍, 其可靠性較低; 受其他限制因子的共同作用, 生長季初期的P添加, 可能是保持豆科生物量的有效方法。然而, 其作用機理涉及物種生理生態(tài)等多方面的特征, 為了更全面地探討?zhàn)B分添加與群落結(jié)構(gòu)的關(guān)系, 確定其內(nèi)在作用機制,尚需更進一步的研究支持。致謝:感謝蘭州大學(xué)高寒草甸與濕地生態(tài)系統(tǒng)定位研究站提供食宿條件及基礎(chǔ)實驗設(shè)施。
[1] LOVELOCK C E, FELLER I C, BALL M C, et al. Testing the Growth Rate vs. Geochemical Hypothesis for latitudinal variation in plant nutrients[J]. Ecology Letters, 2007, 10(12): 1154–1163.
[2] HE Jinsheng, WANG Liang, Flynn D F B, et al. Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes[J]. Oecologia, 2008, 155(2): 301–310.
[3] 沈振西, 陳佐忠, 周興民, 等. 高施氮量對高寒矮嵩草甸主要類群和多樣性及質(zhì)量的影響[J]. 草地學(xué)報, 2002, 10(1): 7–17.
[4] HAUTIER Y, NIKLAUS PA, HECTOR A. Competition for light causes plant biodiversity loss after eutrophication[J]. Science, 2009, 324(1): 636–638.
[5] GüSEWELL S. N:P ratios in terrestrial plants: variation and functional significance[J]. New Phytologist, 2004, 164(2): 243–266.
[6] K?RNER C. Alpine Plant Life: Functional Plant Ecology Of High Mountain Ecosystems[M]. New York: Springer Berlin Heidelberg, 2003.
[7] WRIGHT IJ, REICH PB, CORNELISSEN JHC. Assessing the generality of global leaf trait relationships[J]. New Phytologist, 2005, 166(2): 485–496.
[8] ZHANG Renyi, GOU Xin, BAI Yan, et al. Biomass fraction of graminoids and forbs in N-limited alpine grassland: N:P stoichiometry[J]. Polish Journal of Ecology, 2011, 59(1): 105–114.
[9] SCHERER-LORENZEN M, PALMBORG C, PRINZ A, et al. The role of plant diversity and composition for nitrate leaching in grasslands[J]. Ecology, 2003, 84(6): 1539–1552.
[10] ROSCHER C, THEIN S, SCHMID B, et al. Complementary nitrogen use among potentially dominant species in a biodiversity experiment varies between two years[J]. Journal of Ecology, 2008, 96(3): 477–488.
[11] 楊路存, 劉何春, 李長斌, 等. 氮磷鉀不同施肥配方對退化高寒草原植物群落結(jié)構(gòu)的影響[J]. 生態(tài)學(xué)雜志, 2015, 34(1): 25–32.
[12] 陳文業(yè), 戚登臣, 李廣宇, 等. 施肥對甘南高寒草甸退化草地植物群落多樣性和生產(chǎn)力的影響[J]. 中國農(nóng)業(yè)大學(xué)學(xué)報, 2009, 14(6): 31–36.
[13] 陳亞明, 李自珍, 杜國禎. 施肥對高寒草甸植物多樣性和經(jīng)濟類群的影響[J]. 西北植物學(xué)報, 2004, 24(3): 424–429.
[14] HEERWAARDEN LMV, TOET S, AERTS R. Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization[J]. Journal of Ecology, 2003, 91(6): 1060–1070.
[15] REN Zhengwei, LI Qi, CHU Chengjin, et al. Effects of resource additions on species richness and ANPP in an alpine meadow community[J]. Journal of Plant Ecology, 2010, 3(1): 25–31.
[16] CHAPIN FS, K?RNER C. Arctic and alpine biodiversity: patterns, causes and ecosystem consequences[M]. Springer-Verlag, 1995
[17] ELSER J, STERNER R, GOROKHOVA E, et al. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 2000, 3(6): 540–550.
[18] 賓振鈞, 張仁懿, 張文鵬, 等. 氮磷硅添加對青藏高原高寒草甸垂穗披堿草葉片碳氮磷的影響[J]. 生態(tài)學(xué)報, 2015, 35(14): 1–10.
[19] NIU Kechang, LUO Yanjiang, PHILIPPE C, et al. The role of biomass allocation strategy in diversity loss due to fertilization[J]. Basic and Applied Ecology, 2008, 9(5): 485–493.
[20] OELMANNA Y, WILCKEB W, TEMPERTONC VM, et al. Soil and plant nitrogen pools as related to plant diversity in an experimental grassland[J]. Soil Science Society of America Journal, 2007, 71(3): 720–729.
[21] 徐秀麗, 齊威, 卜海燕, 等. 青藏高原高寒草甸40種一年生植物種子的萌發(fā)特性研究[J]. 草業(yè)學(xué)報, 2007, 16(3): 74–80.
[22] 韓潼, 牛得草, 張永超, 等. 施肥對瑪曲縣高寒草甸植物多樣性及生產(chǎn)力的影響[J]. 草業(yè)科學(xué), 2011, 28(6): 926–930.
Effects of nitrogen and phosphorus addition on the plant aboveground biomass on a sub-alpine meadow
ZHANG Renyi1*, SHI Xiaoming1, LI Wenjin1, WANG Gang1, GUO Rui2
1.State Key Laboratory of Grassland and Agro-Ecosystems,School of Life Sciences,Lanzhou University,Lanzhou730000,China
2.City Development Institute,Lanzhou City University,Lanzhou730070,China
To examine response of different plant functional group biomass and community biomass to nutrient addition and determine the optimal nutrient level of plant productivity on a sub-alpine meadow, we conducted N-P addition experiments in the late and early growing season in Hezuo of Gansu Province. Results showed that with the increase of the amount of nutrients, community biomass increased firstly, and then decreased. The optimum amount of N addition and P addition was 16.58 g·m–2and 4.50 g·m–2respectively, and the optimum amount of N-P co-addition was N = 18.47 g·m–2and P = 7.39 g·m–2respectively according to the binomial regression fitting. Interaction of N and P addition was found and the order of positive effect on the community biomass was N-P co-addition > N addition > P addition. Graminoids biomass had a positive response to all the addition treatment, in which the effect of N-P co-addition was the strongest among all nutrient addition treatments. With the amount of nutrients forbs biomass in the early growing season firstly increased and then decreased, but decreased in the latter of the growing season. Sedge biomass, similar with the graminoids, increased with the amount of N addition. P addition favored the increase of legume biomass, while N addition and N-P co-addition, inhibited the legume.
sub-alpine meadow; biomass; nutrient addition; functional group
10.14108/j.cnki.1008-8873.2016.05.003
Q146
A
1008-8873(2016)05-015-06
張仁懿, 史小明, 李文金, 等. 氮、磷添加對亞高寒草甸地上生物量的影響[J]. 生態(tài)科學(xué), 2016, 35(5): 15-20.
ZHANG Renyi, SHI Xiaoming, LI Wenjin, et al. Effects of nitrogen and phosphorus addition on the plant aboveground biomass on a sub-alpine meadow[J]. Ecological Science, 2016, 35(5): 15-20.
2015-06-02;
2015-08-24
國家自然科學(xué)基金面上項目(31470480, 31270472); 國家自然科學(xué)基金青年基金項目(41201285)
張仁懿(1983—), 男, 福建三明人, 博士, 實驗師, 主要從事生態(tài)化學(xué)計量學(xué)研究, E-mail: zrenyi@lzu.edu.cn
*通信作者: 張仁懿, 男, 博士, 實驗師, 主要從事生態(tài)化學(xué)計量學(xué)研究, E-mail: zrenyi@lzu.edu.cn
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
現(xiàn)代農(nóng)業(yè)(2015年5期)2015-02-28 18:40:49
