低鹽脅迫對大底鳉(Fundulus grandis)血漿滲透壓、鰓上皮細胞形態和通道蛋白mRNA表達的影響
2016-05-17 11:24:34關穎GALVEZFernando張國霞張勝卿
生態科學 2016年5期
關穎, GALVEZ Fernando, 張國霞,*, 張勝卿
低鹽脅迫對大底鳉(Fundulus grandis)血漿滲透壓、鰓上皮細胞形態和通道蛋白mRNA表達的影響
關穎1, GALVEZ Fernando2, 張國霞1,*, 張勝卿3
1. 廣東省熱帶病研究重點實驗室,南方醫科大學公共衛生學院, 廣州 510515
2. 路易斯安那州立大學生物系, Baton Rouge 70802
3. 廣州舒國生物科技有限公司, 廣州510000
為了研究水環境中的低鹽度對大底鳉(Fundulus grandis)的適應性影響, 采用生理和熒光定量PCR方法探討了鹽度為5、2、1、0.5和0.1的情況下, 不同飼養時間大底鳉的血漿滲透壓, 鰓上皮細胞超微結構及通道蛋白mRNA表達的變化。飼養時間小于1 d、鹽度小于0.5的脅迫可以導致血漿滲透壓明顯降低(P<0.001); 鰓上皮表面泌氯細胞的體積增大、頂隱窩開口擴大或其細胞的形狀變為三角形或不規則形。當飼養時間超過3 d時, 血漿滲透壓以及鰓上皮表面泌氯細胞的形態都恢復到對照組水平 (鹽度為5); 低鹽脅迫上調了六種鰓通道蛋白mRNA的表達, 并下調了兩種鰓通道蛋白mRNA的表達。結果顯示: 廣鹽性的大底鳉通過調整鰓上皮的形態及通道蛋白mRNA的表達來維持機體滲透壓的平衡狀態。
鹽度; 大底鳉; 血漿滲透壓; 微結構; 通道蛋白; mRNA
1 前言
大底鳉(Fundulus grandis)為輻鰭魚綱鯉齒目鯉齒亞目底鳉科的一種, 分布于西大西洋佛羅里達至古巴西部海域及墨西哥灣沿岸[1]。大底鳉是廣鹽性硬骨魚類, 可適應0.1—35的鹽度變幅。當外界環境的滲透壓發生變化時, 大底鳉可通過滲透壓調節器官如鰓, 小腸和腎臟等維持體內滲透壓平衡。它獨特的鹽度適應性使其成為研究環境鹽度因素的良好模型[2–3]。鰓作為大底鳉主要滲透壓調節器官在維持機體滲透壓平衡以及適應環境鹽度變化中發揮重要的作用。在鰓絲和鰓小片表面覆蓋鰓上皮, 其作用是分隔細胞外液和周圍水環境。鰓上皮主要由泌氯細胞(chloride cells)和扁平細胞(pavement cells, PVC)構成。泌氯細胞主要分布在鰓小片的基底部, 為圓型或卵圓形的大細胞, 其形態分為海水型(seawatertype, SW-type)和淡水型(freshwater-type, FW-type)兩種形態。SW-type的泌氯細胞的表面呈卵圓形, 僅有小量的微絨毛伸向泌氯細胞的中心; 而FW-type的泌氯細胞表面形態不規則, 并且突出于周圍扁平細胞的表面, 且有微絨毛伸向泌氯細胞的中心[4]。泌氯細胞內含有豐富的線粒體和Na+-K+-ATP酶, 主要功能是分泌Cl–, 同時幫助Na+進行跨膜擴散[5]。Na+和Cl–是組成血漿滲透壓的主要的離子成分, 低鹽脅迫后, 硬骨魚類必須通過自身的調節來維持機體Na+和Cl–的平衡狀態。同時, 鰓上皮的通道蛋白(如水通道蛋白-3, 多種轉運蛋白如(Na+-K+-ATP酶)以及緊密連接蛋白如Claudins等也參與離子轉運, 從而保持血漿滲透壓的平衡[6–7]。
氣候變化如全球變暖, 冰川融化, 降雨量增加以及在潮汐的動力驅動下, 海水鹽分通過對流輸運和稀釋擴散等作用導致大底鳉生存環境鹽度的降低。近年來對大底鳉的生物學特征、酸堿平衡以及繁殖等方面的研究日益增多[8–9]。然而, 大底鳉在低鹽環境中的生存機制還不明確。為了探討大底鳉如何適應環境鹽度的變化特別是微小的鹽度變化, 本研究在實驗室中模擬低鹽環境來觀察低鹽脅迫對大底鳉血漿滲透壓、鰓超微結構以及通道蛋白mRNA表達的影響。實驗結果有助于了解大底鳉滲透壓調節的機制, 從而為大底鳉增養殖及資源保護提供理論支撐。
2 材料與方法
2.1實驗材料
實驗于2012年5月至2013年12月在美國路易斯安那州立大學生物學系進行。實驗用水是用鹽度為20的砂濾海水和爆氣自來水按比例配制, 溫度控制在(21±1) ℃。水系統一周部分換水2次, 保持連續充氣, 并通過生物過濾器和紫外線消毒裝置來對水系統進行消毒滅菌。實驗用魚是從墨西哥灣沿岸海域捕獲的天然大底鳉(Fundulus grandis), 日投餌量為其體質量的2%。實驗前, 將天然大底鳉暫養在鹽度為5的水系統中30天。
2.2實驗方法
2.2.1 實驗設計
實驗設置5個鹽度梯度分別為5、2、1、0.5和0.1, 其中鹽度5為對照組。實驗時, 將暫養在鹽度為5的水系統中的成年大底鳉(體質量4.3 -16.8 g)隨機分配到4個鹽度梯度的水系統中, 每個鹽度設置了3個平行對照。實驗的取樣時間為6 h, 1 d, 3 d和7 d, 從各實驗組隨機取9尾魚, 取血液及雙側鰓弓。取樣前12 h禁食。
2.2.2 大底鳉血液樣本的制備和血生化的測定
用MS-222(間氨基苯甲酸乙酯甲烷磺酸鹽)將大底鳉麻醉, 用毛細吸管在魚體臀鰭下方動脈取血,血液離心3000 r·min-1, 8 min至完全分層, 收集上清,采用Fiske210微型冰點滲透壓儀測定滲透壓。血漿中Na+和Cl–含量分別采用火焰原子吸收光譜法及硫氰酸鹽容量法[10]測定。
2.2.3 大底鳉鰓絲掃描電鏡樣本的制備
取左側鰓弓的第一條鰓絲, 用生理鹽水清洗干凈后, 用2.5%的戊二醛(用0.1(mol·L-1)磷酸緩沖液配制, pH 7.4)溶液固定, 固定后樣本在4 ℃下保存12 h,然后用0.1(mol·L-1)磷酸緩沖液(pH 7.4)沖洗15 min,重復3次; 再用體積分數為1%鋨酸溶液固定后,在4 ℃下保存2 h。用0.1(mol·L-1)磷酸緩沖液配制(pH7.4)沖洗三次, 每次15 min。然后用50%—100%乙醇逐級脫水, 醋酸異戊酯置換, 常規臨界干燥,真空離子鍍膜, 在JSM-6610掃描電鏡下觀察和拍照。
2.2.4 大底鳉鰓組織RNA提取及熒光定量PCR
用Trizol法提取鰓組織(左側第2、3、4條鰓絲及右側鰓弓)的總RNA。通過NanoDrop2000微量紫外分光光度儀(Thermo公司)檢測其純度和濃度, 控制260/280吸光值的比率在2至2.1的范圍內。總RNA經0.1%瓊脂糖凝膠電泳檢測RNA完整性, 采用High capacity cDNA Reverse Transcription試劑盒(Applied Biosystems)將RNA轉為單鏈cDNA。利用Primer 3.0軟件設計熒光定量PCR的特異性引物如表1所示。熒光定量PCR在ABI7500熒光定量PCR儀上進行, 總反應體系為20 μL(5 μL模板cDNA, 2 μL SYBR試劑,終濃度為10 nM的正向和反向引物)。反應程序為95 ℃(15 s), 60 ℃(1 min), 40個循環, 待檢測樣本和內參分別設置3個重復, 18s-rRNA為內參, 以確保特應性擴增。標準曲線有系統分析, 基因表達水平根據2–ΔΔCt計算相對定量[11]。
2.2.5 數據處理與分析
所得數據為均值±標準差(N=9), 應用SPSS20.0 統計軟件進行分析。數據分析采用析因設計單因素方差分析, 多個樣本比較采用單因素方差分析和SNK檢驗。
3 結果與分析
3.1低鹽度脅迫對大底鳉血漿滲透壓的影響
與鹽度為5的對照組相比, 將大底鳉置于鹽度為0.5的環境中6 h時, 血漿滲透壓迅速下降19% (P<0.001), 1 d后其值恢復到對照組水平; 當大底鳉置于鹽度為0.1的環境中6 h時, 血漿滲透壓降低了10%, 1 d內血漿滲透壓降低了20%, 達到最低點。而當環境鹽度降到2或1時, 即使增加大底鳉低鹽暴露的時間, 血漿滲透壓也無明顯變化(P=0.798或0.879)。可見, 只有環境鹽度降低到0.5以下時, 才會對血漿滲透壓產生明顯的影響。由表2可知, 血漿滲透壓降低的幅度依賴于鹽度的降低幅度以及低鹽環境暴露的時間。
3.2低鹽度脅迫對大底鳉血漿Na+和Cl–濃度的影響
血漿滲透壓主要由Na+和Cl–構成, 圖1結果顯示, 與鹽度為5的對照組相比, 當大底鳉置于鹽度為0.1的水系統中1 d, 血漿中Na+和Cl–濃度分別降低了18%和22% (P<0.001); 兩種離子濃度都能在3 d后再恢復到對照組的水平。
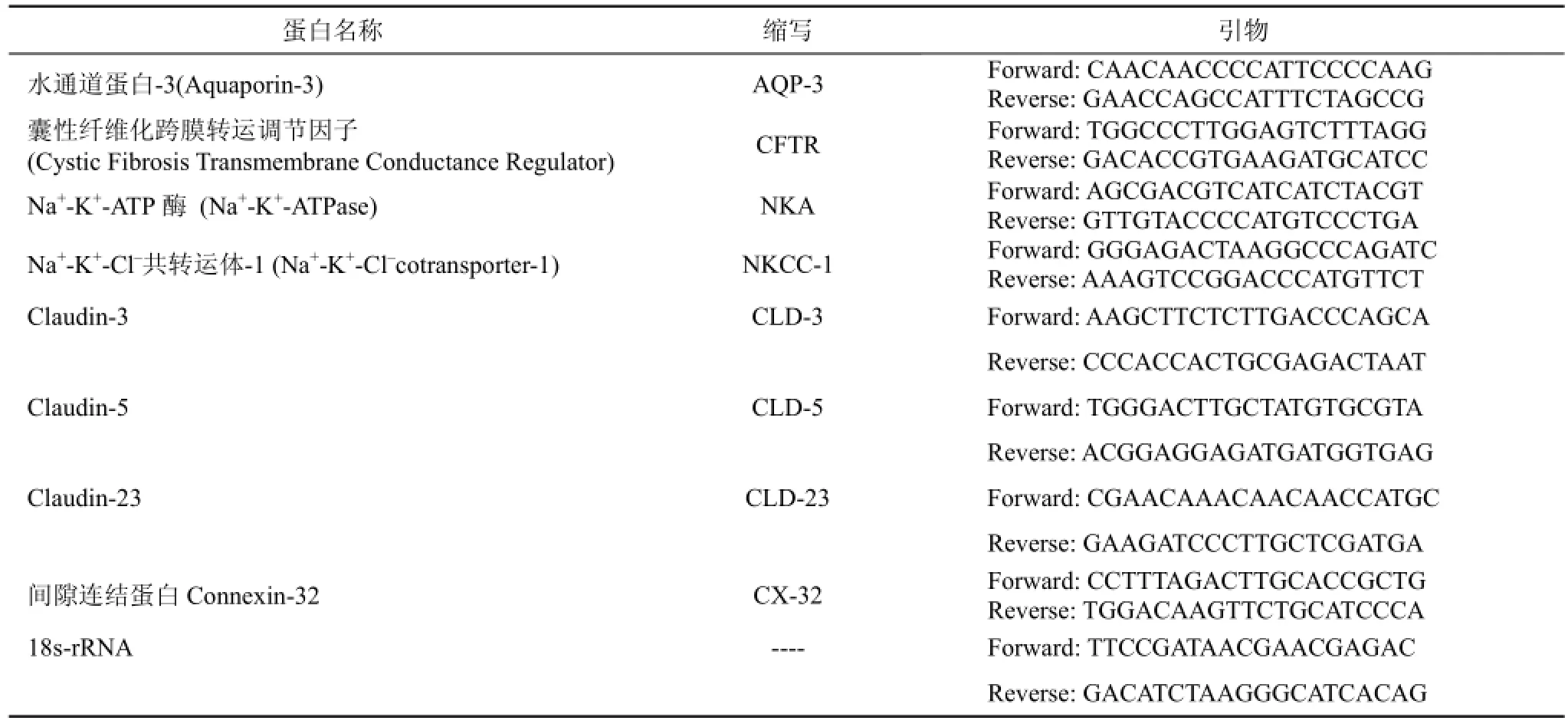
表1 熒光PCR引物序列Tab. 1 Sequences of forward and reverse primers used for quantitative PCR
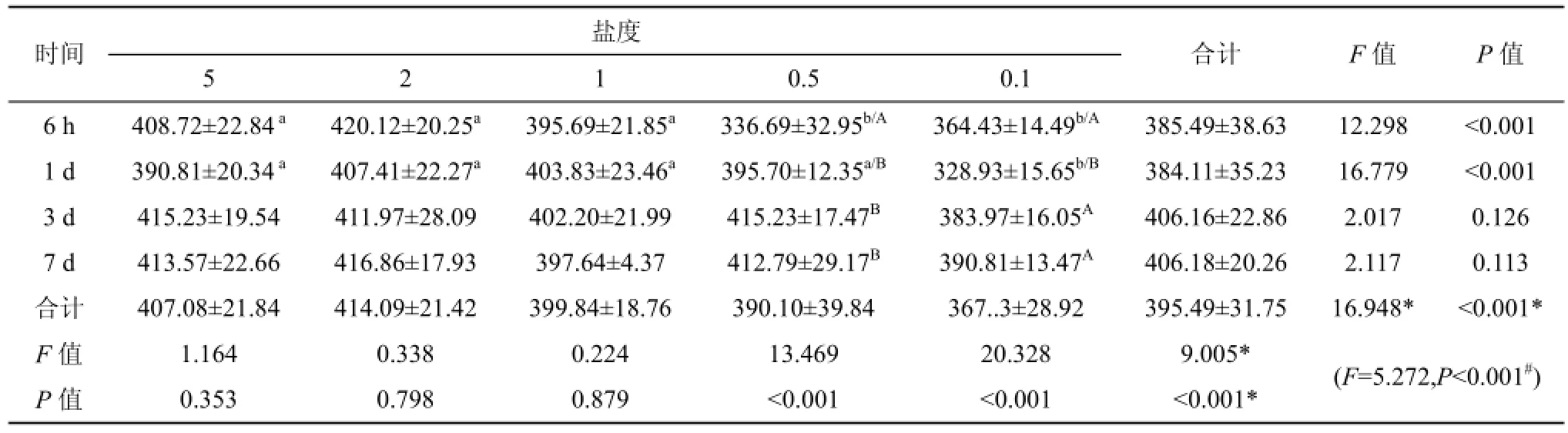
表2 鹽度對大底鳉血漿滲透壓(mOsm·Kg–1)的影響Tab. 2 The effects of different salinities on plasma osmolality (mOsm·Kg–1)
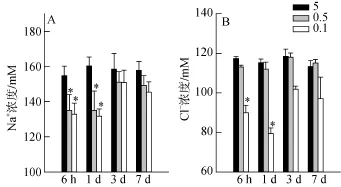
圖1 低鹽度脅迫對大底鳉血漿Na+和Cl–濃度的影響Fig. 1 Effects of low salinity stress on plasma Na+and Cl–concentration.
3.3低鹽度脅迫對大底鳉鰓絲超微結構的影響
短時間(≤1 d)的低鹽度環境的暴露會導致血漿的滲透壓和離子濃度的明顯降低; 特別是在環境鹽度降低到0.1 (表1, 圖1)。掃描電鏡的結果顯示: ①對照組鰓上皮細胞為多邊形, 形成了由環形隆起嵴組成的四邊形至六邊形不同形狀的圖案。與對照組比較, 在鹽度為2或1的環境中7 d內, 鰓上皮細胞的形態無明顯變化, 為海水型泌氯細胞形態; ②當環境鹽度降為0.5 (6 h)時, 泌氯細胞膨大, 體積明顯增大, 泌氯細胞的頂隱窩(apical crypt)開口擴大, 使得該上皮區域向外突出, 鋪蓋其上面的扁平細胞區域變小。這種變化在暴露1 d后發生好轉。當環境鹽度降為0.1 (6 h) 時, 泌氯細胞的形狀變為三角形或不規則形, 對周圍扁平細胞的擠壓更加明顯; 暴露時間延長到1 d, 泌氯細胞膨脹的更加劇烈, 已出現明顯的淡水型泌氯細胞的形態, 表現為鰓絲中泌氯細胞向外凸起, 并且可見發達的微絨毛[12]。3 d內泌氯細胞形態轉變為SW型(結果未顯示)。
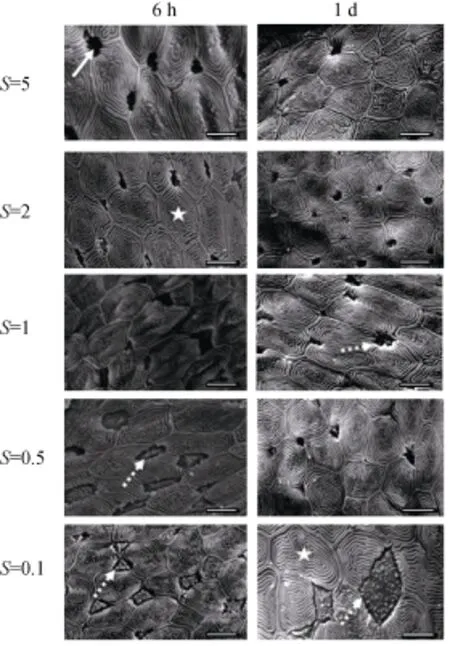
圖2 低鹽度脅迫對大底鳉上皮結構掃描電鏡觀察Fig. 2 Representative scanning electron micrographs of the afferent filamental epithelium of F. grandis.
3.4低鹽度脅迫對大底鳉鰓上皮通道蛋白mRNA表達的影響
隨著時間延長, 八種蛋白的mRNA表達水平都出現了明顯的變化。將對照組所有mRNA的表達水平定義為1, 和對照組比較, AQP-3和NKA的mRNA表達量分別增加了17倍(3 d)和2倍(7 d) (圖3A, 3C); CLD-3, 5, 23的mRNA表達量分別增加了2.3倍(1 d), 2.1倍(3 d)和4倍(6 h) (圖3E-G); CX-32mRNA表達量增加了2.1倍(1 d) (圖3H)。然而, 和對照組相比, CFTR和NKCC-1的mRNA表達量出現明顯下調趨勢, 分別下降了47% (3 d)和29% (3 d) (圖3B, 3D)。
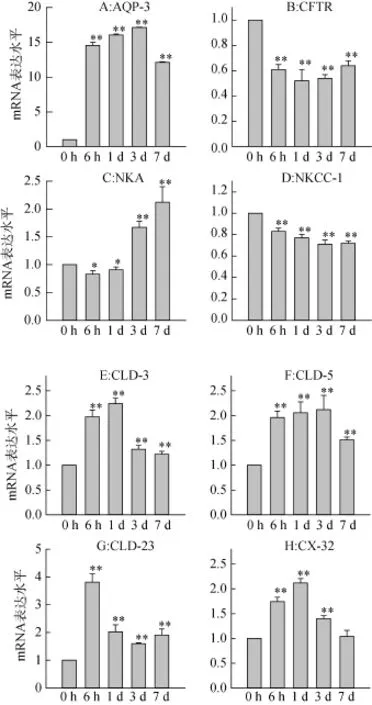
圖3 低鹽度脅迫對不同通道蛋白mRNA表達水平的影響Fig. 3 The effects of hypoosmotic exposure on the relative mRNA expressions of different transporters in gills of F. grandis.
4 討論
4.1低鹽度脅迫對血漿滲透壓及鰓上皮結構的影響
通過實驗室模擬的方法, 研究環境鹽度的微小變化對大底鳉血漿滲透壓, 鰓超微結構以及通道蛋白mRNA表達的影響。大底鳉對鹽度的適應過程可以分為兩個主要階段: 一是鹽度變化短期內(≤1 d),血漿滲透壓呈迅速下降的趨勢; 二是滲透壓向變化前的狀態逐漸恢復的過程。大底鳉在最初的1 d內是被動的適應外界環境, 低鹽環境刺激了機體滲透壓調節機制, 開始對滲透壓進行主動性的恢復。大底鳉對鹽度變化的適應能力很強(≤1 d), 不同種類的魚適應鹽度變化所需的時間不同, 如墨西哥灣鱘(Acipenser oxyrinchus de sotoi)[13], 由淡水向鹽水轉移后, 1 d內滲透壓可恢復; 而布氏棘鯛(Acanthopagrus butcheri)[14]由鹽水向淡水轉換后, 需要3-6個月才能恢復其滲透壓。本研究結果顯示血漿滲透壓表現出隨鹽度的下降而下降的趨勢(表1), 該結果和其他研究者的報道表現出相同的趨勢[15–16]。
環境鹽度微小的變化即從1降到0.5就會導致大底鳉血漿滲透壓和主要離子(Na+和Cl–)濃度的失衡(表2,圖1), 血漿滲透壓在大底鳉暴露于鹽度為0.5的鹽水中6 h就顯著降低, 但是其恢復速度較快(1 d內); 但是, 當鹽度降低到0.1時, 滲透壓降低更加明顯, 而且需要更長的時間來恢復到對照組的水平(表1)。與滲透壓變化相應的出現鰓上皮的形態的改變(圖2)。隨著血漿滲透壓的降低(≤0.5), 泌氯細胞體積膨大, 并出現泌氯細胞表面形態由SW型轉變為FW型, 該結果與其他研究一致[17–18]。同時, 低鹽脅迫過程中, 鰓上皮細胞出現了三角形的FW細胞, 它們與周圍的扁平細胞鋸齒狀相連, Laurent將這些特殊的FW泌氯細胞定義為立方體細胞, 這種細胞對離子的通透性差, 可以防止離子從鰓上皮細胞中丟失, 起到保持血漿離子濃度的作用[19]。雖然泌氯細胞在鰓上皮細胞中所占的比例小, 但是它們在硬骨魚類滲透壓調節中至關重要。鰓上皮細胞中的泌氯細胞的形態從SW型轉變為FW型, 導致了鰓上皮細胞的重塑。多種環境因素或生理因素都會導致硬骨魚鰓上皮細胞的重塑, 如缺氧可以導致鯽(Carassius carassius)層間細胞團(interlamellar cell mass, ILCM)增生[20], 而低氧和高溫皆可以導致鯽鰓上皮細胞ILCM凋亡而導細胞重塑[21]。雖然目前上皮細胞重塑的機制不詳, 但是這種快速的細胞形態的轉換有利于廣鹽性魚適應外周環境的急劇變化。
4.2低鹽脅迫對鰓上皮通道蛋白mRNA表達的影響
低鹽脅迫導致鰓上皮中六種通道蛋白mRNA表達增加, 包括AQP-3, NKA, Claudins-3, 5, 23以及CX-32(圖3)。水通道蛋白是一種位于細胞膜上的蛋白質, 在細胞膜上組成“孔道”, 可以控制水進出細胞。低鹽脅迫導致AQP-3 mRNA表達增加(圖3A),該結果和其它研究結果一致[8]; 高鹽脅迫下, AQP-3 mRNA在大西洋鮭(Salmo salar)[22]和歐洲鰻(Anguilla Anguilla)[23]鰓上皮中表達量降低。這些結果提示硬骨魚類可以通過水通道蛋白來調節水進出細胞來從而保持滲透壓的平衡。NKA不僅可以從外環境中吸收Na+, 它還可以提供離子主動轉運所需要的能量,其表達量隨低鹽暴露時間的延長而增加表示可能有更多耗能的通道蛋白參與到維持機體滲透壓的平衡中(圖3C)。而緊密連接蛋白如Claudins家族和Connexin家族在調節膜的滲透性和細胞旁通道離子轉運中起到關鍵作用[24–25]。鰓上皮中Claudins和CX-32 mRNA水平受環境鹽度的響應(圖3E-H), 但對Claudins蛋白的認知主要來源于哺乳動物, 其在魚類中的功能和分布還不明確[26]。Claudins mRNA表達增加可能和滲透壓的調節有關, 研究表明,一些緊密連接蛋白會形成“緊密”的連接孔, 而另一些更容易形成“滲透性”緊密連接孔[27], 這些所謂的“緊密”或“滲透”連接孔體現了細胞旁路的通透性[28]。由連接蛋白組成的間隙連接通道是細胞間離子的轉運通道, 其表達增加意味著離子有更多的機會通過細胞旁間隙。同時, 本研究結果表明低滲暴露導致大底鳉鰓組織內CFTR蛋白mRNA表達降低(圖3B), 該結果與前期研究結果一致[29]。本研究顯示低鹽脅迫導致鰓組織中NKCC-1 mRNA表達減低(圖3C), 免疫熒光的結果顯示NKCC表達在泌氯細胞中, 其利用電化學梯度來將Na+和Cl–轉入到胞質中, 先前的研究結果表明鹽度變化影響鱸魚(Morone saxatilis)[30]和底鳉(Fundulus heteroclitus)[8]魚鰓組織中NKCC 蛋白mRNA的表達, 但是具體的作用還不明確。
總之, 低鹽脅迫導致血漿滲透壓的降低, 大底鳉通過調整鰓上皮細胞的形態以及通道蛋白mRNA的表達來進行滲透壓的調整, 使其適應環境鹽度的變化, 并在低鹽環境中生存下來。
5 結論
廣鹽性的大底鳉通過調整鰓上皮的形態及通道蛋白mRNA的表達來維持機體滲透壓的平衡狀態,進而適應周圍環境鹽度的變化。
[1] PATTERSON J T, GREEN C C. Physiological management of dietary deficiency in n-3 fatty acids by spawning Gulf killifish (Fundulus grandis)[J]. Fish Physiology and Biochemistry, 2015, 41(4): 971–979.
[2] MARSHALL W S. Rapid regulation of NaCl secretion by estuarine teleost fish: coping strategies for short-duration freshwater exposures[J]. Biochimica et Biophysica Acta, 2003, 1618(2): 95–105.
[3] BURNETT K G, BAIN L J, BALDWIN W S, et al.Fundulusas the premier teleost model in environmental biology: Opportunities for new insights using genomics[J]. Comparative Biochemistry and Physiology Part D: Genomics Proteomics, 2007, 2(4): 257–286.
[4] EVANS D H, PIERMARINI P M, CHOE K P. The multifunctional fish gill: dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste[J]. Physiology Reviews, 2005, 85(1): 97–177.
[5] WHITEHEAD A. The evolutionary radiation of diverse osmotolerant physiologies in Killifish (Fundulussp.)[J]. Evolution, 2010, 64(7): 2070–2085.
[6] WHITEHEAD A, GALVEZ F, ZHANG Shujun, et al. Functional genomics of physiological plasticity and local adaptation in killifish[J]. Journal of Heredity, 2011, 102(5): 499–511.
[7] KOLOSOV D, BUI P, CHASIOTIS H, et al. Claudins in teleost fishes[J]. Tissue Barriers, 2013, 1(3): e25391.
[8] WHITEHEAD A, ROACH J L, ZHANG Shujun, et al. Salinity- and population dependent genome regulatory response during osmotic acclimation in the killifish (Fundulus heteroclitus) gill[J]. Journal of Experimental Biology, 2012, 215(8): 1293–1305.
[9] BROWN C, GOTHREAUX C, GREEN C. Effects of temperature and salinity during incubation on hatching and yolk utilization of Gulf killifishFundulus grandisembryos[J]. Aquaculture, 2011, 315(3): 335–339.
[10] MUNNS R, WALLACE P A, TEAKLE N L, et al. Measuring soluble ion concentrations Na+, K+, Cl–in salt-treated plants[J]. Methods in Molecular Biology, 2010, 639(23): 371–382.
[11] FLEIGE S, WALF V, HUCH S, et al. Comparison of relative mRNA quantification models and the impact of RNA integrity in quantitative real-time RT-PCR[J]. Biotechnology Letters, 2006, 28(19): 1601–1613.
[12] KATOH F, HASEGAWA S, KITA J, et al. Distinct seawater and freshwater types of chloride cells in killifish,Fundulus heteroclitus[J]. Canadian Journal of Zoology, 2001, 79(5): 822–829.
[13] ALTINOK I, CHAPMAN F A, GALLI S M. Ionic andosmotic regulation capabilities of juvenile Gulf of Mexico sturgeon,Acipenser oxyrinchus de sotoi[J] . Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 1998, 120(4): 609–616.
[14] GAVIN J P, GREG I J. The effect of salinity on growth and survival of juvenile black bream (Acanthopagrus butcheri)[J]. Aquaculture, 2002, 210(1): 219–230.
[15] KOZAK G M, BRENNAN R S, BERDAN E L, et al. Functional and population genomic divergence within and between two species of killifish adapted to different osmotic niches[J]. Evolution, 2014, 68(1): 63–80.
[16] PATTERSON J, BODINIER C, GREEN C. Effects of low salinity media on growth, condition, and gill ion transporter expression in juvenile Gulf killifish,Fundulus grandis[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2012, 161(4): 415–421.
[17] KATOH F, KANEKO T. Short-term transformation and long-term replacement of branchial chloride cells in killifish transferred from seawater to freshwater, revealed by morphofunctional observations and a newly established 'time-differential double fluorescent staining' technique[J]. Journal of Experimental Biology, 2003, 206(22): 4113–4123.
[18] WILSON J M, LAURENT P. Fish gill morphology: inside out[J]. Journal of Experimental Zoology, 2002, 293(3): 192–213.
[19] LAURENT P, CHEVALIER C, WOOD C M. Appearance of cuboidal cells in relation to salinity in gills ofFundulus heteroclitus, a species exhibiting branchial Na+but not Cluptake in freshwater[J]. Cell and Tissue Research, 2006, 325(3): 481–492.
[20] SOLLID J, DE ANGELIS P, GUNDERSEN K, et al. Hypoxia induces adaptive and reversible gross morphological changes in crucian carp gills[J]. Journal of Experimental Biology, 2003, 206(20): 3667–3673.
[21] SOLLID J, NILSSON G E. Plasticity of respiratory structures - adaptive remodeling of fish gills induced by ambient oxygen and temperature[J]. Respiratory Physiology & Neurobiology, 2006, 154(1/2): 241–251.
[22] TIPSMARK C K, S?RENSEN K J, MADSEN S S. Aquaporin expression dynamics in osmoregulatory tissues of Atlantic salmon during smoltification and seawater acclimation[J]. Journal of Experimental Biology, 2010, 213(3): 368–379.
[23] CUTLER C P, CRAMB G. Branchial expression of an aquaporin 3 (AQP-3) homologue is downregualted in the European eel (Anguilla anguilla) following seawater acclimation[J]. Journal of Experimental Biology, 2002, 205(17): 2643–2651.
[24] VAN ITALIE C M, ANDERSON J M. Claudins and epithelial paracellular transport[J]. Annual Review of Physiology, 2006, 68(1): 403–429.
[25] CARLISLE T C, RIBERA A B. Connexin 35b expression in the spinal cord of Danio rerio embryos and larvae[J]. Journal of Comparative Neurology, 2014, 522(4): 861–875.
[26] CLELLAND E S, BUI P, BAGHERIE-LACHIDAN M, et al. Spatial and salinity-induced alterations in claudin-3 isoform mRNA along the gastrointestinal tract of the pufferfish Tetraodon nigroviridis[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2010, 155(2): 154–163.
[27] SAS D, HU M, MOE OW, et al. Effect of claudins 6 and 9 on paracellular permeability in MDCK II cells[J]. American Journal of Physiology: Regulatory, Integrative and Comparative Physiology, 2008, 295(5): 1713–R1719.
[28] COYNE C B, GAMBLING T M, BOUCHER R C, et al. Role of claudin interactions in airway tight junctional permeability[J]. American Journal of Physiology: Lung Cellular and Molecular Physiology, 2003, 285(5): 1166–1178.
[29] SCOTT G R, CLAIBORNE J B, EDWARDS SL, et al. Gene expression after freshwater transfer in gills and opercular epithelia of killifish: insight into divergent mechanisms of ion transport[J]. Journal of Experimental Biology, 2005, 208(14): 2719–2729.
[30] TIPSMARK C K, MADSEN S S, BORSKI R J. Effect of salinity on expression of branchial ion transporters in striped bass (Morone saxatilis)[J]. Journal of Experimental Zoology Part A: Comparative Experimental Biology, 2004, 301(12): 979–991.
GUAN Ying1, GALVEZ Fernando2, ZHANG Guoxia1,*, ZHANG Shengqin3
1.Guangdong Provincial Key Laboratory of Tropical Disease Research,School of Public Health,Southern Medical University,Guangzhou510515,China
2.Department of Biology,Louisiana State University,Baton Rouge70802,USA
3.Guangzhou Shuguo Bio-Technique Co.Ltd.,Guangzhou510000,China
The objective of this paper was to describe the effect of hypoosmotic challenge on the plasma osmolality, ultrastructure of branchial epithelia, and mRNA expression of branchial transporters inFundulus grandis. Adult fish were reared in the salinity from 5 (control) to 2, 1, 0.5 and 0.1, and fish were randomly sampled from each salinity at 6 h, 1 d, 3 d, and 7 d. The results showed that plasma osmolality decreased significantly only in fish transferred to the salinity at 0.5 and 0.1, but recovered to the salinity at 5 control values by 1 d and 3 d, respectively. The ultrastructure of branchial epithelia showed that the surface of apical crypt was larger, and the volume of chloride cell swelled within 6 h in the salinity at 0.5. In salinity at 0.1,chloride cells showed a triangle or irregular shape, and they squeezed surrounding pavement cells. The mRNA expression of six branchial transporters was up-regulated, and the levels of two transporters were down-regulated during hypoosmotic challenges. The regulation of the gill morphology and mRNA expressions of branchial transporters may contribute to the freshwateradaption inFundulus grandis
salinity;Fundulus grandis; plasma osmolality; ultrastructure;transporters; mRNA
10.14108/j.cnki.1008-8873.2016.05.007
Q175
A
1008-8873(2016)05-043-07
關穎, GALVEZ Fernando, 張國霞, 等. 低鹽脅迫對大底鳉(Fundulus grandis)血漿滲透壓、鰓上皮細胞形態和通道蛋白mRNA表達的影響[J]. 生態科學, 2016, 35(5): 43-49.
GUAN Ying, GALVEZ Fernando, ZHANG Guoxia, et al. Effects of hypoosmotic challenges on plasma osmolality, the morphology of gill epithelia and mRNA expression of branchial transporters inFundulus grandis[J]. Ecological Science, 2016, 35(5): 43-49.
2015-06-26;
2015-08-14
廣東省醫學科研基金(B2014234); 南方醫科大學公共衛生與熱帶醫學學院院長基金(GW201431, GW201435)
關穎(1980—), 女, 黑龍江七臺河人, 博士, 講師, 主要從事水生生物適應性研究, E-mail: ynguan@echobelt.org
*通信作者: 張國霞, 女, 博士, 副教授, 主要從事環境和人體微生物組學, E-mail:guoxiazhang@126.com
Effects of hypoosmotic challenges on plasma osmolality, the morphology of gill epithelia and mRNA expression of branchial transporters in Fundulus grandis
猜你喜歡
現代臨床醫學(2022年4期)2022-09-29 07:38:00
中老年保健(2021年12期)2021-08-24 03:30:40
昆明醫科大學學報(2021年4期)2021-07-23 01:21:50
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
云南醫藥(2019年3期)2019-07-25 07:25:14
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
海南醫學(2016年8期)2016-06-08 05:43:00
