不同產地太子參蕪菁花葉病毒CP基因的分離與序列差異性分析
2016-05-30 09:33:58匡云波葉煒李金輝于靜靜葉祖云賴鐘雄
熱帶作物學報 2016年10期
匡云波 葉煒 李金輝 于靜靜 葉祖云 賴鐘雄
摘 要 通過對不同產地的8份太子參病葉樣品進行蕪菁花葉病毒(Turnip mosaic virus,TuMV)外殼蛋白(coat protein,CP)基因的RT-PCR檢測,從其中7份樣品中擴增得到特異性目的條帶,對所獲得的13個克隆子序列進行下一步的序列分析。結果顯示:所得CP基因序列均為775 bp,存在36個核苷酸多態性位點,總變異數為37個,部分點突變僅在某產地發現,可能存在重組;各地分離物之間的遺傳分化程度高,遺傳漂變可能是造成遺傳分化的主要原因;在系統進化樹分析結果中,太子參TuMV CP序列均聚類在World B組,并呈現一定的地域特異性。本研究為建立更有效的太子參TuMV檢測方法和針對性制定病毒病防控策略奠定重要基礎。
關鍵詞 太子參;TuMV;CP;RT-PCR;序列差異性分析
中圖分類號 S432.1 文獻標識碼 A
Abstract RT-PCR method was used to detect 8 infected leaf samples collected from Pseudostellaria heterophylla from different producing areas and 7 samples generated an expected fragment, and 13 clones of the coat protein (CP)genes Tunip mesaic virns(TuMV) were selected for sequence analysis. The results showed that the sequences of all CP genes were 775 bp in length. There existed 36 polymorphic sites, 37 mutations and some of them were geographic region-specific sites. Recombination was found in CP genes. The data showed high genetic differentiation among the isolates from Guizhou and Shandong, Jiangsu and Shandong provinces. Genetic drift could be the main cause of genetic differentiation. All the CP genes obtained in this study were classified into World B group in the phylogenetic tree, and indicated somehow geographic region specificity. The results reported in this manuscript laid an important foundation for the establishment of a more efficient molecular detection method and the targeted strategy for the control of TuMV disease in P. heterophylla.
Key words Pseudostellaria heterophylla;TuMV;CP;RT-PCR;Sequence difference analysis
doi 10.3969/j.issn.1000-2561.2016.10.021
太子參(Pseudostellaria heterophylla),又名孩兒參、童參,為石竹科太子參屬多年生草本藥用植物,收載于中華人民共和國藥典中,具有益氣健脾、生津潤肺之功效。太子參在中國華東、華中、華北和西北等地都有分布,主產于福建、貴州、江蘇、山東、湖南等地,栽培歷史已過百年。長期以來,太子參均以塊根進行無性繁殖,導致病毒不斷積累和擴散,太子參種性退化,病毒病害日趨嚴重。據報道,已知侵染太子參的病毒病病原有4種,其中蕪菁花葉病毒(Turnip mosaic virus,TuMV)是侵染太子參的主要病毒病病原,分布于江蘇和山東的各個太子參產區[1]。
TuMV 屬于馬鈴薯 Y 病毒屬(Potyvirus)成員,可通過汁液或蚜蟲傳播,在自然條件下主要靠蚜蟲以非持久性方式傳播。TuMV是RNA病毒,其寄主范圍廣泛,而且大部分為十字花科作物。近年來,關于TuMV的分子進化和遺傳結構的研究已經取得了較大的研究進展[2]。大量研究結果表明,由于地理起源和株系特化等原因,TuMV易產生變異,導致TuMV存在多種株系和致病型[3-4]。TuMV分離物劃分為2個致病型:B型和BR型。B型只侵染蕓苔屬(Brassica),BR型既侵染Brassica,又侵染蘿卜屬(Raphanus)[3]。根據外殼蛋白(coat proein,CP)等基因進行系統發育分析,TuMV分離物大致分為Basal-B、Basal-BR、Asian-BR和World-B組[3-4]。
迄今為止,有關太子參TuMV的鑒定與檢測僅以病毒形態學、生物學和血清學特征為主要依據[1,5-7]。隨著分子生物學的發展,植物病毒的鑒定所需要的信息更全面、更精確,有必要對太子參TuMV進行進一步的分子鑒定。鑒于此本研究擬對來自于不同產地的太子參病毒病病樣進行TuMV CP基因的RT-PCR擴增和克隆,以了解TuMV在太子參產地的發生、分布情況;同時對所得病毒序列進行遺傳多樣性、分子進化等相關的生物信息學分析,以探討地理因素對太子參TuMV遺傳特性的影響,為太子參TuMV病毒的分子檢測、病毒病防控提供有效的理論依據。
1 材料與方法
1.1 材料
供試毒源為2015年3月取自于福建、江蘇、湖南、貴州、山東的太子參病葉,觀察到植株矮小、葉片皺縮、伴有重花葉等典型癥狀,經液氮速凍后于-80 ℃保存備用。
1.2 方法
1.2.1 病葉總RNA的提取 采用Trizol試劑(Invitrogen)按照說明書提取太子參病葉總RNA。采用1%瓊脂糖凝膠電泳檢測RNA的質量,并采用核酸測定儀測定RNA的濃度。
1.2.2 引物設計 根據GenBank數據庫中的TuMV CP的核苷酸序列的保守區域,采用Primer 5.0軟件設計1對簡并引物:上游引物TuMV-F為5′-AGGTGAAAYGCTTGATGCAGGTY-3′,下游引物TuMV-R為5′-GTTHCCATCCARKCCGAACAAAT-3′。引物由北京六合華大基因科技股份有限公司進行合成。
1.2.3 CP基因的克隆 取病葉總RNA 5 μg,采用RevertAid First Strand cDNA Synthesis Kit(Fermentas)進行cDNA第一鏈的合成(10 μL體系)。以反轉錄產物為模板,進行PCR擴增。PCR 反應體系為:cDNA 模板1 μL,上、下游引物各1 μL(10 μmol/L),10×Taq Plus Buffer 2.5 μL,dNTP Mix(2.5 mmol/L)4 μL,Taq Plus(2.5 U/μL)0.25 μL,加H2O至總體積25 μL。PCR擴增條件為:94 ℃預變性4 min;94 ℃變性30 s,53 ℃退火30 s,72 ℃延伸50 s,共35個循環;最后72 ℃延伸10 min。PCR擴增所采用的試劑為2×Taq Plus PCR Master Mix(TianGen)。取3 μL PCR擴增產物進行瓊脂糖(1%)電泳檢測。采用DNA快速純化回收試劑盒(Solarbio)回收目的片段,以pMD-18T(TaKaRa)連接載體,Escherichia coli(DH5α)感受態(TianGen)對回收片段進行連接、克隆和轉化,陽性克隆子送北京六合華大基因科技股份有限公司進行測序。
1.2.4 序列分析 測序獲得的核苷酸序列及其推導的氨基酸序列在NCBI的GenBank數據庫中進行在線Blast,相關序列登錄GenBank。采用DNAMAN軟件進行序列比對(不含引物序列),利用DnaSP v.5.10.1軟件進行DNA多樣性分析、同義與非同義替換估算、基因重組、遺傳分化與基因流分析等,統計核苷酸多樣性(π)、核酸單體型多態性(Hd)、遺傳分化系數(Fst)和基因流(Nm)等參數。使用MEGA 6.06軟件中的Neighbor-Joining(鄰位相連法,NJ)法建立系統發育樹(P-distance法),并應用bootstrap法(重復1 000次)評估系統發育樹。
2 結果與分析
2.1 不同產地太子參TuMV CP基因的克隆
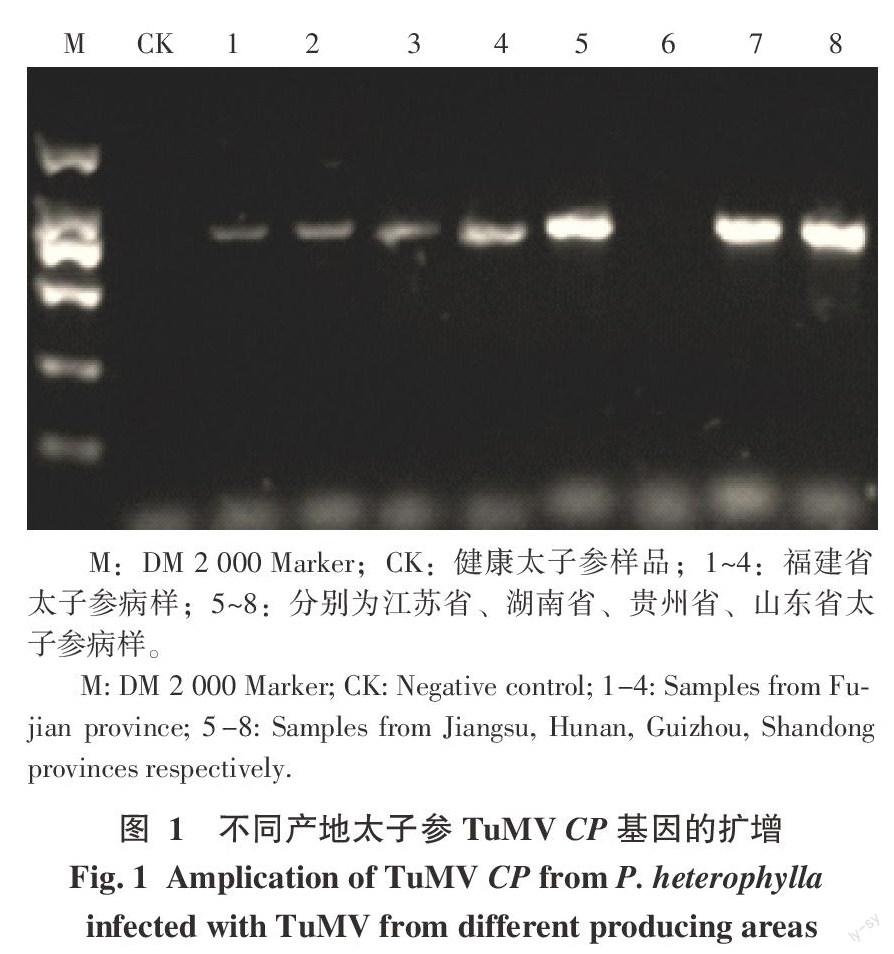
以健康太子參為對照,采用TuMV病毒CP基因的間并引物對8份不同產地的太子參病樣進行PCR擴增,其中除樣品“HN”無擴增條帶外,其他7份樣品均獲得了1條明亮、清晰的大小約為775 bp的特異性條帶,符合預期目的條帶大小(圖1)。對目的條帶進行回收純化、克隆和測序,來自每個樣品的測序克隆數為3~5個。將測序得到的cDNA片段的核苷酸序列及其推導的氨基酸序列在NCBI的GenBank數據庫中在線進行Blast,結果顯示與其它植物TuMV CP基因在核苷酸序列和氨基酸序列上均具有很高的同一性,這表明所得片段為太子參TuMV CP基因的部分序列。
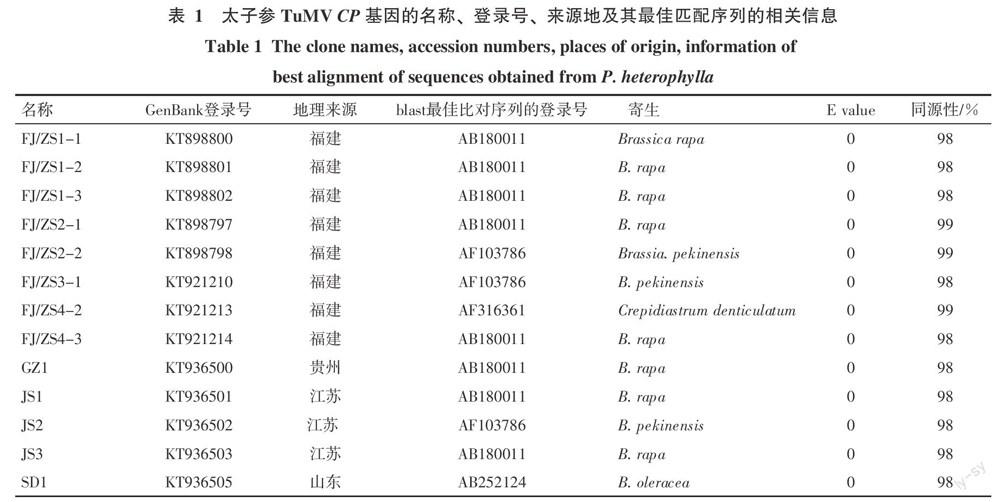
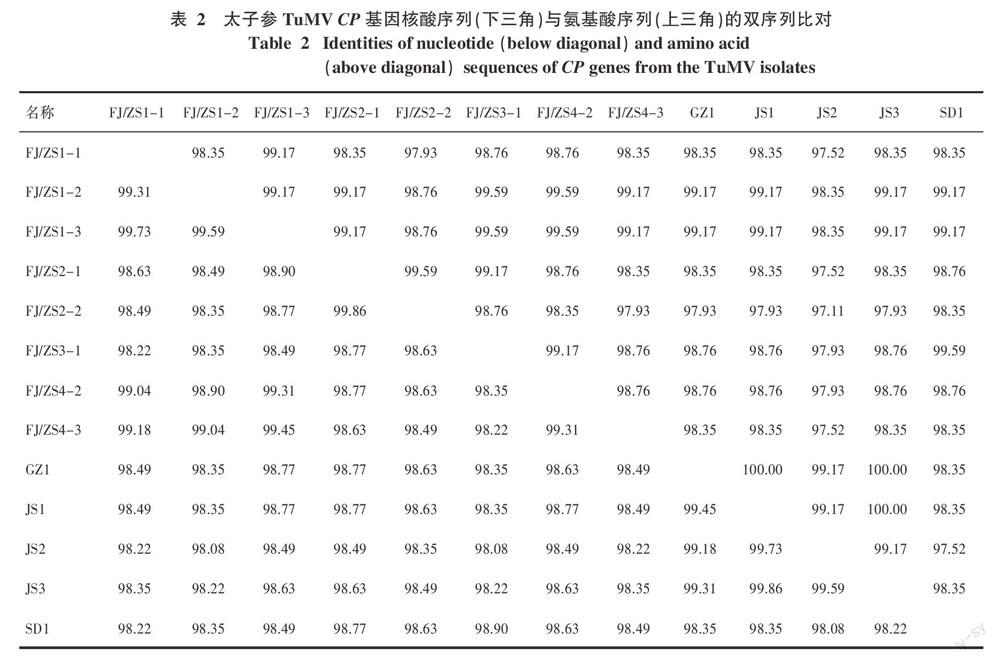
結果表明,7個太子參病樣的CP擴增產物共獲得20個克隆序列,均為775 bp。去掉引物序列后,篩選其中具有序列差異的13個克隆,進行下一步的序列分析,其克隆序列的名稱、GenBank登錄號、地理來源、blast最佳比對序列的登錄號、E value和同源性見表1。太子參TuMV CP序列與NCBI上登錄的黃瓜假還陽參(Crepidiastrum denticulatum,AF316361)分離物等同源性最高,達99%。雙序列比對結果顯示,來自不同樣品的克隆間核苷酸和氨基酸序列的同源性分別在98.08%~99.45%和97.52%~100%;來自同一樣品擴增產物的克隆序列同源性更高,其核苷酸和氨基酸序列同源性分別在99.31%~99.86%和98.35%~100%(表2)。
2.2 不同產地太子參TuMV CP基因的遺傳多樣性分析
將上述太子參TuMV CP基因核苷酸序列導入DnaSP v.5.10.1中。經DNA多樣性分析,結果發現其總單倍型多樣性Hd為1.000,總多態性指數Pi為0.013 14;無序列插入或缺失現象。存在36個多態性位點,其中單一多態位點20個,簡約信息位點16個;總變異個數為37,其中10個同義突變,27個異義突變;部分點突變僅在某區域性產地發現,如江蘇分離物(第64、701個位點)、山東分離物(第165、368、677個位點)、貴州分離物(第419個位點)等。進行同義與非同義替換估算,結果顯示,同義位點數為157.32,非同義位點數為571.68,同義突變多樣性πsyn=0.012 55小于非同義突變多樣性πnonsyn=0.013 30,說明CP基因同義突變的遺傳變異程度小于非同義突變。經重組分析結果發現,在太子參分離物CP基因上發生的最小遺傳重組事件數為4個,分別在以下位點之間:92~125、125~203、263~341、341~686。
將太子參TuMV CP基因序列按地理來源分為福建分離物、貴州分離物、江蘇分離物、山東分離物4組以進行遺傳分化和基因流分析。福建分離物組內病毒CP基因的核苷酸多樣性和單倍體多樣性均為最高。不同分離物間總的遺傳分化系數Fst為0.750 93,總基因流Nm為0.17,暗示各地分離物間遺傳分化程度高,基因交流不頻繁,容易發生遺傳漂變。各地分離物的遺傳分化系數Fst在0.544 74~1.000 00,其中貴州分離物和山東分離物之間的Fst值最高,為1.000 00,說明這2個地區間遺傳分化程度相對較高。
2.3 TUMV CP基因的分子進化分析
應用MEGA 6.06軟件,對所得13條太子參TuMV CP基因序列與已報道的其他寄主植物TuMV CP序列進行系統進化分析,結果見圖2,TuMV病毒分離物可以分為Basal-BR、World-B、Asian-BR、Basal-B 4個組,每個組內均包含了不同寄主植物的TuMV序列,與Ohshima等[3]的研究結果基本一致。太子參TuMV序列與PV0054、KEN1、USA1、GBR7聚類在World-B組,可能屬于B致病類型。不同產地太子參TuMV CP基因序列進一步分成3個小分支,呈現明顯的地域特異性。貴州的GZ1與江蘇的JS1、JS2、JS3聚在一個分支。福建的FJ/ZS1和福建的FJ/ZS4聚在一個分支,而山東的SD1與福建的FJ/ZS2、FJ/ZS3分離物聚在一個分支,與FJ/ZS3親緣關系最近。來自福建的4個分離物之間也存在著一定的遺傳差異,可能是因為來源于不同太子參品種的原因。
3 討論與結論
本研究采用簡并引物對不同產地的8個太子參病樣進行RT-PCR檢測,除來自湖南的樣品“HN”外,其他7個樣品均擴增到了特異目的條帶,說明TuMV分布范圍很廣,福建、貴州、江蘇、山東均存在TuMV侵染太子參的情況,而湖南可能存在其他花葉病病毒侵染。
中國地域遼闊,地形復雜,生境多樣,存在多種TuMV變異類型[8-10]。首先,從系統進化樹分析看,所有TuMV CP序列分為Basal-BR、World-B、Asian-BR、Basal-B 這4個組,每個組內均包含了不同寄主植物的TuMV序列,無明顯的寄主特異性。本研究獲得的不同地理來源的太子參TuMV CP序列均屬于World-B組。據報道,World-B組包含的TuMV病毒大多為B致病型,少數為BR致病型[3]。但宋云枝[8]從山東省3個地市感病的白菜和蘿卜上分離的6個TuMV分離物屬于World-B組,均被鑒定為BR致病型。太子參TuMV的致病類型還有待進一步研究確定。其次,太子參TuMV CP基因序列的聚類具有明顯的地域特異性。雖然不同產地太子參TuMV CP基因序列的同源性達到了98%以上,具有高度的保守性,但仍具有一定的分子變異。本研究結果發現不同產地太子參TuMV CP基因序列存在36個多態性位點,37個突變,其中10個同義突變,27個異義突變。這些突變體都是通過核苷酸堿基轉換或顛換形成的,未出現序列插入或缺失現象,而且有些突變是某產地所特有的,如江蘇分離物、山東分離物、貴州分離物均發現1~3個地區特異性點突變,說明不同產地太子參TuMV間存在一定的地理間隔。
不同地理來源的太子參TuMV分離物CP基因間可能存在遺傳重組。據報道,遺傳重組是馬鈴薯Y病毒屬病毒變異的一個重要因素,TuMV含有多個重組熱點,多數重組發生在P1基因和CI/6K2/VPg基因區域[11],只有極少數重組發生在CP和3′ UTR區域[10,12]。無論在植物還是動物RNA病毒中,重組都是很常見的現象,被認為是病毒毒性改變的決定因素,涉及新的病毒株系的出現[11,13-14]。貴州分離物與山東分離物、江蘇分離物與山東分離物之間遺傳分化程度最高,遺傳漂變可能是造成遺傳分化的主要原因。
本研究通過RT-PCR擴增獲得了不同產地太子參TuMV的CP基因序列,并進一步進行了相關的序列分析,探明了太子參各產地TuMV的發生、分布情況,闡明了太子參TuMV CP基因的分子變異、遺傳分化程度與歸屬類群,為建立太子參TuMV的特異性分子檢測方法和針對性制定病毒病防控策略奠定了重要基礎。
參考文獻
[1] 宋榮浩, 濮祖芹. 太子參(Pseudostellaria heterophylla)病毒病病原鑒定[J]. 上海農業學報, 1991, 7(2): 80-85.
[2] Ohshima K. Plant potyvirus evolution: the survey of the genetic structure of populations[J]. Uirusu, 2012, 62(2): 151-160.
[3] Ohshima K, Yamaguchi Y, Hirota R, et al. Molecular evolution of Turnip mosaic virus: evidence of host adaptation, genetic recombination and geographical spread[J]. J Gen Virol, 2002, 83(6): 1 511-1 521.
[4] Yasaka R, Ohba K, Schwinghamer M W, et al. Phylodynamic evidence of the migration of Turnip mosaic potyvirus from Europe to Australia and New Zealand[J]. J Gen Virol, 2015, 96(3): 701-713.
[5] 陸家云. 藥用植物病害[M]. 北京: 中國農業出版社, 1995.
[6] 黃勇毅,林叢發. 閩東太子參花葉病發病規律調查及防治途徑研究[J]. 寧德師專學報(自然科學版), 2004, 16(1): 65-68.
[7] 劉清琪,陳棣華. 太子參花葉病病原及其防治的初步研究[J]. 中藥材科技, 1983, 6(2): 11-12.
[8] 宋云枝. 蕪菁花葉病毒山東分離物外殼蛋白基因的克隆及序列分析[J]. 中國農業科學, 2005, 38(3): 504-510.
[9] 張成玲. 不同寄主植物上蕪菁花葉病毒外殼蛋白基因克隆及分析[J]. 江蘇農業學報, 2014, 30(1): 37-41.
[10] 田延平. 蕪菁花葉病毒不同分離物3′-cDNA片段的克隆及序列分析[J]. 植物病理學報, 2005, 51(S1): 38-42.
[11] Tan Z Y, Wada Y, Chen J S, et al. Inter- and intralineage recombinants are common in natural populations of Turnip mosaic virus[J]. J Gen Virol, 2004, 85(9): 2 683-2 696.
[12] Tan Z Y, Gibbs A J, Tomitaka Y, et al. Mutations in Turnip mosaic virus genomes that have adapted to Raphanus sativus[J]. J Gen Virol, 2005, 86(Pt 2): 501-510.
[13] Ohshima K, Tomitaka Y, Wood J T, et al. Patterns of recombination in Turnip mosaic virus genomic sequences indicate hotspots of recombination[J]. J Gen Virol, 2007, 88(1): 298-315.
[14] Han Z X, Zhang T T, Xu Q Q, et al. Altered pathogenicity of a tl/CH/LDT3/03 genotype infectious bronchitis coronavirus due to natural recombination in the 5′-17kb region of the genome[J]. Virus Res, 2015, 213(1): 140-148.
