象耳豆根結線蟲Me—mapk1基因的克隆及RNAi分析
2016-05-30 09:33:58魏麗莎子龍海波陳綿才趙志祥
熱帶作物學報 2016年10期
魏麗莎子 龍海波 陳綿才 趙志祥


摘 要 促分裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)是線蟲體內信號轉導的重要組分,在生長發育過程中具有重要作用。本研究從象耳豆根結線蟲中克隆了1個促分裂原活化蛋白激酶基因Me-mapk1 cDNA序列,該序列全長1 369 bp,包含1個1 185 bp的完整開放閱讀框,推導編碼394個氨基酸,蛋白質分子量為45. 39 ku。同源性搜索比對結果發現,該基因編碼的蛋白序列與南方根結線蟲MI-MAPK1具有99%的一致性。通過構建原核表達載體pET-32a-MAPK,在IPTG的誘導下得到70 ku左右的特異融合蛋白(包括大小約26 ku的標簽序列)。利用RNAi技術對象耳豆根結線蟲二齡幼蟲Me-mapk1基因進行沉默,線蟲誘導番茄根結形成的數量顯著減少,推測Me-mapk1基因的下調表達有可能影響象耳豆根結線蟲二齡幼蟲(J2)的生長發育。
關鍵詞 象耳豆根結線蟲;促分裂原活化蛋白激酶;基因克隆;原核表達;RNA干擾
中圖分類號 R532.1 文獻標識碼 A
Abstract Mitogen-activated protein kinases(MAPKs)are important components in signal transduction pathway involving in the process of nematode growth and development. In this study, a MAPK gene(Me-mapk1)from the root-knot nematode Meloidogyne enterolobii was cloned. The Me-mapk1 cDNA sequence consisted of a 1 185 bp open reading frame(ORF)encoding 394 amino acid residues that were franked by a 27 bp 5′-untranslated region(UTR)and a 157 bp 3′-UTR. The protein molecular mass was 45.39 ku. Homology analysis showed that the identity was 99% between Me-mapk1 deduced protein ME-MAPK1 and MI-MAPK1 protein of M. incognita. The recombinant protein ME-MAPK1 with a molecular mass of 70 ku which included tag protein sequence of 26 ku was produced by prokaryotic expression. The number of nematode-induced galls on tomato was significantly reduced after knocking-down Me-mapk1 gene by RNA interference, indicating an important role of the gene to nematode development.
Key words Meloidogyne enterolobii; Mitogen activated protein kinase; Gene cloning; Prokaryotic expression; RNAi
doi 10.3969/j.issn.1000-2561.2016.10.022
象耳豆根結線蟲(Meloidogyne enterolobii)是中國熱帶、亞熱帶地區作物上廣泛發生的一種重要的植物病原線蟲,也是世界上公認的危害最大的根結線蟲種類之一[1-2]。該線蟲最早在海南儋州象耳豆樹上被發現,隨后在歐洲、美洲和非洲等報道發生,造成的產量損失可達65%以上[3-5]。近年來,象耳豆根結線蟲在海南島迅速蔓延擴散,目前已在海南全島發生危害,受害作物包括瓜果蔬菜、南藥、熱帶果樹等多種農作物,并造成重大經濟損失[6]。越來越多的調查結果表明,象耳豆根結線蟲正逐漸取代南方根結線蟲(M. incognita),成為海南熱帶水果和反季節蔬菜上的優勢根結線蟲種類[6-7]。
促分裂原活化蛋白激酶(mitogen-activated protein kinases, MAPK)是一種真核生物體內普遍存在且高度保守的絲氨酸/蘇氨酸蛋白激酶。MAPK與促分裂原活化蛋白激酶激酶(mitogen-activated protein kinase kinase, MAPKK)、促分裂原活化蛋白激酶激酶激酶(mitogen-activated protein kinase kinase kinase,MAPKKK)組成MAPK級聯信號通路,接受外界信號刺激并將信號轉導至細胞內,調控基因表達、細胞增殖與分化、細胞生存與凋亡等多種生命活動[8-9]。MAPK還被證實與多種植物病原物包括真菌、細菌及類病毒的致病性密切相關[10-12]。迄今,有國內外研究人員已從真菌、哺乳動物和植物中分離克隆了多種MAPK基因,如在擬南芥基因組中鑒定出20余種MAPK基因[13-14]。目前植物病原線蟲中關于MAPK的研究較少,僅在南方根結線蟲、甘藍根結線蟲和松材線蟲中克隆了少量地MAPK基因,其具體功能和作用機制均未開展深入研究。本研究從象耳豆根結線蟲中克隆并鑒定出一個新的促分裂原活化蛋白激酶基因Me-mapk1。同時,利用RANi技術對Me-mapk1進行誘導沉默,分析MAPK在象耳豆根結線蟲生長發育過程中的功能地位,旨在為防治該線蟲提供新的理論依據和防治策略。
1 材料與方法
1.1 材料
象耳豆根結線蟲Meloidogyne enterolobii由本實驗室培養,在室內用改進的貝爾曼漏斗法分離得到。培養和接種象耳豆根結線蟲的番茄品種均為“特級大明星”。
1.2 方法
1.2.1 第一鏈cDNA的合成 采用Trizol法提取M. enterolobii總RNA,其RNA沉淀用30 μL無RNase 水溶解。取5 μL總RNA為模板,以oligo dT為引物,用反轉錄酶(TaKaRa PrimeScriptTMⅡ1st Strand cDNA Synthesis Kit)進行反轉錄,合成cDNA第1鏈。
1.2.2 Me-mapk1 cDNA末端的序列擴增 以M. enterolobii總RNA為模板,利用GeneRacerTM RACE 試劑盒(Invitrogen)反轉錄合成獲得帶3′接頭和5′接頭的cDNA第一鏈(RACE模板)。同時,根據實驗室已有的象耳豆根結線蟲cDNA文庫序列信息,設計Me-mapk1基因末端擴增特異引物,結合RACE接頭引物,采用巢式PCR分別擴增基因的5′及3′末端序列。其中,5′末端序列擴增第1輪所用引物為GeneRacer5′ primer和M-S1(表1),第2輪PCR以第1輪PCR產物為模板,所用引物為GeneRacer5′ nested primer和M-S2(表1);3′末端序列的擴增第1輪PCR所用引物為GeneRacer3′primer和M-A1(表1),第2輪PCR所用引物為GeneRacer3′ nested primer和M-A2(表1)。PCR擴增產物經電泳檢測、純化、連接、轉化并送諾賽基因測序。
1.2.3 Me-mapk1原核表達 以第1鏈cDNA為模板,用引物M1-Bam HI和M1-Not I擴增Me-mapk1基因的開放閱讀框區域。PCR 產物經TA克隆,挑取單克隆,搖菌并進行PCR鑒定,將陽性克隆送諾賽公司測序,并進行序列分析。同時,提取陽性克隆和pET-32a質粒,分別經Bam HI和Not I雙酶切后,連接,構建表達重組子pET-32a-MAPK。轉化到感受態細胞BL21(DE3)中,涂板培養。挑取轉化子接種于含氨芐青霉素的培養液中振蕩培養至菌液OD值約為0.6時,加入100 mmol/L IPTG使終濃度為0.8 mmol/L,37 ℃振蕩培養4 h后離心收集菌體,向收集的菌體中加入50 μL蛋白上樣緩沖液,沸水加熱5 min,將樣品取出冷卻至室溫,離心后取8 μL上清于12%分離膠和5%濃縮膠中進行SDS-PAGE電泳分析。以pET-32a空載體誘導表達物作為對照。
1.2.4 Me-mapk1 dsRNA合成 設計在5′端引用T7啟動子序列的dsRNA特異引物M-I1和M-I2(表1),以第一鏈cDNA為模板PCR擴增獲得Me-mapk1特異靶標序列。將獲得的PCR產物純化后,隨后以該片段為模板結合反轉錄酶合成Me-mapk1特異的dsRNA序列。反應體系:DNA模板1 μg,10× T7 Reaction Buffer 2 μL,ATP Solution 2 μL,CTP Solution 2 μL,GTP Solution 2 μL,UTP Solution 2 μL, T7 Enzyme Mix 2 μL,補充Nuclease-free Water至總體積20 μL。具體反應步驟參照MEGAascript RNAi kit(Life Technologies)說明書進行。獲得的dsRNA產物用微量核酸蛋白檢測儀(NanoUV-3000)檢測濃度和質量。
1.2.5 Me-mapk1 RNAi效應分析 RNAi采用活體外dsRNA浸泡二齡幼蟲法進行,略有修改[15]。浸泡體系為:2 mg/mL dsRNA,0.5%間苯二酚,0.05%明膠,3 mmol/L亞精胺,浸泡溶液用1xPBS緩沖液調節。對照處理分2組,一組為不含dsRNA 的1xPBS緩沖液(含0.05%明膠,0.5%間苯二酚,3 mmol/L亞精胺),一組為清水處理。每處理所用新鮮二齡幼蟲數量約20 000條,浸泡時間為24 h,溫度為22 ℃,浸泡過程中保持輕微振動。浸泡結束后用滅菌水反復沖洗蟲體,在清水中恢復2 h后,每處理取約10 000條二齡幼蟲提取總RNA。分別取等量濃度RNA(200 ng/μL)反轉錄合成cDNA,RT-PCR分析沉默效果,所用引物為M-R1和M-R2(表1)。反應體系:cDNA 1 μL,2× pfu PCR MasterMix(TIANGEN)10 μL,上下游引物各1 μL,補充滅菌雙蒸水至總體積20 μL。同時選擇內參基因β-actin為擴增對照,引物為Actin-F1和Actin-R1(表1),反應體系不變。將dsRNA浸泡處理(含對照)的二齡幼蟲接種至生長30 d番茄苗(cv. 特級大明星)根部,每個處理接種300條線蟲,重復8次。接種30 d后收集番茄根系,統計根結數。病情級別和病情指數依據《中華人民共和國國家標準農藥田間藥效實驗準則(二)》中所列方法和公式計算,具體如下:
2 結果與分析
2.1 Me-mapk1 cDNA克隆與序列分析
用RACE技術分別克隆獲得了象耳豆根結線蟲Me-mapk1 cDNA的5′末端和3′末端序列,其長度分別為960、505 bp,拼接后全長序列為1 369 bp。Me-mapk1 cDNA全長序列包含長度為27 bp的5′非編碼區、157 bp的3′非編碼區(包括24個堿基polyA尾巴)和1個1 185 bp的完整開放閱讀框,推導編碼1個含394 aa的蛋白序列。Me-mapk1 cDNA在NCBI GenBank上提交注冊登錄號為KT380882。
象耳豆根結線蟲促分裂原活化蛋白激酶ME-MAPK1蛋白序列由394個氨基酸殘基組成,預測分子量大小為45.39 ku,等電點pI=6.39。BlastP同源性搜索及多序列比對顯示ME-MAPK1與南方根結線蟲M. incognita(ABI96897)促分裂原活化蛋白激酶MI-MPK1有99%的一致性,與甘藍根結線蟲M. artiellia(CAD56894)、松材線蟲Bursaphelenchus xylophilus(ACT46908)及馬來絲線蟲Brugia malayi(XP_001900759)等其它線蟲促分裂原活化蛋白激酶具有86%~93%的一致性(圖1)。蛋白保守結構域分析結果表明,ME-MAPK1第43~331位氨基酸包含11個保守的絲氨酸/蘇氨酸蛋白激酶亞結構域,并且在第VII和第VIII亞結構域間具有胞外信號調節激酶保守的TEY基序序列(圖1)。
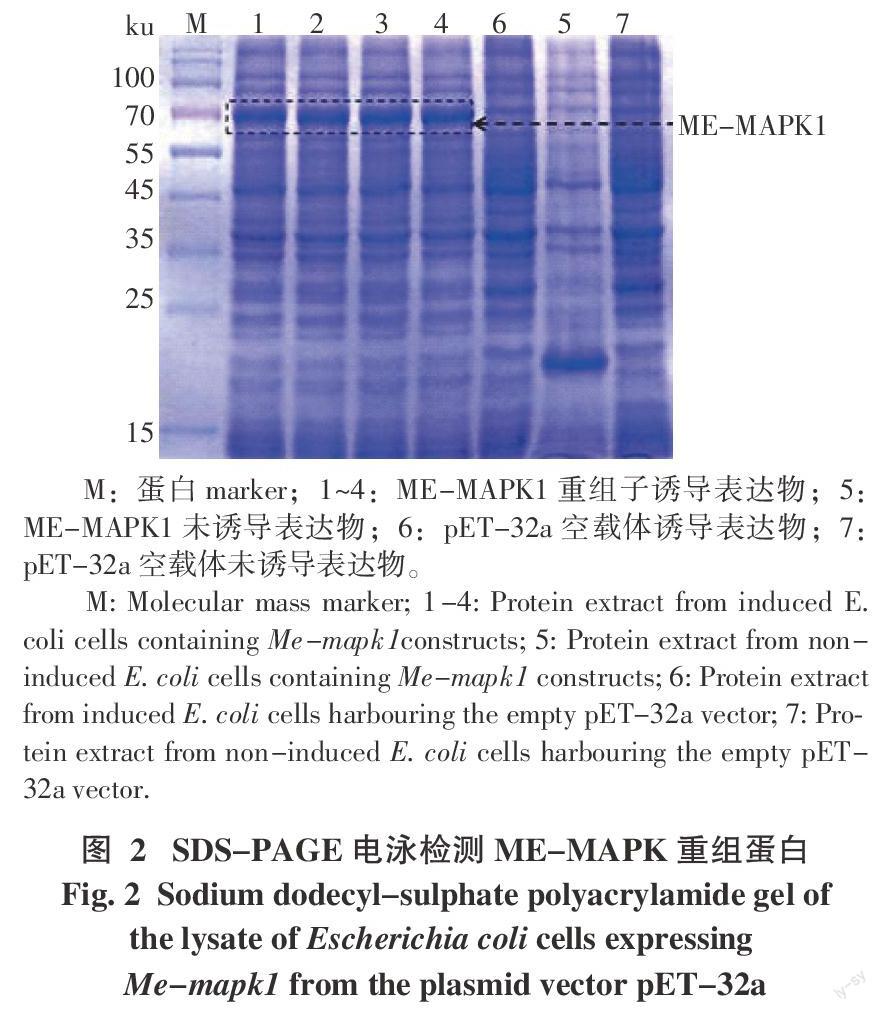
2.2 ME-MAPK1重組蛋白誘導表達
將構建好的重組表達載體質粒pET-32a-MAPK轉入大腸桿菌表達菌株BL21(DE3)進行原核表達。SDS-PAGE電泳檢測結果表明,在IPTG的誘導下ME-MAPK1融合蛋白成功誘導表達,獲得大小約為70 ku的特異條帶,與預測大小相符(包括大小約26 ku的標簽序列);而未經IPTG誘導的重組子及空載體對照無ME-MAPK1融合蛋白條帶(圖2)。
2.3 dsRNA對Me-mapk1基因的沉默效果
通過引入T7啟動子序列的Me-mapk1基因特異引物M-I1/M-I2擴增獲得Me-mapk1靶標片段,隨后在反轉錄酶的作用下合成了長度為332 bp的dsRNA(圖3)。分別提取Me-mapk1 dsRNA、不含dsRNA的緩沖液和清水浸泡處理24 h的二齡幼蟲總RNA,反轉錄RT-PCR擴增Me-mapk1和對照基因β-actin。電泳檢測結果顯示,經Me-mapk1 dsRNA浸泡處理后的二齡幼蟲Me-mapk1轉錄豐度明顯低于無dsRNA 和清水浸泡處理后的Me-mapk1轉錄水平(圖4-A),而非靶標基因β-actin經以上3個處理后其轉錄豐度未發生相對變化(圖4-B)。
2.4 Me-mapk1 RNAi效應表型分析
由表2可知,象耳豆根結線蟲二齡幼蟲經Me-mapk1 dsRNA浸泡處理后導致番茄根結形成數相比對照明顯減少,30 d 后其平均根結形成數為21.5±8.9(平均±SD)。不含dsRNA的PBS緩沖液和清水對照處理形成平均根結數分別為61.8±13.5(平均±SD)和63.1±10.6(平均±SD),對照之間差異不顯著(表2)。同時,二齡幼蟲經Me-mapk1 dsRNA浸泡處理接種后的番茄病情指數為16.7,而不含dsRNA的PBS緩沖液和清水對照處理的番茄病情指數分別為44.4和47.2(表2)。
3 討論與結論
本研究從象耳豆根結線蟲中成功克隆了1個促分裂原活化蛋白激酶基因Me-mapk1,其編碼蛋白ME-MAPK與其它線蟲的MAPK具有80%以上的一致性,屬于典型的絲氨酸/蘇氨酸激酶家族成員。結果表明,各生物體MAPK高度相似,都包含有11個保守的絲氨酸/蘇氨酸亞結構域,并且在第VII和第VIII亞結構域之間的T-loop環上有1個由蘇氨酸(T)、酪氨酸(Y)和X氨基酸組成的TXY基序,是決定MAPK活性的關鍵位點[16-17]。在植物TXY中的X常為E(谷氨酸)或D(天冬氨酸),而動物和酵母中是E、P(脯氨酸)或G(甘氨酸)。多序列分析比對結果顯示象耳豆根結線蟲ME-MAPK與其它線蟲MAPK的TXY基序均為TEY(圖1),表明線蟲MAPK對底物特異性的選擇上可能具有相似性,而本研究通過原核表達獲得的ME-MAPK1融合蛋白也為進一步研究MAPK的生化特性奠定了基礎。
Kamath等[18]利用RNA干擾技術對模式秀麗小桿線蟲mapk基因誘導沉默后,秀麗小桿線蟲出現不育和胚致死等表型,表明MAPK在線蟲的生長發育過程中具有重要作用。而陳國華等[19]用體外合成的南方根結線蟲mapk-1 dsRNA浸泡卵塊,會導致孵化的J2成活率顯著降低,同樣表明MAPK對于植物寄生線蟲的重要性。本研究中,通過體外合成與象耳豆根結線蟲Me-mapk1同源的dsRNA片段并刺激J2吞咽取食,成功誘導了Me-mapk1基因的下調表達,表現為Me-mapk1 mRNA的轉錄豐度降低(圖4)。而經Me-mapk1 dsRNA處理的J2接種后誘導番茄根結形成的數量顯著減少,病情指數下降。由此推測,Me-mapk1基因的下調表達對象耳豆根結線蟲在根內的寄生發育造成了影響。該研究結果表明Me-mapk1可作為抑制象耳豆根結線蟲危害的一個候選靶標基因,具有潛在的應用價值。需要注意的是,利用活體外RNA干擾技術對于基因的沉默具有時效性,不能完全抑制靶標基因的表達,在后期靶標基因的表達水平有可能會恢復到正常水平,從而一定程度上影響表型的分析。因此,需進一步構建能在體內穩定持久表達dsRNA的轉基因植株實現對根結線蟲靶標基因的持續沉默,深入研究MAPK的作用機制和功能地位,為最終提出新的防治象耳豆根結線蟲病害策略奠定理論基礎。
參考文獻
[1] 符美英, 芮 凱, 肖彤斌,等. 海南島象耳豆根結線蟲的種類鑒定及其rDNA-ITS序列分析[J]. 生物安全學報, 2012, 21(1): 79-84.
[2] Castagnone-Sereno P. Meloidogyne enterolobii(M. mayaguensis): profile of an emerging,highly pathogenic, root-knot nematode species[J]. Nematology, 2012, 14(1): 133-138.
[3] Yang B, Eisenback J. Meloidogyne enterolobii n. sp. (Meloidogynidae), a root-knot nematode parasitizing pacara earpod tree in China[J]. Journal of Nematology, 1983, 15(2): 381-391.
[4] Kiewnick S, Dessimoz M, Franck L. Effects of the Mi-1 and the N root-knot nematode-resistance gene on infection and reproduction of Meloidogyne enterolobii on tomato and pepper cultivars[J]. Journal of Nematology, 2009, 41(1): 134-139.
[5] Onkendi E M, Moleleki L N. Distribution and genetic diversity of root-knot nematodes(Meloidogyne spp.)in potatoes from South Africa[J]. Plant Pathology, 2013, 62(6): 1 184-1 192.
[6] 龍海波, 孫燕芳, 白 成,等. 海南省象耳豆根結線蟲的鑒定研究[J]. 熱帶作物學報, 2015, 36(2): 371-376.
[7] 賈本凱, 王會芳, 陳綿才. 海南島茄果類蔬菜根結線蟲種類鑒定[J]. 廣東農業科學, 2012, 7(1): 104-106.
[8] Widmann C, Gibson S, Jarpe M B, et al. Mitogen-activated protein kinase:conservation of a three-kinase module from yeast to human[J]. Physiology Review, 1999, 79(1): 143-180.
[9] Rohila J S, YangY N. Rice mitogen-activated protein kinase gene family and its role in biotic and abiotic stress response[J]. Journal of Integrative Plant Biology, 2007, 49(4): 751- 759.
[10] 范永山, 劉穎超, 谷守芹, 等. 植物病原真菌的MAPK基因及其功能[J]. 微生物學報, 2004, 44(3): 547-551.
[11] Hamel L P, Nicole M C, Duplessis S, et al. Mitogen-activated protein kinase signaling in plant-interacting fungi:distinct messages from conserved messengers[J]. Plant Cell,2012, 24(7): 1 327-1 351.
[12] 彭靜靜. 植物病原真菌中MAPK級聯通路研究進展[J]. 江蘇農業科學, 2014, 42(1): 11-15.
[13] Ichimura K, Mizoguchi T, Irie K, et al. Isolation of ATMEKK1(a MAP kinase kinase kinase): interacting proteins and analysis of a MAP kinase cascade in Arabidopsis[J]. Biochemical and Biophysical Research Communications, 1998, 253(3): 532-543.
[14] Ren D T, Liu Y D, Yang K Y, et al. A fungal-responsive MAPK cascade regulates phytoalexin biosynthesis in Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(12): 5 638-5 643.
[15] Chen Q, Rehman S, Smant G, et al. Functional analysis of pathogenicity proteins of the potato cyst nematode Globodera rostochiensis using RNAi[J]. Molecular Plant-Microbe Interactions, 2005, 18(3): 621-625.
[16] Bardwell A J, Abdollahi M, Bardwell L. Docking sites on mitogen-activated protein kinase(MAPK) kinases, MAPK Phosphatases and the Elk-1 transcription factor compete for MAPK binding and are crucial for enzymic activity[J]. Biochemical Journal, 2003, 370(5): 1 077-1 085.
[17] Colcombet J, Hirt H. Arabidopsis MAPKs: A complex signaling network involved in multiple biological processes[J]. Biochemical Journal, 2008, 413(1): 217-226.
[18] Kamath R S, Fraser A G, Ahringer J, et al. Systematic functional analysis of the Caenorhabditis elegans genome using RNAi[J]. Nature, 2003, 421(1): 231-237.
[19] 陳國華, 肖 羅, 張雙慶,等. 南根結線蟲促分裂原活化蛋白激酶(MAPK)基因的RNAi效應分析[J]. 植物病理學報, 2008, 38(3): 509-531.
