腐片鐮刀菌醇提取物與苯并噻二唑誘導水稻對抗稻瘟病相關酶活性的比較
2016-05-30 10:48:04王紅楊濤楊鎮肇瑩陳珣曹君李躍馬曉穎肖軍
南方農業學報 2016年1期
王紅 楊濤 楊鎮 肇瑩 陳珣 曹君 李躍 馬曉穎 肖軍

摘要:【目的】探討微生物源和化學農藥對水稻稻瘟病相關抗性酶系活性的影響,為研發安全、綠色的生物農藥提供理論依據。【方法】于水稻3葉1心期進行微生物源農藥[腐片鐮刀菌(Fusarium solani)次生代謝產物,WS]和化學農藥苯并噻二唑(BTH)誘抗處理,以噴灑蒸餾水為對照(空白對照CK1和陽性對照CK2),采用外接稻瘟病病菌法測定相關抗性酶系活性及植株抗病性。【結果】接種稻瘟病菌后0~9 d,WS處理的苯丙氨酸解氨酶(PAL)、過氧化物酶(POD)和多酚氧化酶(PPO)活性及BTH處理的PAL和POD活性均高于CK2,BTH處理對提高PAL活性的效果優于WS處理,但WS對提高PPO活性的持續性較優;兩種誘抗劑前期(0~3 d)均可提高葉片過氧化氫酶(CAT)活性,但后期CAT活性均低于CK2;接種稻瘟病菌后各處理的肉桂醇脫氫酶(CAD)活性均無明顯變化。WS和BTH誘導處理后,水稻病株率、病斑數及病情指數均降低,其中WS處理的發病情況極顯著低于CK(P<0.01),誘抗效果達56.21%,比BTH處理高14.78%(絕對值)。【結論】WS和BTH對誘導水稻抗稻瘟病均有一定效果,且WS的誘抗效果整體上優于BTH。
關鍵詞: 腐片鐮刀菌;苯并噻二唑;稻瘟病;誘導抗性;PAL;POD;CAD;PPO;CAT
中圖分類號: S511.01;S435.11 文獻標志碼:A 文章編號:2095-1191(2016)01-0024-05
0 引言
【研究意義】稻瘟病是水稻生產中的常發性病害,每隔幾年就有一次較大的流行。我國稻瘟病發生面積約在380萬ha以上,其損失一般為10%~20%,大流行年份有時在50%以上,甚至顆粒無收(趙先麗等,2014)。稻瘟病是由稻瘟病菌(Magnaporde grisea)引起的水稻真菌性病害,病原小種種類復雜,變異快,僅通過篩選新的抗病品種并不能有效防治稻瘟病肆虐。目前,防治水稻稻瘟病的方法除了培育抗性品種外,化學防治、生物防治及合理的栽培措施等也是常用的防治方法(溫小紅等,2013)。其中,生物防治因其藥害低、不易產生抗藥性、生產原料廣泛、使用安全放心、環境兼容性好及選擇性強等優點,在綜合治理水稻病害中發揮著越來越重要的作用(李海靜,2014)。生物農藥最能體現生物防治的特點,其作用效果一般是多因素、多基因共同作用的結果,不宜使靶標生物產生抗藥性。因此,通過探討生物農藥對水稻稻瘟病抗病酶系的影響,對進一步研究生物農藥在水稻抗病上的應用具有重要意義。【前人研究進展】目前,在稻瘟病的防治方面,一般以化學藥劑為主,如葛秀春等(2002)研究了苯并噻二唑(BTH)誘發水稻對稻瘟病抗性酶系活性的變化,結果表明,BTH誘導后,肉桂醇脫氫酶(CAD)、脂氧合酶(LOX)、過氧化物酶(POD)活性大幅度升高,但苯丙氨酸解氨酶(PAL)活性無變化;徐沛東等(2014)研究表明,利用8%烯丙苯噻唑顆粒劑處理水稻幼苗后,防御酶系活性均高于對照,田間藥效試驗對稻瘟病防治效果為66.26%。近年來,生物農藥、尤其是微生物源農藥在防治稻瘟病上的研究受到廣泛關注。遲莉(2014)篩選出2株抑菌能力較強的枯草芽孢桿菌,其發酵液噴施水稻葉片后,POD和多酚氧化酶(PPO)活性增加4~6倍,PAL活性增加25%,小區試驗防治效果達54%;肖軍等(2015)利用內生菌醇提取物誘導水稻抗性,結果表明,PAL活性增加30.28%,PPO和幾丁質酶活性分別增加1.18和1.70倍,稻瘟病情指數降低12.89。【本研究切入點】本課題組前期研究從遼寧西北地區采集的野生蘆葦根系中分離得到一株植物內生真菌[腐片鐮刀菌(Fusarium solani)],其次生代謝產物對稻瘟病菌的抑制率為38.75%,對盆栽水稻稻瘟病的誘導抗病效果為76.82%(陳珣等,2012),并確定次生代謝產物濃度為50 ng/mL、噴霧處理時的誘導抗病效果最佳(肖軍等,2015)。本研究擬在前期研究的基礎上進一步對比分析該次生代謝產物與化學農藥對水稻稻瘟病防御酶系的作用效果。【擬解決的關鍵問題】于水稻3葉1心期進行微生物源農藥(野生蘆葦內生真菌Fusarium solani醇提取物,WS)和化學農藥苯并噻二唑(BTH)誘抗處理,以噴灑蒸餾水為對照,采用外接稻瘟病病菌法測定相關抗性酶系活性及植株抗病性,對比微生物源和化學農藥對水稻稻瘟病相關抗性酶系活性的影響,為研發安全、綠色的生物農藥提供理論依據。
1 材料與方法
1. 1 試驗材料
供試水稻品種為遼星1號;病原菌為稻瘟病生理小種703,由沈陽農業大學生物技術研究室提供;供試菌株為Fusarium solani,由遼寧省農業科學院微生物工程中心保存;供試農藥為BTH(先正達公司研發,諾華公司用歐洲商標BION或美國商標Actigard生產)。
1. 2 試驗方法
1. 2. 1 病菌孢子懸浮液的制備 稻瘟病菌703接種在番茄燕麥培養基上,28 ℃培養15 d,菌絲長滿全皿時用棉簽刮去菌絲,將培養皿放在濕潤處,2 d后收集孢子,并制備成濃度為1.0×105個/mL(徐沛東等,2014)的孢子懸浮液備用。
1. 2. 2 次生代謝產物(WS)—菌絲醇提取物的制備 WS為野生蘆葦內生真菌Fusarium solani的菌絲醇提取物,參照陳珣等(2012)的方法進行提取,使用濃度為50 ng/mL(肖軍等,2015)。
1. 2. 3 抗病誘導水稻幼苗 將水稻種子用0.1%HgCl消毒15 min后于28 ℃浸種,催芽4 d,播種在塑料盆內(100株/盆),共播種4盆,在25 ℃的植物生長箱內培養。稻苗長至3葉1心時,1盆水稻幼苗噴灑50 mL的WS(50 ng/mL,含0.1%吐溫-20),1盆噴灑50 mL的 BTH(136 μg/mL,含0.1%吐溫-20),另外2盆各噴灑50 mL蒸餾水(含0.1%吐溫-20)作為空白對照(CK1)和陽性對照(CK2)。誘導處理3 d后,噴霧接種稻瘟病菌分生孢子(CK1不接種),按葛秀春等(2002)的方法保溫保濕培養。分別于接種稻瘟病菌0、3、6和9 d后取樣測定水稻幼苗抗病相關酶系活性,每個指標稱取葉片0.4 g左右(4片葉片),3次重復。
1. 3 測定項目及方法
1. 3. 1 抗病相關酶系活性測定 PAL、PPO活性參照徐沛東等(2014)的方法測定;CAD活性參照葛秀春等(2002)的方法測定;POD活性參照張穗等(2003)的方法測定;過氧化氫酶(CAT)活性參照唐勇軍等(2011)的方法測定。
1. 3. 2 誘導水稻抗稻瘟病效果檢測 水稻幼苗培養和接種方法同1.2.3,接種8 d后調查每片葉片的發病情況,按照GB/T 15790-2009記錄發病級數、總株數和發病指數,計算病情指數。
1. 4 統計分析
利用Excel 2010對不同處理條件下抗性酶系變化趨勢進行分析,采用SPSS 13.0對抗稻瘟病效果進行多重比較。
2 結果與分析
2. 1 不同處理對水稻葉片PAL活性的影響
由圖1可以看出,在水稻3葉1心期(0 d),不同處理間葉片PAL活性基本穩定,為74.17~75.59 U/g·min。接種稻瘟病菌孢子后3 d,WS和CK2處理的葉片PAL活性明顯降低,分別比0 d時降低10.87%和15.84%;而BTH和CK1處理的葉片PAL活性無明顯變化。隨著接種時間的延長,WS處理的葉片PAL活性有所回升,BTH處理的葉片PAL活性略有降低,CK2處理的葉片PAL活性持續降低,CK1處理的葉片PAL活性先降低后升高。接種后6~9 d,4個處理的葉片PAL活性表現為BTH>CK1>WS>CK2。表明誘抗劑對接種后的水稻葉片PAL活性具有明顯的系統誘導作用,且BTH的誘導效果優于WS。
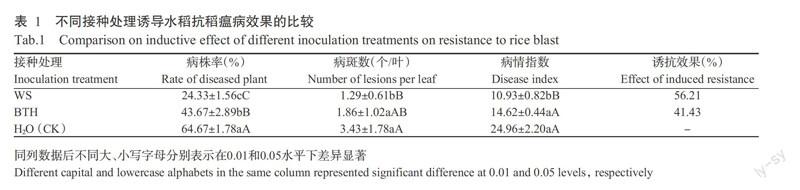
2. 2 不同處理對水稻葉片POD活性的影響
由圖2可以看出,接種稻瘟病菌孢子后3 d,WS、BTH和CK2處理的水稻葉片POD活性均明顯升高,分別比0 d時升高174.85%、95.41%和72.61%,CK1處理的POD活性變化不明顯。隨著接種時間的延長,WS處理的葉片POD活性先降低后升高,其他處理則持續升高,至第9 d時,WS和BTH處理的葉片POD活性分別為78.16和79.16 U/g·min,均高于CK1和CK2處理。表明使用誘抗劑,尤其是WS可使水稻幼苗在受到稻瘟病菌侵染初期時的POD活性迅速增加,在一定程度上使葉片抗逆反應提前,有助于保護植物免受病原菌進一步侵染,雖然CK2處理的活性也有所升高,但其增幅明顯低于誘抗處理。
2. 3 不同處理對水稻葉片CAD活性的影響
接種稻瘟病菌孢子后0~9 d,4個處理的水稻葉片CAD活性無明顯變化,分別為1.90~1.96、1.96~2.14、1.95~2.02和1.93~2.08 U/g·min。說明誘抗處理對水稻葉片的CAD活性影響不明顯。
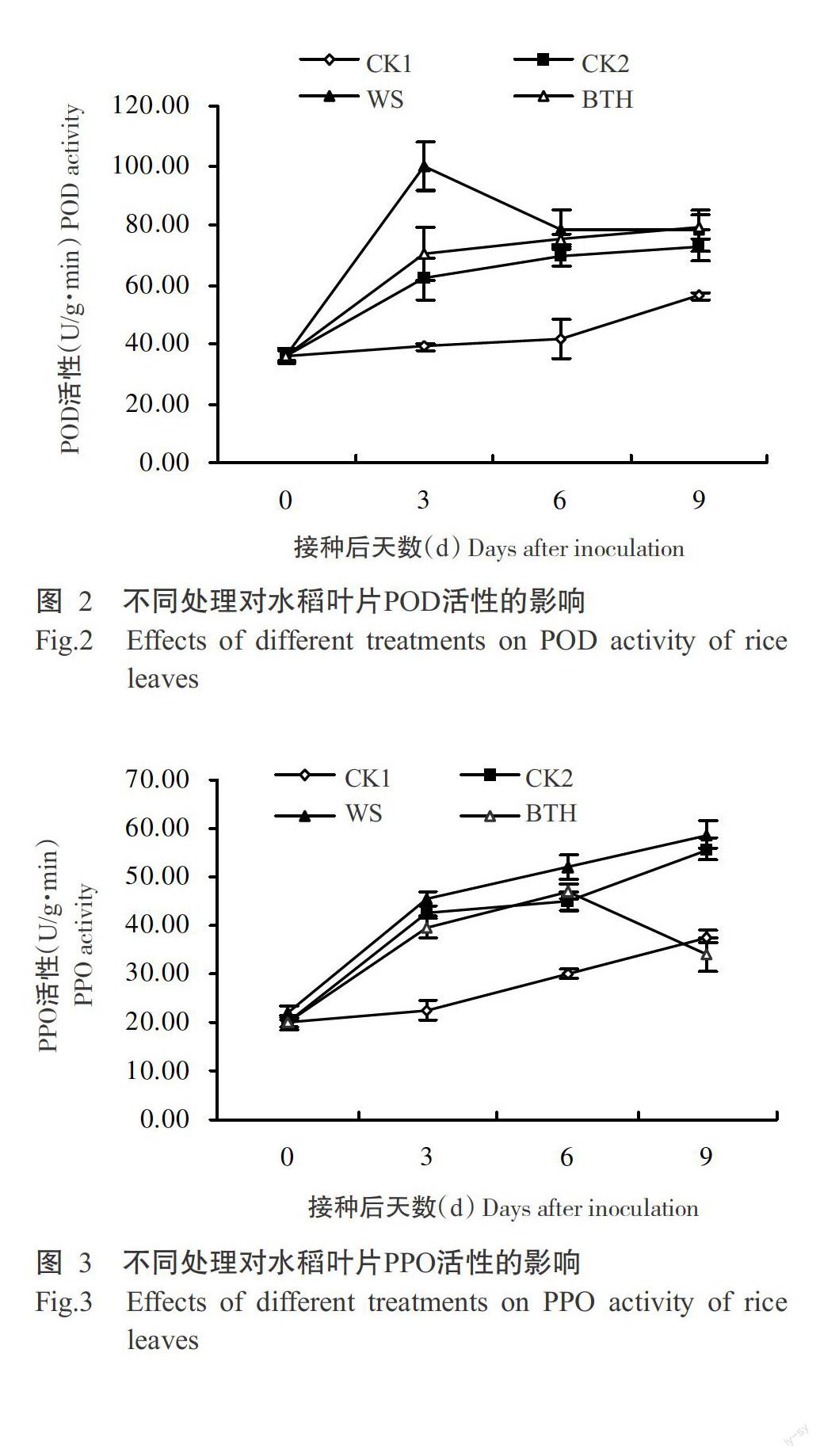
2. 4 不同處理對水稻葉片PPO活性的影響
由圖3可以看出,接種稻瘟病菌孢子后3 d,WS、BTH和CK2處理的水稻葉片PPO活性均明顯升高,分別為45.42、39.68和42.62 U/g·min,CK1處理的PPO活性變化不明顯。隨著接種時間的延長,WS、CK1和CK2處理的PPO活性持續上升,BTH處理的PPO活性先升高后降低,至接種后9 d,WS處理的POD活性最高,為58.67 U/g·min,BTH處理的PPO活性最低,為34.01 U/g·min。表明WS處理使水稻葉片受到病原菌侵染時PPO活性持續增加,BTH處理雖在前期也能促使水稻葉片PPO活性增加,但后期PPO活性下降較快,誘抗效果不及WS。
2. 5 不同處理對水稻葉片CAT活性的影響
由圖4可以看出,接種后3 d,CK2處理的水稻葉片CAT活性大幅降低,隨著自身防御機制的逐漸啟動,CAT活性逐漸升高,第6 d時達到峰值,第9 d時略有下降,但仍高于其他處理。接種后0~6 d時,WS和BTH處理的CAT活性均呈先升高后降低的變化趨勢,且BTH處理的活性均高于WS處理;但至第9 d時,BTH處理后的CAT活性大幅度降低,降至126.72 U/g·min,在4個處理中活性最低,而WS處理的CAT活性仍持續升高,為153.91 U/g·min,表明WS的誘導持久性優于BTH。同時可以看出,在受到病原菌侵染初期,誘抗劑有助于提高水稻葉片的CAT活性,使植物免受病原菌的侵染,但隨著侵染時間的延長,誘抗劑提高CAT活性的能力弱于植物自身的防御功能。
2. 6 不同接種處理誘導水稻抗稻瘟病效果的比較
分別以WS、BTH和H2O(CK)進行誘導水稻抗稻瘟病試驗,由表1結果可知,WS和BTH誘導處理后,水稻病株率、病斑數及病情指數均降低,其中WS處理的發病情況極顯著低于CK(P<0.01),誘抗效果達56.21%,比BTH處理高14.78%(絕對值)。
3 討論
壞死型病原菌侵染常能引起植物隨后抗病性的系統性誘導,被稱為系統獲得抗性(Systemic acquired resintance,SAR)(Ryals et al.,1996)。近年來,有研究證明BTH可誘導多種植物產生系統抗性,如水稻抗稻瘟病(葛秀春等,2002)、番茄抗灰霉病(Azami-Sardooei et al.,2013)、辣椒抗黃色葉病毒(Trejo-Saavedra et al.,2013)等。本研究通過比較已知能夠誘導多種植物產生SAR的化學誘抗劑BTH和自主分離的Fusarium solani次生代謝產物WS對水稻抗稻瘟病酶系的影響,結果表明,BTH和WS在不同程度上對水稻抗病酶系的活性均有促進作用,均通過提高酶類活性而提高水稻抗病性,但其作用效果不同,在稻瘟病菌脅迫下,BTH對PAL活性的提高優于WS,WS前期對POD活性的提高優于BTH,兩者前期對PPO和CAT活性的影響趨勢基本相同,但BTH的持續性不佳。WS和BTH在水稻抗稻瘟病作用上均有一定效果,但WS的誘抗效果優于BTH。同時,在水稻接種培養階段,觀察到噴施BTH的水稻苗的生長速度低于其他處理,因此該化學誘抗劑可能不適合在苗期使用。WS為微生物源誘抗劑,屬生物農藥,可最大程度地降低化學農藥的使用,減輕環境污染。
有研究表明,SAR表現過程中激發植物防衛基因表達及反應機制啟動,其中編碼PAL的基因是一個很重要的防衛基因(遲莉,2014)。植物遭遇病原物侵染時會合成許多新的物質, 包括植保素、木質素、酚類化合物及病程相關蛋白等, 這些物質大多需要通過苯丙烷類代謝途徑合成,而PAL是苯丙烷類代謝途徑的限速酶。黎軍英等(2004)研究表明,通過使水稻植株PAL基因表達量提高、PAL活性升高,可達到水稻抗稻瘟病的目的,與本研究中使用WS和BTH誘導后,水稻葉片PAL活性升高,有助于PAL基因的表達,促進系統獲得抗性建立的結果一致。另一方面,防御反應機制主要涉及細胞壁木質化,POD和CAD均是植物體內重要的防御類氧化酶,也是木質素合成的關鍵酶。本研究結果表明,誘抗劑可提高水稻葉片POD活性,使其參與防御反應機制,但CAD活性幾乎沒有變化。這與葛秀春等(2002)研究發現BTH可使水稻葉片CAD活性在接種后2 d內顯著增高的結果不同,可能是由于本研究中所用誘抗劑的濃度較低,不足以激起CAT活性變化。
PPO是一種含銅的氧化酶,在植物體內催化各種酶類物質氧化形成相應的醌和木質素前體。CAT是清除H2O2的主要酶類,與POD協作清除體內的超氧自由基。本研究結果表明,WS和BTH處理均可使水稻葉片PPO和CAT活性升高,有助于植物產生抗病性,且PPO的變化趨勢與Zhong等(2010)研究表明接種稻瘟病菌24 h后,藥劑噴施處理PPO活性升高的趨勢基本一致。本研究中,WS的持續性優于BTH,但兩者提高CAT活性的能力弱于植物自身的防御功能。
本研究就誘抗劑對水稻稻瘟病抗性酶系的影響及病情指數進行了研究,今后尚需進一步研究微生物源誘抗劑WS抗稻瘟病的機理及其作為生物農藥應用于田間試驗時存在的劑型、合適劑量及穩定性等一系列問題。
4 結論
本研究結果表明,不同誘抗劑誘導水稻抗稻瘟病的效果存在差異,微生物源誘抗劑WS的作用效果整體上優于化學藥劑BTH。
參考文獻:
陳珣,楊鎮,肖軍,龔娜,丑靜,王娜,王紅,肇瑩,楊濤. 2012. 內生真菌No051的鑒定及其代謝產物對稻瘟病的誘導抗病研究[J]. 廣東農業科學,(24):79-81.
Chen X,Yang Z,Xiao J,Gong N,Chou J,Wang N,Wang H,Zhao Y,Yang T. 2012. Indentification of endophytic fungus No051 and its metabolites on induced resistances of rice blast[J]. Guangdong Agricultural Sciences,(24):79-81.
遲莉. 2014. 水稻稻瘟病拮抗菌篩選及施用后對水稻植株的影響[D]. 北京:中國農業科學院.
Chi L. 2014. Effect on rice plants antimicrobial rice blast screening and application of antagonistic[D]. Beijing:Chinese Academy of Agricultural Sciences.
葛秀春,宋鳳鳴,陳永葉,鄭重. 2002. 苯并噻二唑誘發水稻對稻瘟病抗性中防衛相關酶活性的變化[J]. 中國水稻科學,16(2):171-175.
Ge X C,Song F M,Chen Y Y,Zheng Z. 2002. Changes in activities of defense-related enzymes in rice resistance induced by benzothiadiazole to blast fungus(Magnaporthe grisea)[J]. Chinese Journal of Rice Science,16(2):171-175.
李海靜. 2014. 烯丙苯噻唑對水稻的安全性和對稻瘟病的防治效果研究[D]. 大慶:黑龍江八一農墾大學.
Li H J. 2014. Study on the safety evaluation on rice and the control effect on rice blast by probenazole[D]. Daqing:Heilongjiang Bayi Agricultural University.
黎軍英,郭澤建,張炳欣. 2004. 轉PAL基因水稻抗稻瘟病性和過氧化物酶活性的研究[J]. 中國水稻科學,18(4):303-308.
Li J Y,Guo Z J,Zhang B X. 2004. Rice blast resistance and perxidase activity od rice transformed with PAL gene[J]. Chinese Journal of Rice Science,18(4):303-308.
唐勇軍,彭麗莎,鄒俊,田云,盧向陽. 2011. 幾丁質酶產生菌發酵液對水稻生長及防御酶活性的影響[J]. 熱帶作物學報,32(3):393-397.
Tang Y J,Peng L S,Zou J,Tian Y,Lu X Y. 2011. The effects of the fermentation liquor from the chitinase producing strains on rice growth and the activities of defensive enzymes[J]. Chinese Journal of Tropical Crops,32(3):393-397.
溫小紅,謝明杰,姜健,楊寶靈,邵艷龍,何偉,劉麗,趙毅. 2013. 水稻稻瘟病防治方法研究進展[J]. 中國農學通報,29(3):190-195.
Wen X H,Xie M J,Jiang J,Yang B L,Shao Y L,He W,Liu L,Zhao Y. 2013. Advances in research on control method of rice blast[J]. Chinese Agricultural Science Bulletin,29(3):190-195.
肖軍,楊鎮,陳珣,王紅,肇瑩,劉國麗,楊濤. 2015. 內生菌醇提取物對水稻抗稻瘟病的誘導作用[J]. 中國生物防治學報,31(3):433-438.
Xiao J,Yang Z,Chen X,Wang H,Zhao Y,Liu G L,Yang T. 2015. Induced resistance against blast disease of rice by ethanol extract from endophytesto[J]. Chinese Journal of Biological Control,31(3):433-438.
徐沛東,常冬冬,蘭波,楊迎青,李湘民. 2014. 烯丙苯噻唑對水稻主要防御酶活性的影響及其對稻瘟病的防治效果[J]. 華中農業大學學報,33(4):60-65.
Xu P D,Chang D D,Lan B,Yang Y Q,Li X M. 2014. Influence of probenazol on main defense enzymes in rice plants and its control efficacy against rice blast[J]. Journal of Huazhong Agricultural University,33(4):60-65.
張穗,趙清華,唐文華,Wolf G A. 2003. 井岡霉素A對水稻抗性相關酶活性的影響[J]. 植物保護學報,30(2):177-179.
Zhang S,Zhao Q H,Tang W H,Wolf G A. 2003. The influence of Jinggangmycin A on the activities of resistance related enzymes in rice[J]. Journal of Plant Protection,30(2):177-179.
趙先麗,劉志恒,紀瑞鵬,張淑杰,蔡福,程海濤,劉明,張玉書. 2014. 遼寧水稻主產區稻瘟病發生特征分析[J]. 氣象與環境學報,30(2):88-92.
Zhao X L,Liu Z H,Ji R P,Zhang S J,Cai F,Cheng H T,Liu M,Zhang Y S. 2014. Characteristic of occurrence of rice blast in major rice producing areas of Liaoning province[J]. Journal of Meteorology and Environment,30(2):88-92.
Azami-Sardooei Z,Seifi H S,Vleesschauwer D D,H■fte M. 2013. Benzothiadiazole(BTH)-induced resistance against Botrytis cinerea is inversely correlated with vegetative and generative growth in bean and cucumber,but not in tomato[J]. Auatralasian Plant Pathology,42(4):485-490.
Ryals J A,Neuenschwander U H,Willits M G,Molina A,Steiner H Y,Hunt M D. 1996. Systemic acquired resistance[J]. Plant Cell,8(10):1809-1819.
Trejo-Saavedra D L,Garcia-Neria M A,Rivera-Bustamante R F. 2013. Benzothiadiazole(BTH) induces resistance to pepper golden mosaic virus(PepGMV)in pepper(Capsicum annuum L.)[J]. Biological Research,46(4):333-340.
Zhong L J,Zhao X H,Zhang Q H,Xu C,Zhu H L. 2010. Rice resistance against blast induced by tetramycin[J]. Plant Disease and Pests,1(3):6-8.
(責任編輯 王 暉)
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中老年保健(2021年12期)2021-11-30 02:58:01
軍事文摘(2021年22期)2021-11-26 00:43:51
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
文苑(2020年6期)2020-06-22 08:41:52
科技傳播(2019年22期)2020-01-14 03:06:54
文苑(2019年22期)2019-12-07 05:29:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
攝影之友(影像視覺)(2019年2期)2019-03-05 08:27:14
