犬瘟熱病毒疫苗免疫逃逸株的血凝素基因變異研究
2016-06-03 01:43:08隋曉梅王怡男
華北農(nóng)學(xué)報 2016年2期
黃 娟,隋曉梅,王怡男,單 虎
(青島農(nóng)業(yè)大學(xué) 動物科技學(xué)院,山東省高校預(yù)防獸醫(yī)學(xué)重點實驗室,山東 青島 266109)
?
犬瘟熱病毒疫苗免疫逃逸株的血凝素基因變異研究
黃娟,隋曉梅,王怡男,單虎
(青島農(nóng)業(yè)大學(xué) 動物科技學(xué)院,山東省高校預(yù)防獸醫(yī)學(xué)重點實驗室,山東 青島266109)
摘要:為了弄清是否有新基因型犬瘟熱病毒出現(xiàn),對山東省不同地區(qū)的免疫水貂、狐貍和犬進行了犬瘟熱病毒檢測,擴增了病毒核衣殼基因和血凝素基因(H),并對13 個犬瘟熱病毒山東株(8株貂源,2株狐貍源,3株犬源)血凝素基因進行了測序。結(jié)果表明,13個犬瘟熱病毒山東株之間H基因核苷酸同源性為99.1%~100.0%,均為Asia-1型,這也是我國目前水貂犬瘟熱病毒唯一的基因型。與2012年之前的犬瘟熱病毒中國分離株及疫苗株相比,13個毒株H基因有4個氨基酸的一致性變異,即 P200S、M263I、I542N和Y549H,N-糖基化位點比Onderstepoort 株多6或7個,比CDV3株多3或4個。抗原性分析表明,與疫苗株相比,13株山東株在540-549位氨基酸出現(xiàn)一個新的抗原區(qū)域,210-230,327-347位的抗原區(qū)域也與疫苗株不同,這些變異可能是我國貂狐犬瘟熱免疫失敗的原因。
關(guān)鍵詞:犬瘟熱病毒;血凝素基因;免疫失敗
犬瘟熱病毒(Caninedistempervirus,CDV)是高度傳染性的病原,可感染許多食肉目動物,尤其是犬科動物。過去人們認為貓科動物對CDV抵抗力最強,但美國曾報道過貓科動物的致死性感染[1]。CDV被認為是非人靈長類動物和瀕危動物如東北虎的重要威脅[2-3],引起世界范圍內(nèi)人們對CDV可能引起人類感染和危害野生動物保護的擔(dān)憂。弱毒疫苗廣泛應(yīng)用于犬和其他動物來預(yù)防CDV感染,然而,免疫犬和水貂仍發(fā)生CDV感染是和疾病已見幾例報道[4-5],表明定期監(jiān)測和檢測CDV是非常重要和必要的。
CDV屬于副黏病毒科麻疹病毒屬,其負股單鏈基因組RNA編碼6個結(jié)構(gòu)蛋白:核衣殼蛋白(N)、磷蛋白(P)、膜相關(guān)基質(zhì)蛋白(M)、融合蛋白(F)、血凝素(H)和聚合酶(L)[6]。受體結(jié)合蛋白H是CDV主要的保護性抗原蛋白,具有相當(dāng)大的遺傳多樣性,因此,被廣泛用于分子流行病學(xué)調(diào)查研究[5,7-9]。基于H基因序列的進化樹分析表明CDV 毒株可以分為13個基因型:America-1(或疫苗型)、America-2、Asia-1、Asia-2、Asia-3、Asia-4、Rockborn-like、Europe Wildlife、Arctic/Arctic-like、South Africa、South America-1/Europe、South America-2 和 South America-3[7-11]。
在我國,關(guān)于CDV遺傳多樣性的報道多是基于對犬源毒株的研究[5,10-13],而水貂和狐貍源毒株以及免疫動物CDV的基因特征相關(guān)信息較少。2012年以來,山東省免疫水貂和狐貍屢次發(fā)生犬瘟熱疑似病例,所有患病動物都曾經(jīng)免疫過Onderstepoort株或CDV3 活苗,為了弄清免疫動物CDV山東株的基因特征,本試驗對13個來自免疫水貂、狐貍和犬的CDVH基因進行了測序和分析。
1材料和方法
1.1臨床病料
2012年4月至2014年6月,收集門診上來自于山東省不同地區(qū)有免疫接種史的自然發(fā)病死亡動物。據(jù)畜主口述,這些動物臨床癥狀包括發(fā)熱、化膿性的眼和鼻分泌物、足墊變厚。病理解剖發(fā)現(xiàn)疑似CDV感染的病變,如支氣管炎、扁桃體炎、胃腸炎等。隨機選取其中的13份病例,采集肺和腦組織樣品,勻漿之后凍融3次,4 ℃ 8 000 r/min 離心 10 min,取上清保存于-80 ℃ 冰箱備用。
1.2RNA提取
用TRIzol 試劑(Invitrogen,美國)按說明書操作程序提取組織上清中的總 RNA,提取到的 RNA 溶解在20 μL DEPC水中。CDV3 疫苗毒和健康水貂組織分別作為陽性對照和陰性對照。
1.3RT-PCR檢測CDV
取3 μL總RNA采用一步法 RT-PCR擴增265 bpN基因片段,用于CDV 的檢測。引物見表1。反應(yīng)條件如下:42 ℃ 60 min,95 ℃ 5 min;94 ℃ 30 s,50.8 ℃ 40 s,72 ℃ 1 min,30個循環(huán);最后72 ℃延伸10 min。用引物CDV-H-F/CDV-H-R擴增H全基因,用反轉(zhuǎn)錄酶XL(AMV,200 U/μL)(TaKaRa,大連)和9-Random Hexamers合成cDNA。PCR反應(yīng)條件為:95 ℃預(yù)變性5 min;94 ℃ 30 s,53 ℃ 1 min,72 ℃ 2 min,35 個循環(huán);72 ℃延伸10 min。

表1 本研究使用的引物
注:a.相對于Onderstepoort株(AF378705)基因組。
Note:a.Corresponding to positions of the Onderstepoort strain genome(AF378705).
1.4基因克隆與測序
PCR產(chǎn)物在1% 瓊脂糖(Biowest,西班牙)凝膠電泳后,目的條帶(1 921 bp)用EZ-10 Spin 柱式 DNA 膠回收試劑盒(上海生工)回收純化。純化的PCR 產(chǎn)物克隆至pMD19-T 載體(TaKaRa,Japan),挑取3個克隆進行Sanger測序,獲得的序列用DNASTAR SeqMan 軟件進行拼接。
1.5H基因進化樹和氨基酸分析
采用MEGA 6.06[14]軟件Clustal W方法進行序列比對和編輯。依據(jù)參考文獻[15]報道的方法,采用MEGA6.06軟件鄰接法對本試驗獲得的13個CDV山東株和35 個 CDV 參考株(13個基因型代表株)構(gòu)建系統(tǒng)發(fā)育進化樹,核苷酸水平差異小于5% 的CDV 毒株歸為同一基因型[16]。利用MEGA 6.06 軟件分析了13個CDV山東株氨基酸序列,關(guān)鍵點的氨基酸位點包括CDV受體-信號淋巴細胞激活分子結(jié)合位點[17]也進行了分析。 H蛋白上可能的N-糖基化位點和抗原性位點分別用NetNGlyc1.0 和DNASTAR Protean 軟件進行分析。
2結(jié)果與分析
2.1免疫動物CDV的檢測
所有的13個病例全部顯示CDV N PCR陽性,H基因擴增、克隆、測序、拼接后登陸GenBank獲得登錄號 KP127955~KP127967,13個臨床樣本信息見表2。
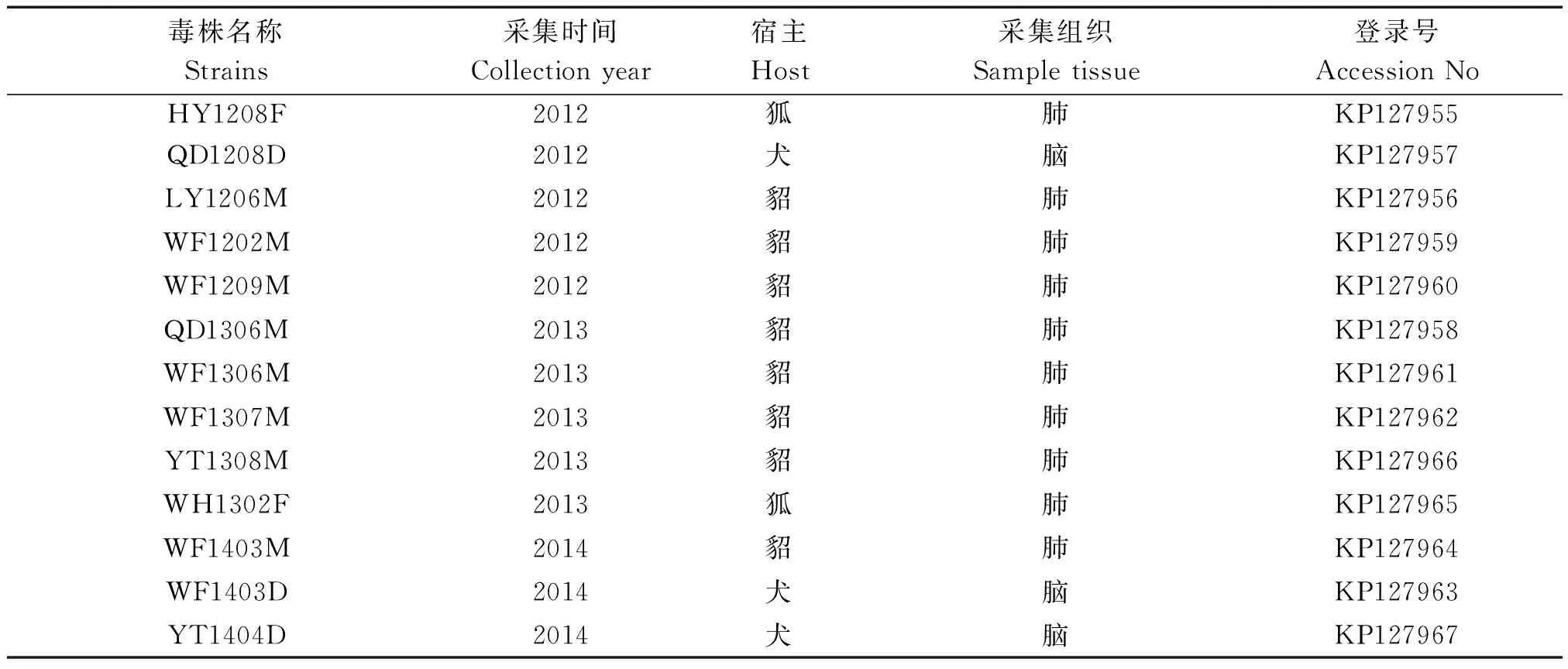
表2 臨床樣本信息
2.2免疫動物CDVH基因核苷酸序列分析
序列分析顯示,13個來自于免疫動物的CDV山東株之間核苷酸同源性較高,為99.1%~100.0%(表3);13個山東株與Asia-1型的095Cr(Accession No:AB286952)、HLJ(07)1(Accession No:EU325721)和HeB(09)3(Accession No:HM448832)同源性最高(96.5%~98.0%),與經(jīng)典CDV株(America-1型或疫苗型)CDV3(Accession No:DQ778941)、Onderstepoort(Accession No:EU143737)和 Lederle(Accession No:DQ903854)核苷酸同源性僅有90.1%~90.9%。CDVH基因編碼區(qū)(1 824 nt)進化樹分析顯示13個山東株均屬于Asia-1基因型(圖 1)。
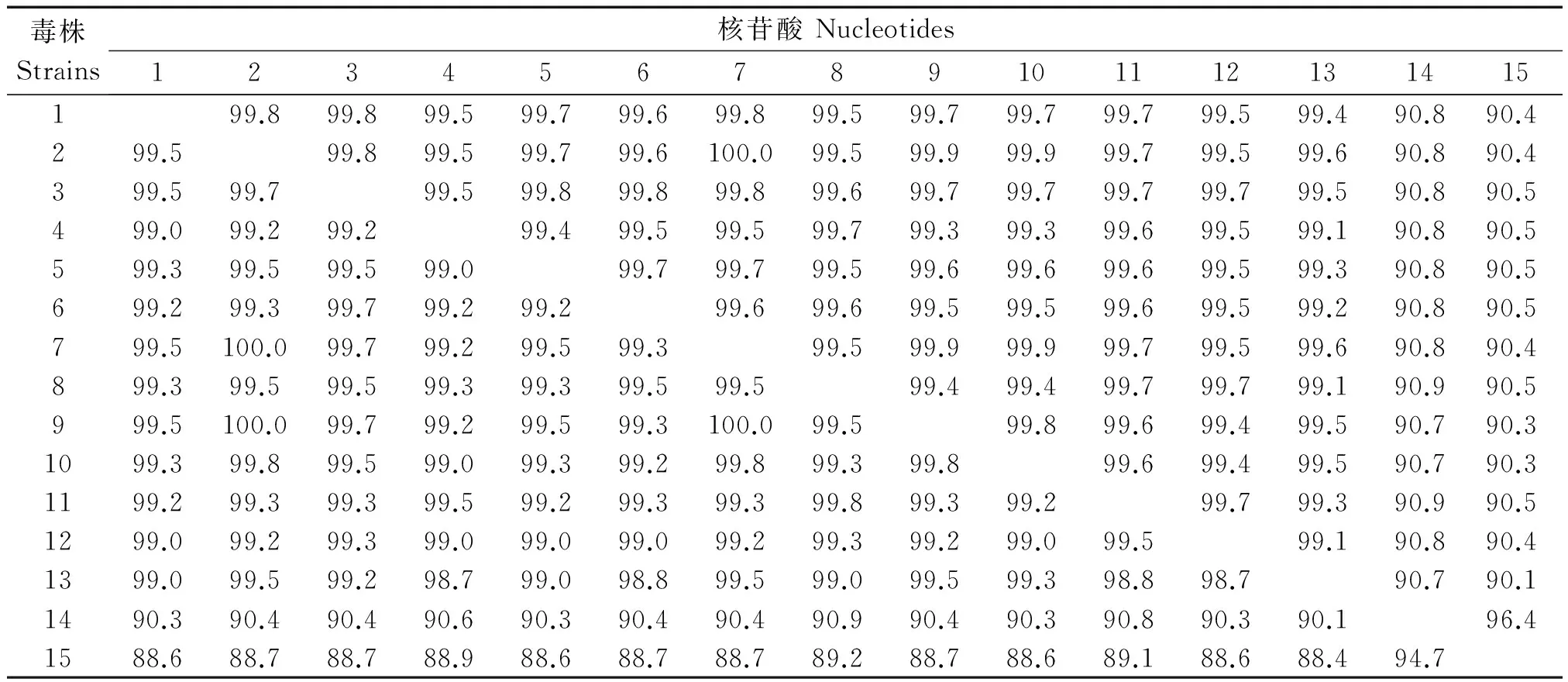
表3 CDV H基因核苷酸和氨基酸同源性比較
注:1.HY1208F;2.LY1206M;3.QD1208D;4.QD1306M;5.WF1202M;6.WF1209M;7.WF1306M;8.WF1307M;9.WF1403D;10WF1403M;11.WH1302F;12.YT1308M;13.YT1404D;14.CDV3;15.Onderstepoot。
2.3免疫動物CDVH基因推導(dǎo)氨基酸分析
氨基酸序列分析顯示,13個山東株之間氨基酸同源性為98.7%~100.0%,與Onderstepoort株和CDV3 氨基酸同源性分別為88.4%~89.2% 和 90.1%~90.9%(表 3)。與Onderstepoort 和 CDV3 相比,來源于免疫動物的13個山東株有91個氨基酸位點的變異。與GenBank登錄的11株2012年之前的非免疫動物來源的中國毒株相比,13個山東株H基因有4個氨基酸的一致性變化,即P200S、M263I、I542N 和Y549H(表4)。LY1206M,WF1306M,WF1403D,WF1403M和YT1404D株在306位是天冬酰胺酸,以前從未報道過。13個山東株H基因530,549位氨基酸殘基沒有區(qū)別。受體SLAM 結(jié)合位點(187-191,503,520,526,529,537,539,548,550)上,13個山東株與疫苗株相比發(fā)生R502K變異,HY1208F株在 537位由酪氨酸(Y)突變?yōu)榘腚装彼?C),其余位點均沒有變異。
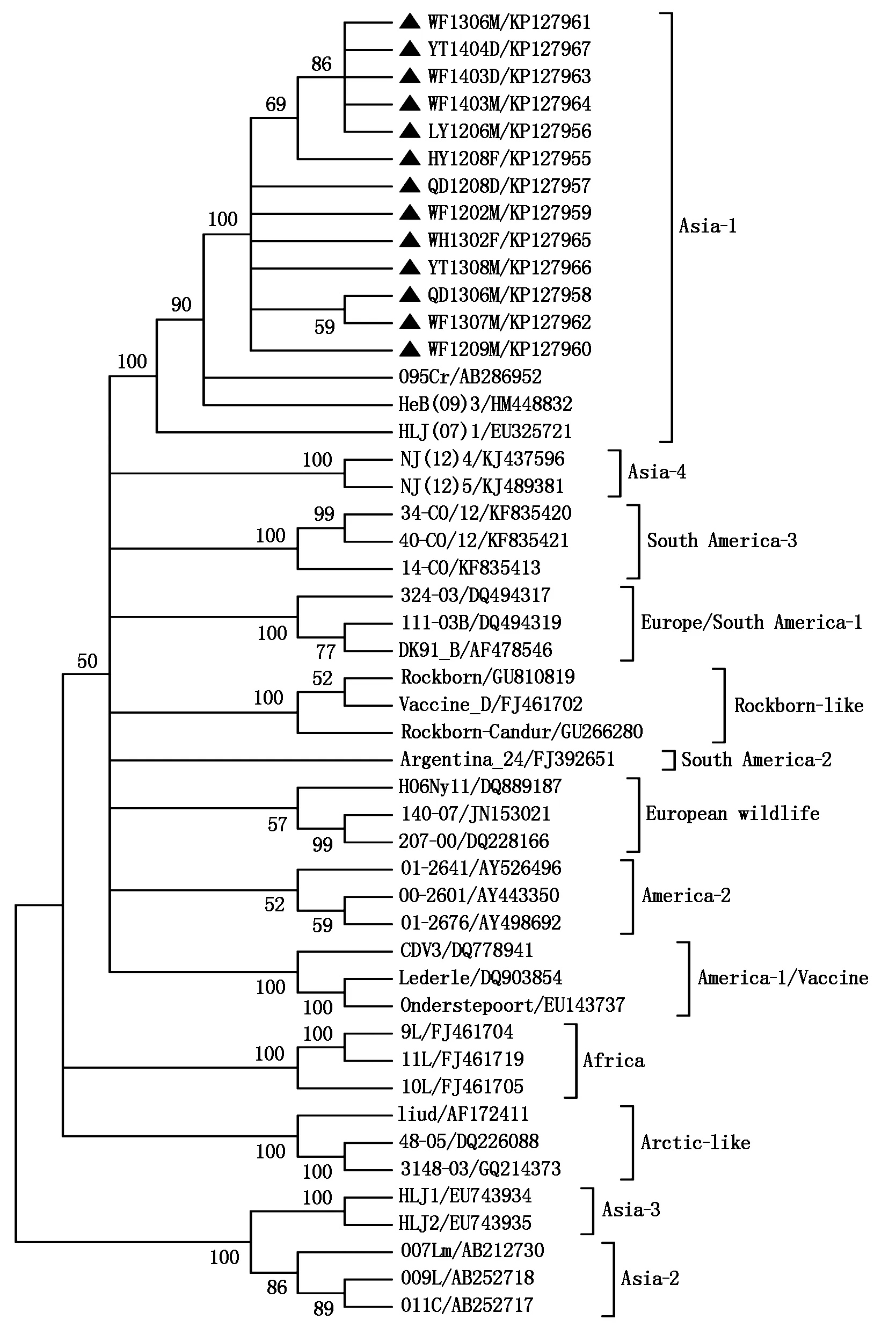
13個山東株和35個參考株采用鄰接法構(gòu)建進化樹,用Tamura 3-parameter計算進化距離,Bootstrap(1 000 重復(fù))
毒株Strains氨基酸殘基Residues187-19120026330636945250250352052652953053753954254854955014/GQ332530RCSGAPMHEIRDYDRGYYITYPCDV-JT1/HM749644..........K...........giantpanda/AF178038..........K...........HeB(07)3/EU325723..........K...........HeB(09)3/HM448832..........K...........HL/EF445052..........K....N......HLJ1/EU743934.......Y..K....R......JL(07)3/EU564812..........K...........
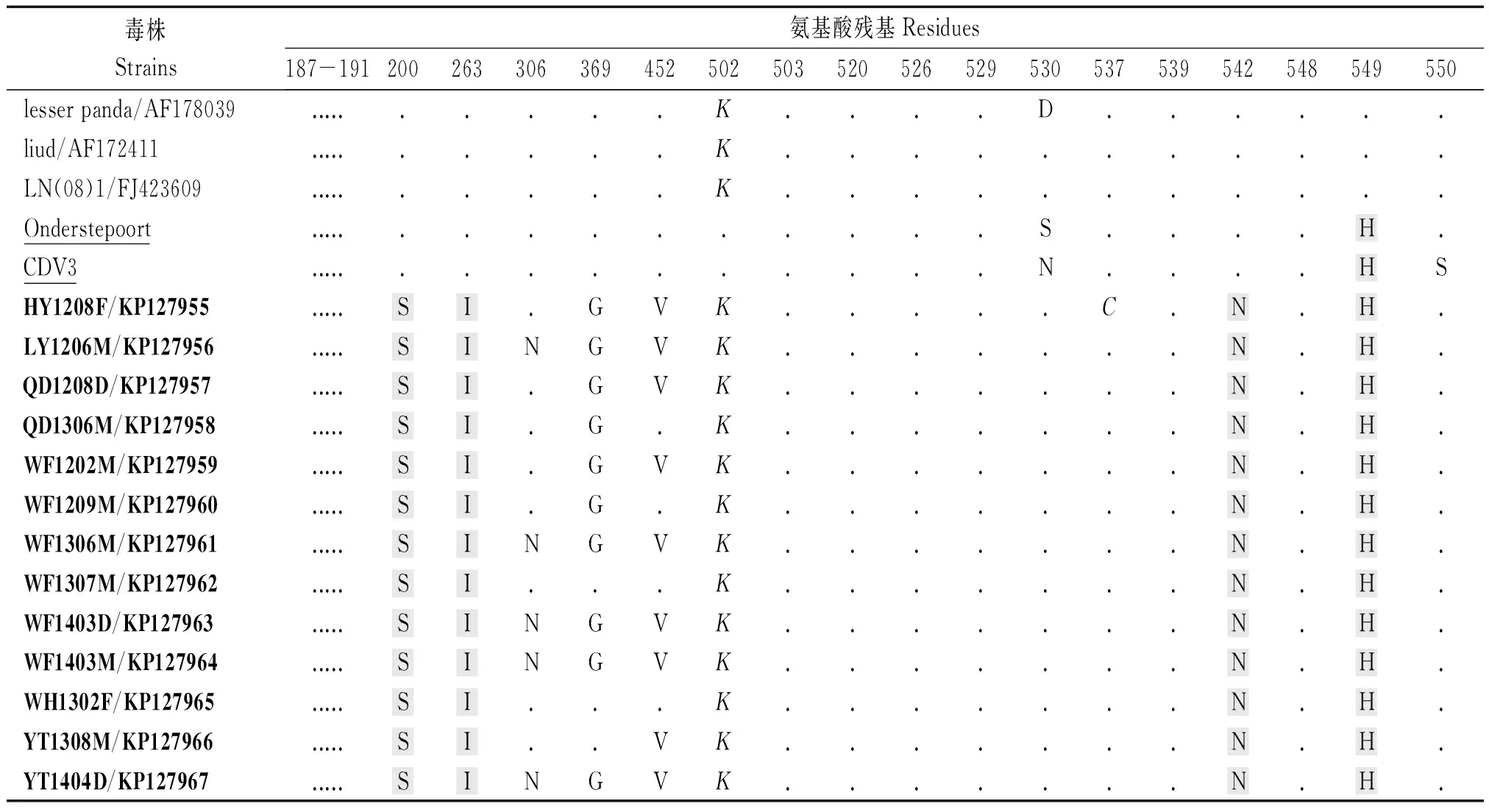
表4(續(xù))
注:上面11個CDV毒株為隨機選取的2012年以前報道的中國株;下面13株為本研究從免疫動物檢測的CDV(加粗);下劃線為疫苗株,以14/GQ332530株為基準;氨基酸殘基相同的以黑點(.)表示;SLAM受體結(jié)合區(qū)域用斜體表示;免疫動物來源的CDV與疫苗株及2012年以前的毒株相比有4個氨基酸殘基的不同(陰影)。
Note:Upper 11 CDV strains are randomly selected from published Chinese isolates before 2012;Lower 13 strains were isolated from vaccinated animals in this paper(bold);Vaccine strains were underlined;Residues identical to those of 14/GQ332530 are indicated with an dark spot(.);Residues of SLAM receptor binding site are italic;4 residues difference between vaccinated and nonvaccinated animals are shadowed.
2.4免疫動物CDV H蛋白抗原性分析
13個山東株潛在N-糖基化位點比Onderstepoort株多6個或7個,比CDV 3株多3個或4個。與Ondersteporrt和CDV3疫苗株相比,13個山東株在309位和542位、12個山東株在584位增加了潛在的N-糖基化位點(表5)。LY1206M、WF1403D、WF1403M和YT1404D株在306位有一潛在的 N-糖基化位點。
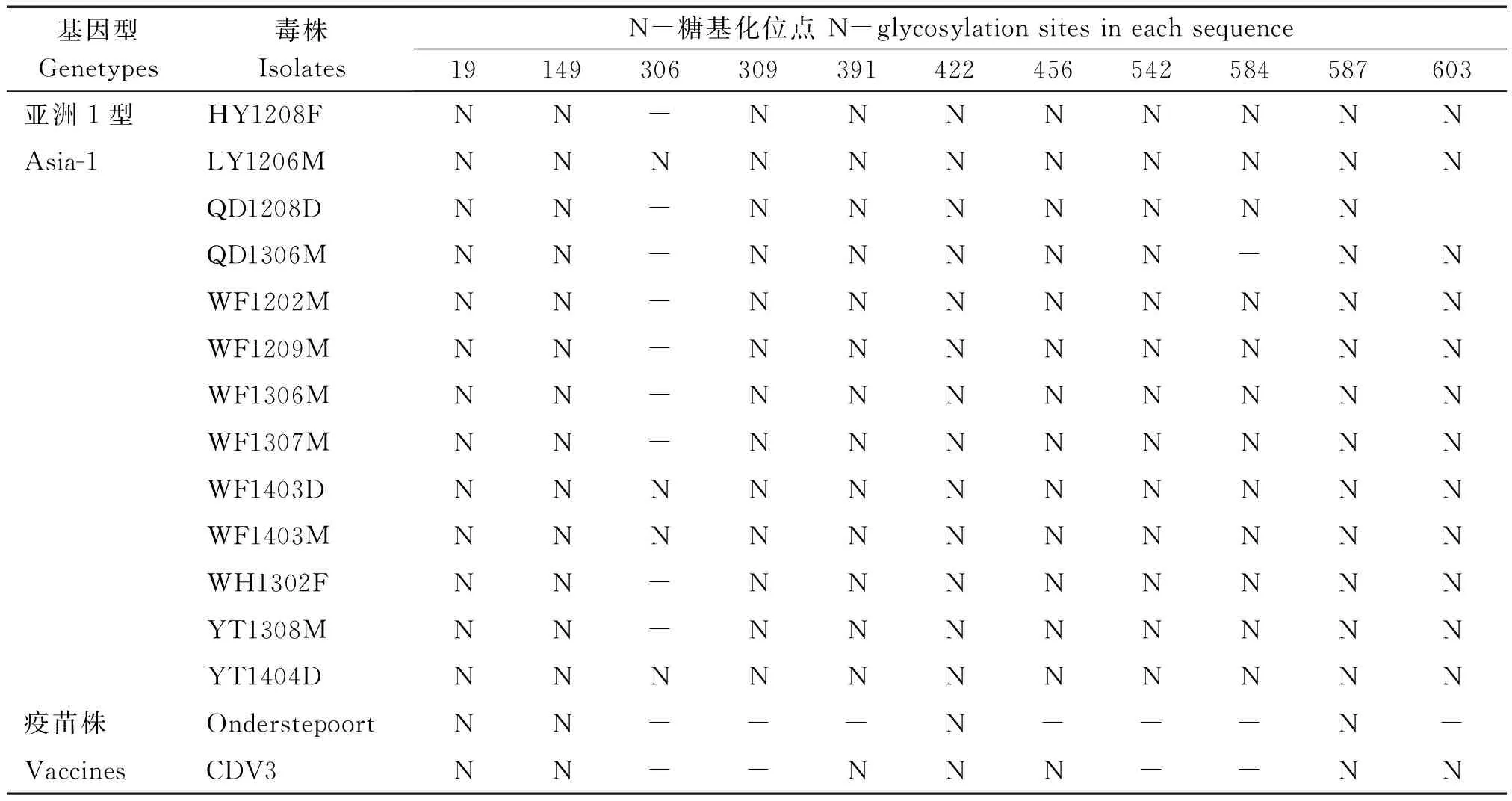
表5 13個山東株和疫苗株CDV H基因潛在的N-糖基化位點
注:N.潛在糖基化位點;-.陰性。
Note:N.Potential N-glycosylation sites;-.Negative.
抗原性分析表明,13個CDV山東株H蛋白推導(dǎo)的氨基酸疏水性和疫苗株不同(圖 2)。Jameson-Wolf 表位預(yù)測表明13個CDV山東株在540-549位比疫苗株Onderstepoort 和CDV3多了一個抗原性區(qū)域,氨基酸210-230 和327-347(Onderstepoort)位點抗原性也明顯不同(圖2)。
540-549位氨基酸增加的一個抗原區(qū)域用黑箭頭標示(↑);其他2個不同的抗原區(qū)域分別用倒三角(▼)和下劃線指示(-)。
3討論
我國曾報道過犬、貉、貂、狐、大熊貓、小熊貓和猴的 CDV 感染[5,10-13],以前的研究表明在我國大陸有6個基因型(Asia-1、Asia-3、Asia-4、Rockborn-like、Arctic-like、Recombinant)[5,10-13,18]。本研究中,所有的CDV陽性患病動物根據(jù)臨床記錄均有免疫接種史。有趣的是,來自于不同宿主(貂、狐和犬)、不同地區(qū)(濰坊、青島等)、不同時間(2012-2014年)的13個CDV毒株H基因序列高度同源(99.1%~100.0%),說明這些毒株由同一個病毒進化而來,在2012-2014年流行傳播引起免疫動物感染發(fā)病。進化樹分析表明這13個毒株均為Asia-1基因型,這是我國目前CDV流行的主要基因型,也是目前我國水貂和山東省報道的唯一基因型,說明水貂來源的CDV-H基因變異較少。因此,貂場CDV 控制策略應(yīng)該主要放在加強生物安全,建立潔凈農(nóng)場;免疫接種犬只,因為犬是已知的CDV儲存宿主[19]。
活疫苗Onderstepoort和CDV3(Onderstepoort克隆株)在我國被廣泛應(yīng)用于免疫接種。本研究中,來源于免疫動物的13個CDV山東株與疫苗株在進化樹上明顯處于不同分支,表明這些動物不可能是由于疫苗毒株返強導(dǎo)致發(fā)病。與2012年之前的中國毒株及疫苗株相比,13個山東株H基因有4個氨基酸的一致性變異,特別是542位異亮氨酸突變?yōu)樘於0匪?導(dǎo)致抗原區(qū)域和N-糖基化位點增加。蛋白上氨基酸的疏水性和N-糖基化位點會影響免疫原性[20],H蛋白上N-糖基化會掩蓋抗原表位,從而妨礙H蛋白與中和抗體的結(jié)合;N-糖基化位點的變異也會影響CDV毒力[21]。新毒株發(fā)生抗原性和毒力變異,能夠逃避疫苗免疫保護,可能是臨床上免疫失敗的原因。對日本東京2個動物醫(yī)院的調(diào)查結(jié)果表明,2/3 被診斷為犬瘟熱的犬有免疫接種史,大多數(shù)犬體內(nèi)有高水平的Onderstepoort株中和抗體,但針對野毒株的中和抗體水平很低,表明Onderstepoort株和野毒株膜蛋白抗原表位不同[22-23]。因此,選擇新分離的當(dāng)?shù)亓餍兄曜鳛镃DV疫苗毒株尤為重要。
CDV毒株致病力增強也可能是導(dǎo)致免疫失敗的原因。由于受體 SLAM是CDV入侵機體、宿主-病毒相互作用中的關(guān)鍵分子[17],也分析了CDV-H上相應(yīng)的 SLAM 結(jié)合位點(位點1:502,503;位點2:526,529,548,550;位點3:187-191;位點4:520,537,539,548)[17]。SLAM 結(jié)合位點 1,2,3,4 上的氨基酸殘基均沒有變異,502位氨基酸除外,13 個CDV 山東株是 K502,2個疫苗株是R502,但這種取代被認為不大可能影響病毒-受體結(jié)合[17]。CDV-H Y549被認為是CDV感染犬科動物的關(guān)鍵氨基酸[24],然而,13個來源于貂、狐和犬的 CDV山東株及 2 個疫苗株均表現(xiàn)為Y549H。因此,本試驗基因分析結(jié)果對了解CDV-宿主相互作用僅能提供有限信息,免疫失敗是否由CDV毒力變異引起尚需通過動物試驗進一步證實。
犬瘟熱病毒的流行和傳播不僅危害貂狐等經(jīng)濟動物的健康,還威脅東北虎、熊等野生動物的健康[3,19]。本研究報道了新的CDV野毒株,其抗原區(qū)域和N-糖基化位點都比疫苗株多,加強對臨床上犬瘟熱疫苗免疫失敗的原因探究,有助于保護毛皮動物、伴侶動物和野生動物的健康。
參考文獻:
[1]Appel M J,Yates R A,Foley G L,et al.Canine distemper epizootic in lions,tigers,and leopards in North America[J].Journal of Veterinary Diagnostic Investigation,1994,6(3):277-288.
[2]Sakai K,Yoshikawa T,Seki F,et al.Canine distemper virus associated with a lethal outbreak in monkeys can readily adapt to use human receptors[J].Journal of Virology,2013,87(12):7170-7175.
[3]Seimon T A,Miquelle D G,Chang T Y,et al.Canine distemper virus:an emerging disease in wild endangered Amur tigers(Pantheratigrisaltaica)[J].mBio,2013,4(4).pii:e00410-13.
[4]Si W,Zhou S,Wang Z,et al.A multiplex reverse transcription-nested polymerase chain reaction for detection and differentiation of wild-type and vaccine strains of canine distemper virus[J].Virology Journal,2010,7:86.
[5]Zhao J J,Zhang H L,Bai X E,et al.Emergence of canine distemper virus strains with two amino acid substitutions in the haemagglutinin protein,detected from vaccinated carnivores in North-Eastern China in 2012-2013[J].Veterinary Journal,2014,200(1):191-194.
[6]Rima B K.The proteins of morbilliviruses[J].Journal of General Virology,1983,64(6):1205-1219.
[7]Bolt G,Jensen T D,Gottschalck E,et al.Genetic diversity of the attachment(H)protein gene of current field isolates ofCaninedistempervirus[J].Journal of General Virology,1997,78(2):367-372.
[8]Martella V,Cirone F,Elia G,et al.Heterogeneity within the hemagglutinin genes ofCaninedistempervirus(CDV)strains detected in Italy[J].Veterinary Microbiology,2006,116(4):301-309.
[9]Espinal M A,Díaz F J,Ruiz-Saenz J.Phylogenetic evidence of a newCaninedistemperviruslineage among domestic dogs in Colombia,South America[J].Veterinary Microbiology,2014,172(1-2):168-176.
[10]Guo L,Yang S L,Wang C D,et al.Phylogenetic analysis of the haemagglutinin gene ofCaninedistempervirusstrains detected from giant panda and raccoon dogs in China[J].Virology Journal,2013,10:109.
[11]Bi Z W,Wang Y S,Wang X L,et al.Phylogenetic analysis ofCaninedistempervirusin domestic dogs in Nanjing,China[J].Archives of Virology,2015,160(2):523-527.
[12]Zhao J J,Yan X J,Chai X L,et al.Phylogenetic analysis of the haemagglutinin gene ofCaninedistempervirusstrains detected from breeding foxes,raccoon dogs and minks in China[J].Veterinary Microbiology,2010,140(1/2):34-42.
[13]Li W,Li T,Liu Y,et al.Genetic characterization of an isolate ofCaninedistempervirusfrom a Tibetan Mastiff in China[J].Virus Genes,2014.49(1):45-57.
[14]Tamura K,Peterson D,Peterson N,et al.MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Molecular Biology and Evolution,2011,28(10):2731-2739.
[15]Budaszewski R F,Pinto L D,Weber M N,et al.Genotyping ofCaninedistempervirusstrains circulating in Brazil from 2008 to 2012[J].Virus Research,2014,180:76-83.
[16]Mochizuki M,Hashimoto M,Hagiwara S,et al.Genotypes ofCaninedistempervirusdetermined by analysis of the hemagglutinin genes of recent isolates from dogs in Japan[J].Journal of Clinical Microbiology,1999,37(9):2936-2942.
[17]Ohishi K,Suzuki R,Maeda T,et al.Recent host range expansion ofCaninedistempervirusand variation in its receptor,the signaling lymphocyte activation molecule,in carnivores[J].Journal of Wildlife Diseases,2014,50(3):596-606.
[18]Han G Z,Liu X P,Li S S.Cross-species recombination in the haemagglutinin gene ofCaninedistempervirus[J].Virus Research,2008,136(1/2):198-201.
[19]Cottrell W O,Keel M K,Brooks J W,et al.First report of clinical disease associated withCaninedistempervirusinfection in a wild black bear(ursus americana)[J].Journal of Wildlife Diseases,2013,49(4):1024-1027.
[20]Jameson B A,Wolf H.The antigenic index:a novel algorithm for predicting antigenic determinants[J].Computer Applications in the Biosciences,1988,4(1):181-186.
[21]Sawatsky B,von Messling V.Canine distemper viruses expressing a hemagglutinin without N-glycans lose virulence but retain immunosuppression[J].Journal of Virology,2010,84(6):2753-2761.
[22]Ohashi K,Iwatsuki K,Nakamura K,et al.Molecular identification of a recent type of canine distempervirus in Japan by restriction fragment length polymorphism[J].Journal of Veterinary Medical Science,1998,60(11):1209-1212.
[23]Iwatsuki K,Tokiyoshi S,Hirayama N,et al.Antigenic differences in the H proteins ofCaninedistemperviruses[J].Veterinary Microbiology,2000,71(3/4):281-286.
[24]Nikolin V M,Wibbelt G,Michler F,et al.Susceptibility of carnivore hosts to strains ofCaninedistempervirusfrom distinct genetic lineages[J].Veterinary Microbiology,2012,156(1/2):45-53.
Sequence Analysis of the Hemagglutinin Gene ofCaninedistempervirusfrom Vaccinated Dogs,Minks and Foxes
HUANG Juan,SUI Xiaomei,WANG Yinan,SHAN Hu
(College of Animal Science and Veterinary Medicine,Qingdao Agricultural University,Key Laboratory of Preventive Veterinary Medicine in Universities of Shandong,Qingdao266109,China)
Abstract:In order to understand whether new genotype emerged,we detected Canine distemper virus(CDV)in immunized dogs,breeding foxes and minks in different regions by amplification of nucleocapsid(N)and hemagglutinin(H)gene.13 strains from CDV-positive cases(8 minks,2 foxes and 3 dogs)were sequenced.The results showed that the 13 Chinese strains shared 99.1%-100.0% nucleotide identity.Compared with those of other Chinese CDV strains isolated before 2012,four consistent amino acid changes,P200S,M263I,I542N and Y549H,were observed in the H coding sequence of CDV isolated from vaccinated animals,which contribute to an additional N-glycosylation site and antigenic epitope.All of the strains obtained from 13 samples were characterized as genotype Asia-1 according to the sequence analysis of the complete H gene,which was the sole genotype in minks in China.These findings provide a comprehensive understanding of current immune failure and the molecular epidemiology of CDV in China.
Key words:Canine distemper virus;Hemagglutinin gene(H);Vaccination failure
doi:10.7668/hbnxb.2016.02.037
中圖分類號:S85;S432.4+1
文獻標識碼:A
文章編號:1000-7091(2016)02-0231-08
通訊作者:單虎(1967-),男,山東威海人,教授,博士,主要從事動物傳染病防制與生物制品學(xué)研究。
作者簡介:黃娟(1977-),女,河南信陽人,副教授,博士,主要從事動物傳染病防制研究。
基金項目:國家自然科學(xué)基金資助項目(31472196)
收稿日期:2016-02-17
