
大刺鰍性腺發育的組織學研究
2016-06-08 16:47:32廖顯平林偉強許政生陳挺梁文浪蘇怡林
安徽農學通報 2016年10期
廖顯平 林偉強 許政生 陳挺 梁文浪 蘇怡林

摘 要:該文應用解剖觀測和組織學的方法研究了大刺鰍的性腺發育特征。結果表明:大刺鰍生殖細胞劃分為5個時相,卵巢和精巢發育劃分為Ⅰ~Ⅵ期。雌魚群體的GSI值變幅為1.05%~7.21%,屬于多次分批產卵類型,繁殖高峰期為5月;雄魚群體GSI值波動在0.09%~0.39%,與雌魚發育同步。產卵和排精后,卵巢和精巢都恢復到Ⅱ期進行越冬,次年3月份重新發育至性成熟。
關鍵詞:大刺鰍;精巢;卵巢;組織學
中圖分類號 S917.4 文獻標識碼 A 文章編號 1007-7731(2016)10-0114-03
大刺鰍(Mastacembelue armatus)在分類上隸屬于鱸形目(Perciformes)、刺鰍科(Mastacembelidae)刺鰍屬(Mastaceblus Scopoli),俗稱辣錐、豬母鋸、石錐等,主要分布在我國的長江、珠江等水系,以廣東、廣西、福建等地江河中的野生資源較為豐富[1]。其肉質細嫩、味道鮮美、蛋白質含量高,深受消費者的喜愛。近年來,學者們已經對大刺鰍的生物學特性、消化系統、馴養生殖、肌肉分析等都進行了研究[2-9],但對大刺鰍性腺發育的研究尚無全面深入的報道。為此,筆者采用組織學方法研究了大刺鰍卵巢和精巢發育的特點,旨在為大刺鰍的資源保護、增殖、人工繁殖技術等提供科學依據。
1 材料與方法
1.1 實驗材料 本研究于2012年1~12月在肇慶市水產科學研究所進行。大刺鰍為經馴養的野生個體,每批隨機抽取大刺鰍5~10尾。共組織切片40批,取大刺鰍223尾,其中雌性個體128尾,雄性個體95尾,平均體重47.6g/尾。
1.2 研究方法 首先解剖觀測大刺鰍性腺發育情況,測量性腺重量和凈體重;并對第Ⅳ~Ⅴ期雌魚統計絕對懷卵量。測量后迅速取性腺前、中、后段放入Bouin氏液中固定,經系列酒精脫水后進行石蠟包埋、切片、H.E染色、固封,最后在光學顯微鏡下觀察和對各時相細胞拍照。性腺和生殖細胞的發育分析參考樓允東[10]的分期標準,將大刺鰍性腺發育分為Ⅰ~Ⅵ期。相關計算公式如下:
絕對懷卵量=取樣中的卵粒數/取樣重量(g)×卵巢重(g);
性腺成熟系數(GSI)=性腺重(g)/魚體重(g)×100。
2 結果與分析
2.1 卵巢發育 雌性大刺鰍的卵巢是一對延長的囊狀結構,前端分離,位于鰾腹壁兩側、肝胰臟背方,左右對稱,后段末端匯合成一短的呈“Y”字型輸卵管通泄殖孔,外有極薄的腹膜包裹,它們通過卵巢系膜與體腔壁層相連。成熟時的絕對懷卵量在1 386~4 175粒。
2.1.1 第Ⅰ期卵巢 出現在當年生幼體,卵巢呈透明細絲狀,淺灰色,外觀上難以分辨雌雄。切片觀察卵巢,生殖上皮具增殖分生而成的密集的卵原細胞,大小不一,形態不規則,有梨形、橢圓形等,卵徑為32.5~45.0μm,部分卵原細胞成束排列,胞質透亮著色淺,細胞核較大,核徑為19.0~20.5μm,其中有染色深的核仁1~2個。
2.1.2 第Ⅱ期卵巢 卵巢呈棒狀,半透明,淺黃色。肉眼尚不能看清卵粒,生殖上皮及結締組織突入卵巢腔形成產卵板,以第Ⅱ時相卵母細胞為主,也有少量卵原細胞。卵母細胞排列緊密,形態不一,卵徑為48.0~317.5μm,核徑為19.5~95.0μm。有核仁22~36個,靠近核膜內側分布。在細胞中出現卵黃核,細胞質呈強嗜堿性,被蘇木精深染成紫藍色,胞質外周出現一層濾泡細胞。胞核與胞質間形成一透明層,透明層內物質密度稀少,顯得透亮,厚度為9.0~13.0μm,隨著細胞繼續發育,透明層變薄漸消失。
2.1.3 第Ⅲ期卵巢 卵巢呈黃色,卵巢外膜上的血管明顯且呈分支狀,透過卵巢膜可見到卵粒,但不易分離脫落。卵巢組織中以第Ⅲ時相卵母細胞為主,但第Ⅱ時相的卵母細胞也較多。Ⅲ時相卵母細胞大多近圓形或橢圓形,卵徑為310.0~577.0μm,核近球形,核膜凹凸不平,核徑為96.0~120.0μm,核仁11~19個。細胞中,靠近質膜的胞質外周出現2~6層直徑為10.0~19.5μm小液泡。后期胞質的皮質部分出現一些被H.E染成紫紅色的卵黃顆粒。
2.1.4 第Ⅳ期卵巢 卵巢呈亮黃色半透明,體積明顯增大,卵巢外膜的血管發達,粗大且分支,呈深紅色。卵巢壁薄,卵粒飽滿,易分離脫落,此時用力擠壓魚腹部可擠出未成熟卵母細胞。以第Ⅳ時相卵母細胞為主,亦有較多的第Ⅱ、第Ⅲ時相卵母細胞。Ⅳ時相卵母細胞大多近圓形,卵徑達862.5~1 350.0μm,核徑為114.0~150.0μm。卵母細胞進入卵黃積累的大生長期,卵黃顆粒充滿細胞質,體積顯著增大,直徑為12.5~25.0μm。核極化,出現放射帶,并增厚達42.0~45.0μm,條紋清晰。
2.1.5 第Ⅴ期卵巢 卵巢呈黃亮色,發育到最大,占據腹部大部分體積。卵巢松軟,富有彈性,卵粒分離,處于流動狀態,擠壓腹部有透明成熟卵子從泄殖孔排出。卵巢以第Ⅴ時相和第Ⅳ時相末期卵母細胞為主,同時也有第Ⅲ時相和第Ⅳ時相早期的卵母細胞。卵子透明,卵徑為1 462.5~1 880.0μm,細胞核已移到卵細胞一側即動物極附近,核膜消失,核質與胞質融合,卵母細胞內充滿松散排列的卵黃顆粒。
2.1.6 第Ⅵ期卵巢 卵巢松軟如空袋狀,表面皺縮,卵巢充血呈肉紅色,不透明,卵細胞質和卵黃膠液化并形成許多空泡和一些吸收的卵子。卵巢產后不久重新回復到第Ⅱ期進行越冬。
2.2 精巢發育 大刺鰍雄性個體精巢是雙葉型結構,前段分離,位于鰾腹壁兩側,左右對稱,后段末端匯合成一短的“Y”字型輸精管通泄殖孔,精巢外面由腹膜包裹,并通過腹膜與腹腔側壁相連。精巢屬于小葉型,有結締組織將其分隔形成許多精小囊所構成,生精細胞在精小囊中發育。
2.2.1 第Ⅰ期精巢 精原細胞增殖期,出現在當年生幼魚中,精巢呈透明細絲線狀,淺灰色,緊貼腹膜,外表無法分辨雌雄。切片觀察到精巢中精原細胞開始增殖使其數量增多,且不定向地分散于間質細胞之間,此時小葉腔未出現。細胞較大,胞徑為9.25~10.75μm,胞質染色淺,核位于中央,核徑較小,為3.75~4.50μm,為弱嗜堿性。
2.2.2 第Ⅱ期精巢 精母細胞生長期,精巢為細帶狀,灰白色,體積略有增大。精葉中可見精原細胞和初級精母細胞,精原細胞數目顯著增多,2~6個細胞成束排列,細胞仍然為圓形或橢圓形,胞徑4.75~7.25μm,核徑2.00~2.55μm。精小葉中出現腔隙較小的小葉腔。精小囊內初級精細胞同步發育,細胞體積比精原細胞要小,核為強嗜堿性。
2.2.3 第Ⅲ期精巢 精母細胞成熟期,精巢為扁帶狀,呈淺白色。初級精母細胞沿小葉邊緣作單層或多層排列,小葉腔中出現數量較少的次級精母細胞,排列無規律,成堆排列。次級精母細胞為完成了第一次減數分裂的細胞,胞徑2.75~3.25μm,細胞核比初級精母細胞小,核徑1.70~1.95μm,染色較深。
2.2.4 第Ⅳ期精巢 精子細胞變態期,精巢為條塊狀,呈灰白色,表面有明顯的血管分布。精巢體積繼續增大,精小葉中存在有精原細胞、初級精母細胞、次級精母細胞和精子細胞,同時有少部分已完成變態的精子,呈不同步發育,主要以初級精母細胞、次級精母細胞和精子細胞為主。精子細胞排列緊密,橢圓形或圓,無明顯的細胞質,只含有強嗜堿性的細胞核,直徑1.75~2.50μm。
2.2.5 第Ⅴ期精巢 精子成熟期,精巢乳白色,體積肥厚,表面的血管更加明顯,呈鮮紅色。主要由精子細胞和成熟的精子組成,輕壓腹部有大量乳白色精液流出。小囊壁破裂,精子溢出至小葉腔,大量的成熟精子在小葉腔中呈渦流狀,精子頭部圓形,直徑1.05~1.75μm,為嗜堿性強,HE染色為深藍色,尾部細長。
2.2.6 第Ⅵ期精巢 精子退化吸收期,產后精巢萎縮,白色微紅至肉紅色,充血明顯,精小葉僅剩由精小囊圍成的一個空腔。大部分精子已經排出,精小葉中僅殘留少量的精子,小葉壁有少量精原細胞和精母細胞,經過短時期回復到第Ⅱ期越冬。
2.3 性腺發育周年變化
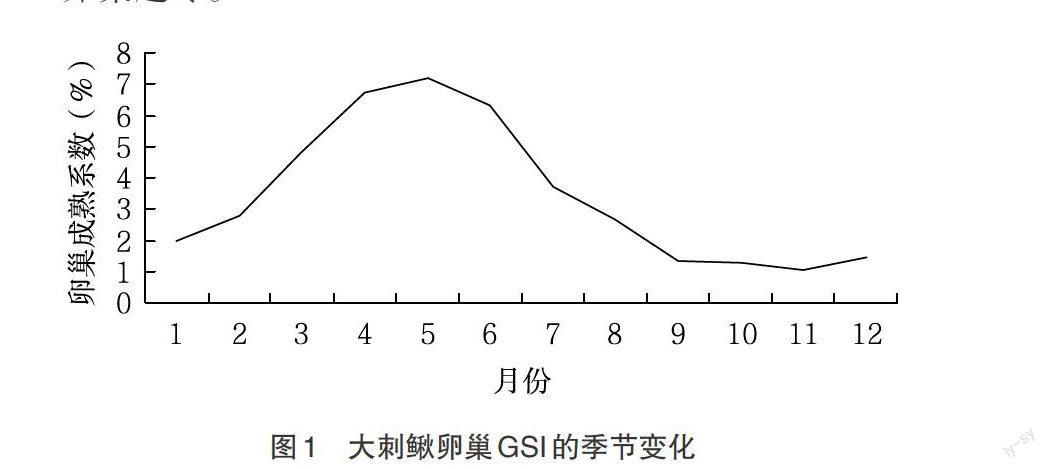
2.3.1 大刺鰍雌魚GSI周年變化 對大刺鰍卵巢周年的組織學觀察結果表明,該群體月平均GSI值變幅為1.05%~7.21%(如圖1),個體GSI值變幅為0.19%~20.38%。個體間GSI值差異較大,解剖中發現有些個體處在幼齡期,性腺尚未分化,屬于第Ⅰ期性腺。從GSI值的大小及時間分布可知,大刺鰍的繁殖期主要在4~7月,屬春夏季產卵類型魚類。而從8月份起到翌年2月份成熟系數變幅較小,證明大刺鰍是以Ⅱ期卵巢越冬。
2.3.2 大刺鰍雄魚GSI周年變化 據對95尾雄魚解剖及性腺切片的觀察,在整個生殖周期內,大刺鰍群體精巢的月平均GSI值波動在0.09%~0.39%(如圖2),個體GSI值為0.02%~0.94%。從圖2可知,大刺鰍的精巢3月份開始進一步發育,成熟在4~7月,從8月份起GSI值明顯下降,且成熟系數變幅較小,組織切片觀測證明大刺鰍是以Ⅱ期精巢越冬。
3 討論
3.1 大刺鰍雌魚繁殖特性 馴養的大刺鰍繁殖時間為4~7月,屬春夏季繁殖類型,5月為高峰期。而劉霆等[4]調查表明,都柳江三都河段的大刺鰍自然繁殖的時間為5~7月,盛產期在6月份;黃永春[2]研究表明,汀江大刺鰍的產卵期為4月底至11月初,產卵高峰期在7月和9月;李曉雙[6]調查表明,大刺鰍人工繁殖的季節為每年4月~7月。大刺鰍繁殖時間存在一定的差異。大刺鰍卵巢中各時相卵母細胞發育的非同步性在年周期中是十分明顯的。在以某一時相卵細胞為主的卵巢中都有該時相以下各期卵細胞的存在。顯微鏡下觀察發現,這些卵母細胞發育正常,可繼續發育成熟。因此,大刺鰍屬于多次產卵類型,結果與李曉雙[6]研究為一次產卵類型不符,與劉霆[4]、黃永春[2]等研究結果相一致。
3.2 大刺鰍雄魚性腺發育特征 實驗解剖和測量結果得知,大刺鰍雄魚的體重比同期雌魚的大但性腺重相比卻很小,這可能與其本身生物學特性有關。從圖1、圖2可以看出,大刺鰍雌雄個體的性腺發育時間是一致,都在5月份達到高峰,這與大刺鰍繁殖特性有關,最大程度確保受精同時進行,有利于種群的延續。這與曹運長等[11]對藍太陽魚、曹伏君等[12]對大彈涂魚的性腺發育研究時得出結論一致。與刀鱭[13]、長薄鰍[14]等大多硬骨魚類一樣,大刺鰍雄魚排精后恢復到Ⅱ期精巢越冬,也與有些種類不同:銀鯧[15]是以Ⅲ期越冬;瓦氏黃顙魚[16]是以Ⅳ期越冬,但各期的生精細胞特點是相一致的。
參考文獻
[1]潘炯華.廣東淡水魚類志[M].廣州:廣東科學技術出版社,1991:518-520.
[2]黃永春.汀江大刺鰍食性和繁殖生物學[J].水產學報,1999(S1):1-6.
[3]溫曉紅.汀江大刺鰍資源的保護及利用[J].中國水產,2005(10):72.
[4]劉霆,李建光,賀兵,等.大刺鰍的性腺調查和懷卵量比較分析[J].淡水漁業,2005,35(5):28-30.
[5]陳忠.大刺鰍的生物學及其養殖技術概要[J].中國水產,2005(10):22-23.
[6]李曉雙.大刺鰍人工繁殖研究初報[J].中國水產,2007(5):45.
[7]初慶柱,陳剛,張健東,等.大刺鰍消化系統的組織學研究[J].淡水漁業,2009,39(2):14-18.
[8]Watanabe T,Arakwa T,Kitajima C,et al.Effect of nutritional qulity of broodstock diets on reproduction of red sea bream[J].Nippon Saisan Gakkaishi,1984,50:495-501.
[9]朱定貴,陳濤.大刺鰍生殖季節雌魚的脂肪酸組成研究[J].廣東農業科學,2011,16:108-110.
[10]樓允東.組織胚胎學[M].2版.北京:中國農業出版社,1996:152-157.
[11]曹運長,李文笙,葉衛,等.藍太陽魚第一次性周期性發育的組織學[J].動物學雜志,2008,43(1):88-95.
[12]曹伏君,羅杰,劉楚吾.大彈涂魚性腺發育的組織學觀察[J].水生生物學報,2010,34(2):418-425.
[13]徐鋼春,聶志娟,張呈祥,等.刀鱭精巢發育的組織學研究[J].華中農業大學學報,2012,32(2):247-252.
[14]王志堅,殷江霞,張耀光.長薄鰍的精巢發育和精子發生[J].淡水漁業,2009,39(1):3-9.
[15]李云航,孫鵬,施兆鴻,等.養殖與野生銀鯧精巢發育形態學和組織學的初步比較[J].海洋漁業,2012,34(3):256-262.
[16]楊家云.瓦氏黃顙魚精巢發育及精子生物學研究[J].西南師范大學學報,2005,30(4):719-724.
(責編:張宏民)
