油茶籽殼總黃酮對黃曲霉毒素B1致肝細胞氧化應激的保護作用及機制
2016-06-14 16:43:05汪秀李菲黃利輝
江蘇農業科學 2016年4期
關鍵詞:氧化應激
汪秀+李菲+黃利輝

摘要:研究油茶籽殼總黃酮(CSF)對黃曲霉毒素B1(AFB1)誘導的人肝細胞(HL-7702)氧化應激的保護作用及其機制。以20 μg/mL AFB1處理2 d建立HL-7702細胞AFB1損傷模型;與AFB1損傷組比較,25、50、100 μg/mL CSF預處理HL-7702細胞1 d,細胞存活率顯著上升(P<0.05),細胞內活性氧(ROS)和脂質過氧化物丙二醛(MDA)水平顯著下降(P<0.05),超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、谷胱甘肽過氧化物酶(GSH-Px)活性顯著升高(P<005),核因子E2相關因子2(Nrf2)及其下游谷胱甘肽S轉移酶A2(GSTA2)的mRNA和蛋白表達水平均顯著升高(P<0.05),核轉錄因子κB(NF-κB)及下游炎癥因子(TNF-α、IL-6、IL-1β)的mRNA表達顯著下降(P<0.05)。因此,油茶籽殼總黃酮通過上調Nrf2,提高Nfr2下游抗氧化酶系的活性,阻斷了由ROS引起的脂質過氧化鏈式反應,同時下調NF-κB,減少炎癥因子相關基因的表達,從而拮抗AFB1對肝細胞的毒性損傷,提高了肝細胞存活率。
關鍵詞:油茶籽殼;總黃酮;黃曲霉毒素B1;氧化應激;細胞保護
中圖分類號: R284.1
文獻標志碼: A
文章編號:1002-1302(2016)04-0308-04
由黃曲霉、寄生曲霉產生的黃曲霉毒素B1(AFB1)經常被發現于發霉變質的飼料、食物中,被認為是中毒性最強、危害最大的一種霉菌毒素[1]。生物體攝入AFB1后,在不同器官誘發產生細胞內活性氧(ROS),破壞機體的氧化-抗氧化平衡,從而使脂質過氧化物丙二醛(MDA)積累,對脂類、蛋白質、DNA等生物大分子造成損傷[2],由此形成的氧化應激對肝臟的損傷是最大的[3]。研究表明,植物黃酮類物質能吸收包括AFB1在內的多種霉菌毒素,減輕其對動物體的損傷,保護肝臟,從而改善牲畜健康,提高畜產品產量。人參皂苷[4]、槲皮苷[5]、柚皮苷[6]、藤黃雙黃酮[7]等都被發現具有黃曲霉毒素細胞保護作用。然而,至今仍然沒有一種商業化的AFB1生物保護劑,主要是由于上述具有細胞保護作用的化合物來源十分有限,價格昂貴。因此,尋找易獲得、廉價的此類化合物來源(如農副產品)十分必要。我國是世界上油茶籽產量最大的國家,油茶籽殼是油茶籽榨油前除去的種皮,大量的廢棄茶籽殼一般被丟棄或用作燃料,既污染環境又浪費資源。研究表明,油茶籽殼中含有豐富的黃酮類物質[8],關于這類黃酮類物質提取工藝的研究已經很多,但對其功能研究目前僅局限在證明其具有較好的體外抗氧化性能[9-10]。因此,筆者推測油茶籽殼中的黃酮類物質所具有的抗氧化能力可以減輕AFB1對細胞的毒性作用。本研究采用AFB1最易損傷的正常肝細胞HL-7702細胞為材料,建立AFB1肝細胞損傷模型,探討油茶籽殼中黃酮類物質對AFB1肝細胞損傷的保護作用及其可能機制,旨在為油茶籽殼的綜合高效利用和天然AFB1生物保護劑開發提供理論依據。
1 材料與方法
1.1 油茶籽殼總黃酮(CSF)樣品制備
油茶籽購自浙江省嘉興市農貿市場,剝殼后自然曬干即得油茶籽殼,粉碎后備用。取5 g油茶籽殼粉碎樣品,依照姜天甲等[9]和高揚等[11]的方法,按1 g ∶30 mL比例加入60%乙醇溶液,浸泡30 min,40 ℃超聲波提取45 min,濾液減壓濃縮后加蒸餾水定容至50 mL,加等體積石油醚充分振蕩,靜置分層后分離收集,經40 ℃真空減壓旋轉蒸發后,制備得油茶籽殼總黃酮并用DMSO配成20 mg/mL儲備液,-20 ℃保存。
1.2 材料
人正常肝細胞系HL-7702購于中國科學院上海細胞庫;AFB1、DMSO、MTT均為Sigma公司產品,AFB1溶解于DMSO 中,配成1 mg/mL母液;RPMI-1640培養基、胰酶、青鏈霉素均為HyClone公司產品;胎牛血清購于四季青公司;MDA試劑盒,SOD、CAT、GSH-Px測定試劑盒購于南京建成工程研究所;ROS檢測試劑盒、BCA蛋白濃度測定試劑盒、PCR試劑盒、TRIzol、脫脂牛奶(BSA)均為碧云天公司產品;PrimeScript RT反轉錄試劑盒購于TaKaRa公司;小鼠抗人Nrf2、GSTA2、β-actin單克隆抗體均為Santa Cruz公司產品;HRP標記的山羊抗小鼠IgG購于武漢博士德生物公司;PCR引物均合成自上海博尚生物公司。
1.3 方法
1.3.1 細胞培養 人HL-7702正常肝細胞以RPMI-1640培養基(含10%胎牛血清、1%雙抗)于37 ℃、5%CO2培養。
1.3.2 AFB1致HL-7702細胞損傷模型建立 采用MTT法檢測HL-7702細胞增殖抑制率,試驗分為對照組和AFB1組。HL-7702細胞按5 000個細胞/孔接入96孔板中,每組設5個復孔,過夜培養待細胞貼壁后加化合物。對照組每孔加入含1%DMSO的完全培養基100 μL,AFB1組每孔加入含5、10、15、20、25、30、35、40 μg/mL AFB1的完全培養基100 μL。各組分別培養0.5、1、2 d后棄去孔內培養基,并加入100 μL含 MTT溶液(終濃度0.5 mg/mL)的無血清培養基,避光37 ℃孵育4 h,棄上清液,每孔加100 μL DMSO,振蕩使結晶物充分溶解,570 nm下測定各孔吸光度,細胞抑制率=(D對照-D試驗)/ D對照×100%。根據MTT結果,選擇AFB1的最佳濃度進行后續試驗。
1.3.3 油茶籽殼總黃酮對AFB1致細胞損傷的預保護試驗 細胞接板和MMT檢測方法同“1.3.2”節,試驗分組如下:設正常對照組、AFB1模型組和油茶籽殼總黃酮低劑量、中劑量、高劑量保護組。正常對照組:細胞先用含1%DMSO(CSF的溶劑)的完全培養基培養1 d,換液后再加入含1%DMSO(AFB1的溶劑)的完全培養基繼續培養2 d;AFB1模型組:細胞先用含1%DMSO(CSF的溶劑)的完全培養基培養1 d,換液后加入含有最佳損傷濃度AFB1(20 μg/mL)的完全培養基繼續培養2 d;CSF保護組:細胞先分別與含25、50、100 μg/mL CSF的完全培養基預孵1 d,換液后加入含有最佳損傷濃度AFB1(20 g/mL)的完全培養基繼續培養2 d。3 d培養結束后檢測細胞存活率,細胞存活率計算公式如下:
細胞存活率=D試驗/ D對照×100%。(1)
1.3.4 HL-7702細胞內MDA生成量的檢測 采用 “1.3.3”節所述方法處理HL-7702細胞,用硫代巴比妥酸(TBA)法測定MDA含量[12]。將各組細胞裂解后按照試劑盒說明書操作步驟進行操作,在532 nm處測定各孔吸光度,采用BCA試劑盒說明書規定的方法測定總蛋白濃度。
1.3.5 HL-7702細胞內ROS水平的檢測 采用 “1.3.3”節所述方法處理HL-7702細胞,采用二氯熒光素雙乙酸(DCFH-DA)法測定細胞內ROS水平。將各組細胞裂解后按照試劑盒說明書操作步驟進行,細胞裂解液與10 nmol/L DCFH-DA 37 ℃孵育30 min后,PBS洗細胞2次。熒光強度用熒光酶標儀以激發波長485 nm 和發射波長535 nm檢測。ROS相對含量計算公式如下:
ROS相對含量=處理組熒光強度/對照組熒光強度×100%。(2)
1.3.6 HL-7702細胞內抗氧化酶系含量測定 采用 “1.3.3”節所述方法處理HL-7702細胞,收集各組細胞,用冰PBS洗,然后在冰上用超聲破碎機200 V、30 s,間隔5 s,3次破碎細胞,取100 L細胞裂解液,按照試劑盒說明書操作步驟測定SOD、CAT、GSH-Px酶活性。酶活性以酶比活力單位(U/mg)表示,并用細胞總蛋白質含量作校正。
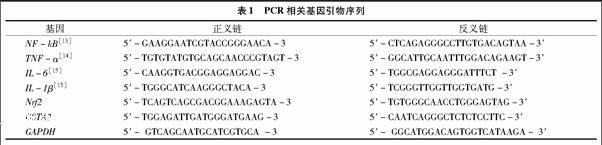
1.3.7 RT-PCR檢測相關基因的改變 采用 “1.3.3”節所述方法處理HL-7702細胞,收集各組細胞,提取細胞總RNA,采用PrimeScript RT反轉錄試劑盒并進行逆轉錄,得到各組細胞cDNA。PCR擴增條件均為:94 ℃ 5 min;94 ℃ 30 s,56 ℃ 30 s,72 ℃ 30 s,72 ℃ 8 min;循環數30。最后PCR產物在1%瓊脂糖凝膠中電泳分離,檢測基因引物見表1,GAPDH為內參。
1.3.8 Western blot檢測蛋白表達的改變 采用 “1.3.3”節所述方法處理HL-7702細胞,收集細胞,用PBS洗滌3次,加入裂解液,冰上充分裂解1 h,總蛋白樣品進行SDS-PAGE分離,轉移到硝酸纖維素膜上,用5%脫脂奶粉室溫封閉1 h,分別用稀釋的Nrf2、GSTA2和標準內參β-actin一抗4 ℃孵育過夜,洗膜后加稀釋的二抗室溫孵育2 h,加ECL發光試劑,顯影、定影。
1.3.9 統計學分析 所有試驗均重復3次,數據采用x±s表示,采用SPSS 18.0軟件統計分析,采用Duncans檢驗多重比較。
2 結果與分析
2.1 HL-7702細胞AFB1受損模型的建立
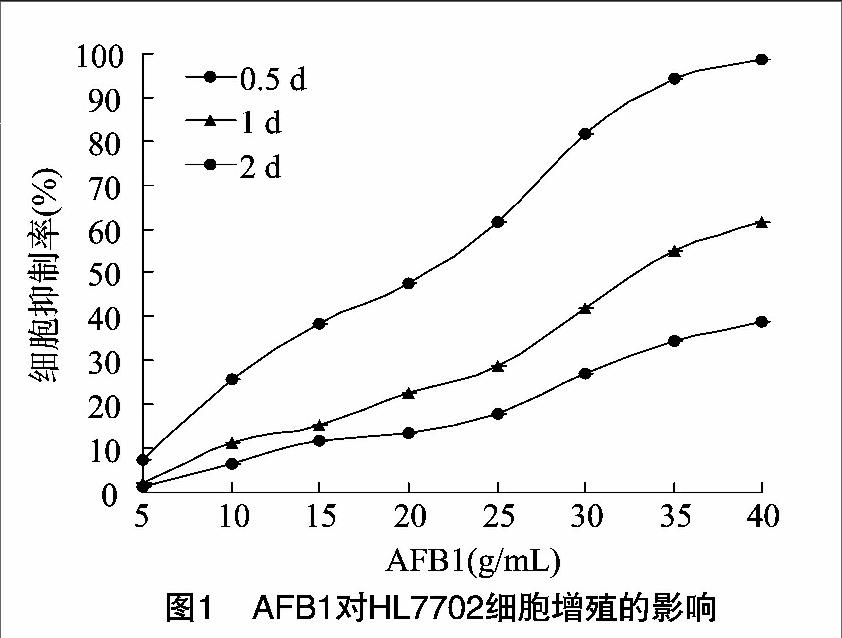
由圖1可見,AFB1對HL-7702的細胞毒性呈現出劑量和時間依賴性。0.5、1 d組各濃度雖也可以明顯抑制細胞增殖,但彼此之間差異不大,2 d組各濃度對應的細胞抑制率之間差異明顯,且濃度與抑制率之間相關性好(r2=0.99),因此以2 d組數據確定AFB1的IC50濃度為20 μg/mL,以 此 作 為AFB1對HL-7702細胞損傷模型的最佳濃度。
2.2 油茶籽殼總黃酮對AFB1損傷的HL-7702細胞存活率的影響
將未經油茶籽殼總黃酮預處理的HL-7702細胞暴露于20 μg/mL AFB1 2 d,與對照組相比,細胞存活率明顯下降(表2)。經25、50、100 μg/mL油茶籽殼總黃酮預保護過的HL-7702細胞再接受相同樣劑量AFB1暴露相同時間后,細胞存活率較AFB1損傷組明顯提高,且其保護效果呈顯著的劑量效應關系。油茶籽殼總黃酮對HL-7702細胞進行預處理可以有效抑制AFB1造成的細胞增殖抑制。
2.3 HL-7702細胞內ROS和MDA水平的改變
由表3可知,與對照組相比,20 μg/mL AFB1作用HL-7702細胞2 d,顯著增加了HL-7702細胞內ROS活性和MDA含量,說明AFB1造成了HL-7702細胞的氧化損傷。經25、50、100 μg/mL油茶籽殼總黃酮預處理后,受損細胞內的ROS活性逐漸降低,與AFB1損傷組相比差異顯著,MDA含量也隨著油茶籽殼總黃酮濃度升高而顯著下降。
2.4 HL-7702細胞內抗氧化酶系的改變
由表4可見,與對照組相比,20 μg/mL AFB1作用HL-7702細胞2 d,顯著降低細胞內抗氧化酶SOD、CAT、GSH-Px活性,說明細胞內源性抗氧化酶系統功能失調。經25、50、100 μg/mL油茶籽殼總黃酮預處理HL-7702細胞1 d,與AFB1損傷組相比,25 μg/mL保護組沒有顯著提升HL-7702細胞內CAT、GSH-Px活性。
2.5 HL-7702細胞內Nrf2及其下游抗氧化酶表達的改變
Nrf2被稱為氧化還原敏感轉錄因子,是調節抗氧化應激反應的重要轉錄因子[16]。與對照組相比,AFB1損傷組Nrf2及其下游GSTA2的mRNA(圖2-a)和蛋白表達水平(圖2-b)均明顯下調。隨著油茶籽殼總黃酮預保護濃度的提高,與AFB1損傷組相比,Nrf2及GSTA2的mRNA和蛋白表達水平都呈現劑量依賴性上調。GSTA2是Nrf2調控的下游眾多抗氧化蛋白之一,SOD、CAT、GSH-Px也受Nfr2的調控,由此可知,油茶籽殼總黃酮在細胞氧化應激的情況下,通過上調Nrf2表達,提高了其下游一系列抗氧化酶的活性,從而對抗由AFB1造成的細胞氧化損傷。
2.6 細胞內NF-κB及下游炎癥因子mRNA表達的改變
研究表明,肝細胞遭受氧化受損及肝炎、肝癌發生時NF-κB信號通路被顯著激活[17-18]。由圖3可知,與對照組相比,AFB1損傷組核轉錄因子NF-κB表達明顯上調,其相關下游炎癥介導因子TNF-α、IL-6、IL-1β也隨之高表達,隨著油茶籽殼總黃酮預保護濃度的提高,與AFB1損傷組相比,NF-κB及炎癥因子表達都呈現劑量依賴性下調。由此可知,油茶籽殼總黃酮對HL-7702細胞的預處理下調了由AFB1引起的NF-κB信號通路的激活,相關炎癥因子表達隨之減少,提示其在預防、治療肝炎方面可能具有的功能。
3 結論與討論
生物有氧代謝過程中產生的過多活性氧(ROS)會導致氧化應激,肝臟因含有豐富的線粒體,是產生ROS較多的器官,也是較易受到ROS攻擊的器官。氧化應激會引起脂質過氧化,破壞酶活性,損傷DNA,造成肝損傷。研究表明,氧化應激與多種肝臟疾病、肝功能障礙密切相關[19]。AFB1進入肝細胞后,通過環氧化反應轉變為有活性的AFB1-8,9環氧化合物,進而引發肝細胞內產生過量ROS,造成細胞的氧化應激,這是AFB1生物毒性尤其是肝臟毒性發生的主要機理[20]。
本研究表明,當肝細胞遭遇AFB1損傷時,油茶籽殼總黃酮與肝細胞的預孵顯著降低了細胞內ROS、MDA水平,同時提升了肝細胞內抗氧化酶系(SOD、CAT、GSH-Px)的活性,因此改善了肝細胞AFB1暴露下的氧化-抗氧化平衡環境,提高了細胞存活率。El-Nekeety 等[3]、 Zhang等[21]、Hou等[22]都曾報道過:當細胞暴露于霉菌毒素時,天然抗氧化劑可以增強細胞內抗氧化酶活性,阻斷脂質過氧化,從而緩解細胞的氧化應激。Nrf2是公認的細胞防御過氧化應激的重要調節因子,在氧化應激狀態下,Nrf2與細胞核內抗氧化反應元件ARE結合,上調一系列解毒酶和抗氧化蛋白表達,從而增強肝細胞的解毒及抗氧化能力,減輕活性氧(ROS)和親電子物質介導的細胞損害,對保護肝臟功能、阻止多種肝臟疾病的發生有著重要意義[23]。已經證明能通過Nrf2途徑改善肝細胞氧化應激狀態,進而發揮保護肝細胞作用的化合物有白藜蘆醇[24]、姜黃素[25]、齊墩果酸[26]、熊去氧膽酸[27] 等,這些化合物主要集中在對肝細胞的乙醇或其他藥物損傷方面的保護,由于這些化合物的來源十分有限,限制了其更為廣泛的應用。本研究表明,來自油茶籽殼中的總黃酮物質與肝細胞的預孵上調了Nrf2、GSTA2表達,GSTA2是Nrf2調控的下游眾多抗氧化蛋白之一,與肝臟解毒功能有關,SOD、CAT、GSH-Px也受Nfr2調控,抗氧化酶是細胞內對抗ROS的主要防御機制,因此可以認為油茶籽殼總黃酮預處理肝細胞后激活了Nfr2信號通路,上調其下游多種抗氧化酶基因的表達,提高細胞內抗氧化酶活性,從而對抗由ROS引起的肝細胞氧化損傷,起到保護肝細胞的作用。Costa等也報道了黃酮類物質在上調Nrf2蛋白和提高抗氧化酶活性方面的重要作用[28]。
細胞內產生過量ROS除了會導致MDA積累外,還可激活NF-κB信號通路,進而使其下游調控的許多炎癥因子(包括TNF-α等)表達增加,持續釋放,肝細胞氧化應激、凋亡,炎癥細胞大量浸潤,最終導致肝炎甚至引發肝癌[29] 。本研究表明,油茶籽殼總黃酮的預處理使得肝細胞NF-κB表達下降及其調控的炎癥因子(TNF-α、 IL-6、IL-1β)下調,這提示其在預防、治療肝炎方面可能具有的功能。油茶籽殼總黃酮通過上調Nrf2,提高Nfr2下游抗氧化酶系(SOD、CAT、GSH-Px、GSTA2)的活性,阻斷了由ROS引起的脂質過氧化鏈式反應,同時下調NF-κB,減少炎癥因子相關基因(TNF-α、IL-6、IL-1β)的表達,從而拮抗AFB1對肝細胞的毒性損傷,提高肝細胞存活率。油茶籽殼作為一種農業副產品,在我國資源豐富、價格低廉,本研究證明了其中的黃酮類物質具有拮抗AFB1肝細胞毒性的功能,為油茶籽殼的環保、高效利用提供了新途徑。
參考文獻:
[1]Rawal S,Kim J E,Coulombe R. Aflatoxin B1 in poultry:toxicology,metabolism and prevention[J]. Research in Veterinary Science,2010,89(3):325-331.
[2]Abdel-Aziem S H,Hassan A M,El-Denshary E S,et al. Ameliorative effects of thyme and calendula extracts alone or in combination against aflatoxins-induced oxidative stress and genotoxicity in rat liver[J]. Cytotechnology,2014,66(3):457-470.
[3]El-Nekeety A A,Mohamed S R,Hathout A S,et al. Antioxidant properties of Thymus vulgaris oil against aflatoxin-induce oxidative stress in male rats[J]. Toxicon,2011,57(7/8):984-991.
[4]Hassan A M,Abdel-Aziem S H,El-Nekeety A A,et al. Panax ginseng extract modulates oxidative stress,DNA fragmentation and up-regulate gene expression in rats sub chronically treated with aflatoxin B1 and fumonisin B1[J]. Cytotechnology,2015,67(5):861-871.
[5]El-Nekeety A A,Abdel-Azeim S H,Hassan A M,et al. Quercetin inhibits the cytotoxicity and oxidative stress in liver of rats fed aflatoxin-contaminated diet[J]. Toxicology Reports,2014,1:319-329.
[6]Guengerich F P,Kim D H. In vitro inhibition of dihydropyridine oxidation and aflatoxin B1 activation in human liver microsomes by naringenin and other flavonoids[J]. Carcinogenesis,1990,11(12):2275-2279.
[7]Farombi E O,Adepoju B F,Ola-Davies O E,et al. Chemoprevention of aflatoxin B1-induced genotoxicity and hepatic oxidative damage in rats by kolaviron,a natural bioflavonoid of Garcinia kola seeds[J]. European Journal of Cancer Prevention,2005,14(3):207-214.
[8]楊俊換,周建平,胡堯超. 油茶籽殼的化學成分分析與掃描電鏡觀察[J]. 湖南農業大學學報:自然科學版,2013,39(5):554-558.
[9]姜天甲,應鐵進,陳秋平,等. 油茶籽殼總黃酮的提取及抗氧化研究[J]. 中國食品學報,2010,10(1):93-99.
[10]楊彤暉,于修燭,蔡學標,等. 超聲波輔助提取油茶籽殼總黃酮及對豬油抗氧化能力研究[J]. 糧油食品科技,2012,20(1):26-29.
[11]高 揚,易若琨,宋家樂.竹葉總黃酮對UVB誘導 HaCaT細胞氧化損傷的保護作用[J]. 南京中醫藥大學學報,2015,31(2):165-169.
[12]Fraga C G,Leibovitz B E,Tappel A L. Lipid peroxidation measured as thiobarbituric acid-reactive substances in tissue slices:characterization and comparison with homogenates and microsomes[J]. Free Radical Biology & Medicine,1988,4(3):155-161.
[13]Zhong X L,Shi Y R,Chen J J,et al. Polyphenol extracts from Punica granatum and Terminalia chebula are anti-inflammatory and increase the survival rate of chickens challenged with Escherichia coli[J]. Biological & Pharmaceutical Bulletin,2014,37(10):1575-1582.
[14]He Y,Fang J,Peng X,et al. Effects of sodium selenite on aflatoxin B1-induced decrease of ileac T cell and the mRNA contents of IL-2,IL-6,and TNF-α in broilers[J]. Biological Trace Element Research,2014,159(1/2/3):167-173.
[15]Pan Z M,Fang Q,Geng S Z,et al. Analysis of immune-related gene expression in chicken peripheral blood mononuclear cells following Salmonella enterica serovar enteritidis infection in vitro[J]. Research in Veterinary Science,2012,93(2):716-720.
[16]李 平,李 芬,葉 昉,等. Nrf2信號通路在鉛致SH-SY5Y細胞氧化應激中作用[J]. 中國公共衛生,2012,28(7):933-935.
[17]于晨輝,杜仲燕,高 佳,等. 4-HNE通過抑制TNF-α介導的NF-κB活化誘導酒精性肝損傷[J]. 中國病理生理雜志,2013,29(6):1046-1052.
[18]李亞萍,黨雙鎖,翟 嵩,等. NF-κB、α-SMA及TGFβ1在實驗性肝損傷中的表達[J]. 臨床肝膽病雜志,2011,27(11):1207-1210.
[19]Cederbaum A I,Lu Y K,Wu D F. Role of oxidative stress in alcohol-induced liver injury[J]. Archives of Toxicology,2009,83(6):519-548.
[20]Beyki M,Zhaveh S,Khalili S T,et al. Encapsulation of mentha piperita essential oils in chitosan-cinnamic acid nanogel with enhanced antimicrobial activity against aspergillus flavus[J]. Industrial Crops and Products,2014,54(4):310-319.
[21]Zhang A H,Sun H,Wang X J. Recent advances in natural products from plants for treatment of liver diseases[J]. European Journal of Medicinal Chemistry,2013,63(33):570-577.
[22]Hou Y J,Zhao Y Y,Xiong B,et al. Mycotoxin-containing diet causes oxidative stress in the mouse[J]. PLoS One,2013,8(3):e60374.
[23]邵洛林,郭建強,許偉華. Nrf2在肝細胞中作用的研究進展[J]. 國際消化病雜志,2009,29(6):381-383,423.
[24]Rubiolo J A,Mithieux G,Vega F V. Resveratrol protects primary rat hepatocytes against oxidative stress damage:activation of the Nrf2 transcription factor and augmented activities of antioxidant enzymes[J]. European Journal of Pharmacology,2008,591(1/3):66-72.
[25]Farombi E O,Shrotriya S,Na H K,et al. Curcumin attenuates dimethylnitrosamine-induced liver injury in rats through Nrf2-mediated induction of heme oxygenase-1[J]. Food and Chemical Toxicology,2008,46(4):1279-1287.
[26]Reisman S A,Aleksunes L M,Klaassen C D. Oleanolic acid activates Nrf2 and protects from acetaminophen hepatotoxicity via Nrf2-dependent and Nrf2-independent processes[J]. Biochemical Pharmacology,2009,77(7):1273-1282.
[27]Kawata K,Kobayashi Y,Souda K,et al. Enhanced hepatic Nrf2 activation after ursodeoxycholic acid treatment in patients with primary biliary cirrhosis[J]. Antioxidants & Redox Signaling,2010,13(3):259-268.
[28]Costa S,Schwaiger S,Cervellati R,et al. In vitro evaluation of the chemoprotective action mechanisms of leontopodic acid against aflatoxin B1 and deoxynivalenol-induced cell damage[J]. Journal of Applied Toxicology,2009,29(1):7-14.
[29]Martindale J L,Holbrook N J. Cellular response to oxidative stress:signaling for suicide and survival[J]. Journal of Cellular Physiology,2002,192(1):1-15.姚 輝,楊悅君. 明清皇家陵寢的“祈福”文化與旅游開發——以清東陵為例[J]. 江蘇農業科學,2016,44(4):578-582.
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
