生物鐘基因Per1對(duì)人口腔鱗狀細(xì)胞癌細(xì)胞增殖、凋亡、細(xì)胞周期和體內(nèi)成瘤的影響及機(jī)理
2016-06-15 07:51:24付小娟楊凱李晗雪趙欽陳丹重慶醫(yī)科大學(xué)附屬第一醫(yī)院口腔頜面外科重慶400016
華西口腔醫(yī)學(xué)雜志 2016年3期
付小娟 楊凱 李晗雪 趙欽 陳丹重慶醫(yī)科大學(xué)附屬第一醫(yī)院口腔頜面外科,重慶 400016
?
生物鐘基因Per1對(duì)人口腔鱗狀細(xì)胞癌細(xì)胞增殖、凋亡、細(xì)胞周期和體內(nèi)成瘤的影響及機(jī)理
付小娟楊凱李晗雪趙欽陳丹
重慶醫(yī)科大學(xué)附屬第一醫(yī)院口腔頜面外科,重慶 400016
[摘要]目的探討生物鐘基因Per1對(duì)細(xì)胞周期相關(guān)基因的調(diào)控作用,以及對(duì)人口腔鱗狀細(xì)胞癌細(xì)胞SCC15增殖、凋亡、細(xì)胞周期和體內(nèi)成瘤的影響。方法構(gòu)建3組針對(duì)Per1 RNA的短發(fā)夾RNA(shRNA)慢病毒重組質(zhì)粒,轉(zhuǎn)染SCC15細(xì)胞,用蛋白質(zhì)印跡法(Western blot)及實(shí)時(shí)熒光定量PCR(qRT-PCR)檢測(cè)篩選RNA干擾作用最強(qiáng)組為實(shí)驗(yàn)組,對(duì)照組(Control-shRNA)為對(duì)任何基因均無干擾效應(yīng)的shRNA序列的質(zhì)粒,空白組為未做任何處理的SCC15細(xì)胞。用qRT-PCR檢測(cè)各組細(xì)胞中細(xì)胞周期相關(guān)基因Per1、p53、Cyclin D1、Cyclin E、Cyclin A2、Cyclin B1、CDK1、CDK2、CDK4、CDK6、p16、p21、Wee1、cdc25、E2F、Rb1 mRNA的表達(dá);用流式細(xì)胞儀檢測(cè)各組細(xì)胞增殖、凋亡和細(xì)胞周期分布;將實(shí)驗(yàn)組和空白組細(xì)胞分別接種于裸鼠背部皮下,觀察成瘤情況。結(jié)果成功構(gòu)建了3組Per1-shRNA慢病毒質(zhì)粒,通過qRT-PCR和Western blot證明Per1-shRNA-Ⅰ組沉默效果最佳,作為實(shí)驗(yàn)組。Per1-shRNA-Ⅰ組中Cyclin D1、Cyclin E、Cyclin B1、CDK1和Wee1 mRNA的表達(dá)水平高于Control-shRNA組和SCC15組(P<0.05),p53、Cyclin A2、p16、p21和cdc25 mRNA的表達(dá)水平降低(P<0.05);Control-shRNA組和SCC15組中各基因mRNA的表達(dá)水平無差異(P>0.05)。CDK2、CDK4、CDK6、E2F和Rb1 mRNA的表達(dá)水平在3組中均無統(tǒng)計(jì)學(xué)差異(P>0.05)。Per1-shRNA-Ⅰ組中細(xì)胞增殖指數(shù)高于Control-shRNA組和SCC15組(P<0.05),凋亡指數(shù)降低(P<0.05),S期的細(xì)胞數(shù)降低(P<0.05),G2/M期的細(xì)胞數(shù)增加(P<0.05)。Control-shRNA組和SCC15組細(xì)胞的增殖指數(shù)和凋亡指數(shù)無統(tǒng)計(jì)學(xué)差異(P>0.05)。Per1-shRNA-Ⅰ組細(xì)胞體內(nèi)成瘤能力顯著增強(qiáng)(P<0.05)。結(jié)論生物鐘基因Perl是重要的抑癌基因,Perl能調(diào)控下游眾多的細(xì)胞周期基因,其表達(dá)變化影響細(xì)胞周期進(jìn)程、增殖和凋亡的平衡失調(diào)及體內(nèi)成瘤能力,對(duì)Per1深入研究有可能進(jìn)一步明確癌癥的發(fā)生發(fā)展機(jī)制,為癌癥的治療提供新的有效分子靶點(diǎn)。
[關(guān)鍵詞]生物鐘;Per1;細(xì)胞周期;基因;口腔癌
Correspondence: Yang Kai, E-mail: cqfyyk@aliyun.com.
生物體內(nèi)的許多生命活動(dòng),如激素分泌、細(xì)胞增殖等,均表現(xiàn)出近24 h的周期波動(dòng),稱為晝夜節(jié)律[1-3]。晝夜節(jié)律是生物適應(yīng)外界環(huán)境長期進(jìn)化的結(jié)果,是生命活動(dòng)的基本特征之一。晝夜節(jié)律的產(chǎn)生由細(xì)胞內(nèi)生物鐘基因呈晝夜節(jié)律性表達(dá)所導(dǎo)致[1-4]。目前人們已發(fā)現(xiàn)14個(gè)生物鐘基因:Per1、Per2、Per3、Cryl、Cry2、Clock、Bmal1、TIM、CK1ε、NPAS2,REV-ERBs、Dec1、Dec2、RORs[1,3-4],這些生物鐘基因通過周期性表達(dá)產(chǎn)生的晝夜節(jié)律使復(fù)雜的生命活動(dòng)相互協(xié)調(diào)有序。Per1是重要的生物鐘基因,具有維持晝夜節(jié)律穩(wěn)定和調(diào)控晝夜節(jié)律周期的作用[5-6]。研究表明:Per1不僅與生物體晝夜節(jié)律有關(guān),而且還能調(diào)控下游許多重要的細(xì)胞周期蛋白[7],Per1的異常表達(dá)與癌癥的發(fā)生發(fā)展密切相關(guān)[7-9]。
細(xì)胞周期紊亂是腫瘤發(fā)生的主要原因[4,10]。正常細(xì)胞周期的運(yùn)行依賴于細(xì)胞周期蛋白(Cyclins)、細(xì)胞周期蛋白依賴性激酶(cyclin dependent kinases,CDKs)和細(xì)胞周期蛋白依賴性激酶抑制因子(cyclin dependent kinase inhibitors,CKIs)組成的細(xì)胞周期分子網(wǎng)絡(luò)調(diào)控系統(tǒng)[4,10-11]。Cyclins包括Cyclin A-Y,其中起主要作用的有Cyclin A2、B1、D1、E;CDKs包括CDK1-CDK16,起重要作用的有CDK1、2、4、6;CKIs包括Ink4家族和Cip/Kip家族,其中起重要作用的主要有Ink4家族中p16和Cip/Kip家族中p21[10-11]。在Cyclins-CDKs-CKIs網(wǎng)絡(luò)系統(tǒng)中,CDKs是調(diào)控細(xì)胞周期的核心,Cyclins和CKIs分別對(duì)CDKs具有正向調(diào)控作用和負(fù)向調(diào)控作用[10-11]。
研究表明:生物鐘基因Per1在多種癌癥中表達(dá)降低[8-9],Per1能調(diào)控下游許多重要的細(xì)胞周期蛋白[7],從而導(dǎo)致細(xì)胞周期進(jìn)程和細(xì)胞增殖水平改變,與癌癥發(fā)生發(fā)展密切相關(guān)[4,7,10]。在Cyclins-CDKs-CKIs網(wǎng)絡(luò)系統(tǒng)中,人們的研究主要集中在Per1對(duì)細(xì)胞周期蛋白的調(diào)控,如Cyclin B1、Cyclin D1、Cyclin E和Wee1等[7],但關(guān)于Per1對(duì)CDKs和CKIs中分子的調(diào)控還了解甚少。為了進(jìn)一步探討Per1基因與癌癥發(fā)生的關(guān)系以及晝夜節(jié)律與細(xì)胞周期兩大周期活動(dòng)之間的相互作用,本研究通過對(duì)人口腔鱗狀細(xì)胞癌細(xì)胞內(nèi)的Per1基因沉默,檢測(cè)Per1沉默后細(xì)胞的細(xì)胞周期、增殖、凋亡和體內(nèi)成瘤能力的改變,并全面檢測(cè)Cyclins-CDKs-CKIs網(wǎng)絡(luò)系統(tǒng)中各重要分子的改變情況,以闡明Per1與癌癥發(fā)生的機(jī)制。
1 材料和方法
1.1細(xì)胞和主要試劑
人口腔鱗狀細(xì)胞癌SCC15細(xì)胞(重慶醫(yī)科大學(xué)生命科學(xué)院),慢病毒表達(dá)載體PLKO.1(Sigma公司,美國),質(zhì)粒提取試劑盒(Invitrogen公司,美國),逆轉(zhuǎn)錄試劑盒(TaKaRa公司,日本);逆轉(zhuǎn)錄聚合酶鏈反應(yīng)(reverse transcription polymerase chainreaction,RT-PCR)試劑盒(Bio-Rad公司,美國),BCA蛋白濃度測(cè)定試劑盒、鼠抗人甘油醛-3-磷酸脫氫酶( glyceraldehyde-3-phosphate dehydrogenase,GAPDH),單克隆抗體(上海碧云天生物技術(shù)有限公司),兔抗人Per1單克隆抗體(Genetex公司,美國),辣根酶標(biāo)記山羊抗兔IgG抗體(北京中杉金橋生物技術(shù)有限公司),低溫冷凍離心機(jī)(Z223-MK-Z,HERMLE公司,德國),熒光定量PCR儀(C1000TMThermal Cycler,Bio-Rad公司,美國),核酸蛋白分析儀(UV-GeneQuant,安瑪西亞公司,瑞典),流式細(xì)胞儀(FACSVantage SE,BD公司,美國)。
1.2Per1短發(fā)夾RNA(Per1-short hairpin RNA,Per1-shRNA)慢病毒干擾質(zhì)粒載體構(gòu)建和鑒定
根據(jù)Genbank數(shù)據(jù)庫中人的Per1 mRNA序列(GeneID: NM_002616),遵循RNA干擾序列設(shè)計(jì)原則[12],選擇Per1 RNA干擾靶序列后進(jìn)行BLAST基因組同源性分析(www.ncbi.nlm.nih.gov/BLAST/),確定3個(gè)干擾作用位點(diǎn)為本實(shí)驗(yàn)Per1基因的靶序列,用軟件BLOCK-iT? RNAi Designer(Invitrogen公司,美國)設(shè)計(jì)3條Per1 RNA干擾序列和1條對(duì)照序列,Per1-Ⅰ:3’-CAGCACCACTAAGCGTAAATG-5’;Per1-Ⅱ:3’-CCAGCACCACTAAGCGTAAAT-5’;Per1-Ⅲ:3’-CCATGGACATGTCCACCTATA-5’;對(duì)照序列:3’-CCTAAGGTTAAGTCGCCCTCG-5’。通過Genbank數(shù)據(jù)庫驗(yàn)證該對(duì)照序列對(duì)任何基因均無干擾效應(yīng)。在DNA連接酶的作用下將各條序列5’末端添加CCGG序列,3’末端添加TTTTTG序列,各條序列的正義鏈與反義鏈之間用Loop環(huán)(CTCGAG)連接,然后在5’和3’末端用DNA連接酶分別連接含限制性內(nèi)切酶Age Ⅰ和EcoR Ⅰ酶切位點(diǎn)的黏性末端。將上述合成的shRNA行PCR擴(kuò)增,用限制性內(nèi)切酶Age Ⅰ和EcoR Ⅰ對(duì)慢病毒載體PLKO.1進(jìn)行雙酶切,得到的產(chǎn)物經(jīng)瓊脂糖電泳分離、割膠回收和純化,然后與shRNA片段在DNA連接酶作用下連接形成Per1-shRNA-PLKO.1。將連接產(chǎn)物轉(zhuǎn)化感受態(tài)大腸桿菌DH5α后涂布于含氨基核苷類抗生素的LB固體培養(yǎng)基上抗性培養(yǎng),將形成的單克隆菌落接種于LB培養(yǎng)液中,37 ℃搖菌過夜,離心(8 000 r·min-1,3 min)后收集細(xì)菌并裂解細(xì)胞,提取質(zhì)粒DNA,然后用Age Ⅰ和EcoR Ⅰ行雙酶切后用Chromas軟件(Technelysium公司,澳大利亞)測(cè)序鑒定。
1.3Per1-shRNA慢病毒質(zhì)粒包裝及轉(zhuǎn)染SCC15細(xì)胞
取對(duì)數(shù)生長期的293T細(xì)胞,以2×106個(gè)·mL-1的細(xì)胞數(shù)接種于直徑為10 cm的培養(yǎng)皿中,放于37 ℃、5% CO2飽和濕度的培養(yǎng)箱中培養(yǎng),在細(xì)胞生長飽和度達(dá)到70%~80%時(shí),加入各質(zhì)粒DNA溶液10 μg和20 μL Lipofectamine 2000脂質(zhì)體進(jìn)行共轉(zhuǎn)染,48 h后收集293T細(xì)胞上清液,離心10 min(4 ℃,4 000 r·min-1)后用直徑0.45 μm的濾器過濾上清液,即獲得慢病毒包裝的Per1-shRNA重組質(zhì)粒病毒液。取對(duì)數(shù)生長期的SCC15細(xì)胞,接種于6孔板中,加入4 mL血清新鮮培養(yǎng)基、10 μL Polybrene和1 mL重組質(zhì)粒病毒液,混勻培養(yǎng)24 h,再向各孔中加入嘌呤霉素(終濃度2.0 μg·mL-1)進(jìn)行傳代篩選。實(shí)驗(yàn)分為5組:Per1-shRNA-Ⅰ組、Per1-shRNA-Ⅱ組和Per1-shRNA-Ⅲ組分別為轉(zhuǎn)染Per1-shRNA-Ⅰ、Per1-shRNA-Ⅱ和Per1-shRNA-Ⅲ質(zhì)粒病毒的SCC15細(xì)胞,對(duì)照組(ControlshRNA)為轉(zhuǎn)染Control-shRNA質(zhì)粒(對(duì)照質(zhì)粒,即只含對(duì)任何基因均無干擾效應(yīng)的shRNA序列的質(zhì)粒)的SCC15細(xì)胞,SCC15組為未作任何處理的SCC15細(xì)胞(空白對(duì)照)。
1.4蛋白質(zhì)印跡法(Western blot)檢測(cè)
收集各組細(xì)胞,用RIPA裂解液提取總蛋白,用BCA蛋白濃度測(cè)定試劑盒測(cè)定蛋白質(zhì)濃度。將提取蛋白與其1/4體積的蛋白上樣緩沖液混合后變性5 min (100 ℃)。取50 μg蛋白上樣,在10%十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)100 V電壓下電泳,所得電泳產(chǎn)物轉(zhuǎn)移至PVDF膜上,用5%脫脂奶粉封閉1 h后加入一抗兔抗人Per1多克隆抗體(1︰200)和鼠抗人GAPDH單克隆抗體(1︰3 000)過夜,用TBST(Tris Buffered Saline with Tween-20)液洗膜3次,每次10 min,然后加入二抗為辣根酶標(biāo)記山羊抗兔IgG抗體(1︰1 000),37 ℃孵育1 h。用TBST液洗膜3遍后,電化學(xué)發(fā)光(electrochemiluminescence,ECL)顯影。最后用Quantity One軟件(Bio-Rad公司,美國)分析條帶灰度值,計(jì)算 PER1和內(nèi)參GAPDH條帶灰度值比值。以上實(shí)驗(yàn)重復(fù)3次。
1.5實(shí)時(shí)熒光定量PCR(quantitative real-time PCR, qRT-PCR)檢測(cè)
按Trizol說明書分別提取各組細(xì)胞的總RNA,將總RNA溶解于焦碳酸二乙酯(diethylpyrocarbonate,DEPC)處理的雙蒸水中,用紫外分光光度儀測(cè)定RNA濃度和純度。用逆轉(zhuǎn)錄試劑盒將RNA反轉(zhuǎn)錄為cDNA,按試劑盒說明進(jìn)行操作,反應(yīng)體系為10 μL,反應(yīng)條件:37 ℃,15 min,85 ℃ 5 s,4 ℃ 4 min。用Primer Premier 5.0軟件設(shè)計(jì)并合成目的基因Per1、p53、Cyclin D1、Cyclin E、Cyclin A2、Cyclin B1、CDK1、CDK2、CDK4、CDK6、p16、p21、Wee1、cdc25、E2F、Rb1和內(nèi)參基因GAPDH的引物序列(表1)。用熒光定量PCR儀進(jìn)行PCR擴(kuò)增,反應(yīng)體系為10 μL:Premix Ex TapTM5 μL,上下游引物各0.5 μL,50 ng·μL-1cDNA模板1 μL,滅菌雙蒸水3 μL補(bǔ)足體積至10 μL。擴(kuò)增參數(shù)為:95 ℃預(yù)變性3 min,95 ℃變性10 s,60 ℃退火延伸30 s,擴(kuò)增40個(gè)循環(huán)。每次qRT-PCR反應(yīng)都設(shè)有以相同體積的滅菌雙蒸水取代cDNA作為空白對(duì)照,每個(gè)樣本設(shè)3個(gè)復(fù)孔,檢測(cè)重復(fù)3次。用Pfaffl法[13]計(jì)算各基因mRNA的表達(dá)。

表1 各基因qRT-PCR引物序列Tab 1 The primers for qRT-PCR of the genes
1.6流式細(xì)胞儀檢測(cè)
根據(jù)Per1 mRNA和蛋白檢測(cè)結(jié)果,篩選RNA干擾作用最強(qiáng)的Per1-shRNA-Ⅰ組作為實(shí)驗(yàn)組。取對(duì)數(shù)生長期的Per1-shRNA-Ⅰ、Control-shRNA和SCC15組細(xì)胞,用0.25%胰蛋白酶消化后吹打成單細(xì)胞懸液,調(diào)整各組細(xì)胞濃度為1×106個(gè)·mL-1,分別取1 mL各組細(xì)胞懸液離心(1 000 r·min-1,4 ℃)5 min,去掉上清液,PBS液洗滌細(xì)胞2次。
細(xì)胞周期分布和細(xì)胞增殖檢測(cè):在各組細(xì)胞中加入0.5 mL70%乙醇(-20 ℃預(yù)冷)固定,4 ℃冰箱過夜,離心(1 000 r·min-1,4 ℃)5 min,用PBS液洗滌細(xì)胞2遍,加入碘化丙啶染色液1 mL,4 ℃避光30 min后用流式細(xì)胞儀檢測(cè)細(xì)胞周期分布,并計(jì)算細(xì)胞增殖指數(shù)(proliferation index,PI),PI=(S+ G2/M)/(G0/G1+S+G2/M)×100%。
細(xì)胞凋亡檢測(cè):在各組細(xì)胞中加入200 μL膜聯(lián)蛋白(Annexin)Ⅴ-異硫氰酸熒光素(fluorescein isothiocyanate,F(xiàn)ITC)染色液,4 ℃避光孵育15 min,再加入碘化丙啶染色液1 mL混勻,5 min后用流式細(xì)胞儀檢測(cè)細(xì)胞凋亡。細(xì)胞凋亡指數(shù)(apoptosis index,AI)=(凋亡細(xì)胞數(shù)/所測(cè)細(xì)胞總數(shù))×100%。實(shí)驗(yàn)重復(fù)3次。
1.7體內(nèi)成瘤實(shí)驗(yàn)
無特定病原體(specific pathogen free,SPF)級(jí)別BALB/c nu/nu品系雌性裸鼠10只,4~6周齡,體重18~22 g(重慶醫(yī)科大學(xué)實(shí)驗(yàn)動(dòng)物研究所)。隨機(jī)分為Per1-shRNA-Ⅰ組(實(shí)驗(yàn)組)和SCC15組(空白組),每組5只。采用0.25%胰酶分別消化對(duì)數(shù)生長期的Per1-shRNA-Ⅰ和SCC15細(xì)胞,冷凍低速離心(4 ℃,1 000 r·min-1)5 min后,用PBS液調(diào)整細(xì)胞濃度為5×106個(gè)·mL-1的單細(xì)胞懸液。將0.2 mL Per1-shRNA-Ⅰ和SCC15細(xì)胞懸液分別注入實(shí)驗(yàn)組和空白組裸鼠的右背部皮下。3周明顯成瘤后,用頸椎脫位法處死裸鼠,取出腫瘤,用電子天平(A250型,Denver公司,美國)稱重量,用游標(biāo)卡尺測(cè)量腫瘤的最大長徑(a)和最小短徑(b),按公式計(jì)算腫瘤體積(V):V=0.5×a×b2。然后將腫瘤用4%多聚甲醛固定,常規(guī)石蠟包埋,切片,常規(guī)蘇木素-伊紅(hematoxylin-eosin,HE)染色后在200倍光學(xué)顯微鏡下觀察。所有實(shí)驗(yàn)操作程序均經(jīng)過重慶醫(yī)科大學(xué)實(shí)驗(yàn)動(dòng)物研究所實(shí)驗(yàn)動(dòng)物使用管理委員會(huì)批準(zhǔn)。
1.8統(tǒng)計(jì)學(xué)分析
采用SPSS 17.0軟件對(duì)3組及以上實(shí)驗(yàn)數(shù)據(jù)進(jìn)行單因素方差分析,組間兩兩比較采用 LSD-t檢驗(yàn);對(duì)體內(nèi)成瘤2組實(shí)驗(yàn)數(shù)據(jù)進(jìn)行t檢驗(yàn)。P<0.05為有統(tǒng)計(jì)學(xué)差異。
2 結(jié)果
2.1Per1-shRNA載體構(gòu)建及鑒定
測(cè)序鑒定表明,3條Per1-shRNA-Ⅰ~Ⅲ重組質(zhì)粒DNA測(cè)序與設(shè)計(jì)的干擾靶序列片段一致(圖1),表明3條Per1-shRNA(Ⅰ~Ⅲ)-PLKO.1重組慢病毒載體構(gòu)建成功。
圖1 Per1-shRNA重組質(zhì)粒測(cè)序圖譜Fig 1 The sequencing atlas of Per1-shRNA recombinant plasmids
2.2Per1基因沉默后SCC15細(xì)胞中Per1 mRNA和蛋白的表達(dá)
Per1-shRNA-Ⅰ~Ⅲ、Control-shRNA和SCC15組細(xì)胞中Per1 mRNA的表達(dá)分別為1.00±0.00、1.45± 0.34、1.50±0.23、2.11±0.31、2.20±0.34,Per1蛋白的灰度值比值分別為1.05±0.21、1.50±0.15、1.63± 0.11、2.18±0.24、2.21±0.33(圖2)。Per1-shRNA-Ⅰ組細(xì)胞中Per1 mRNA和蛋白的表達(dá)均低于其他組(P<0.05),表明Per1-shRNA-Ⅰ組的Per1沉默效果最佳,作為實(shí)驗(yàn)組。
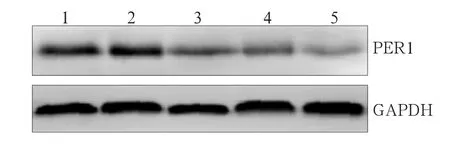
圖2 Western blot檢測(cè)各組Per1蛋白表達(dá)水平Fig 2 Per1 protein levels of each group analyzed by Western blot
2.3Per1沉默后SCC15細(xì)胞內(nèi)細(xì)胞周期相關(guān)基因mRNA的表達(dá)
qRT-PCR檢測(cè)結(jié)果見表2。Per1-shRNA-Ⅰ組中Cyclin D1、Cyclin E、Cyclin B1、CDK1和Wee1 mRNA的表達(dá)水平高于Control-shRNA組和SCC15組(P<0.05),p53、Cyclin A2、p16、p21和cdc25 mRNA的表達(dá)水平降低(P<0.05);Control-shRNA組和SCC15組中各基因mRNA的表達(dá)水平無差異(P>0.05)。CDK2、CDK4、CDK6、E2F和Rb1 mRNA的表達(dá)水平在3組中均無統(tǒng)計(jì)學(xué)差異(P>0.05)。
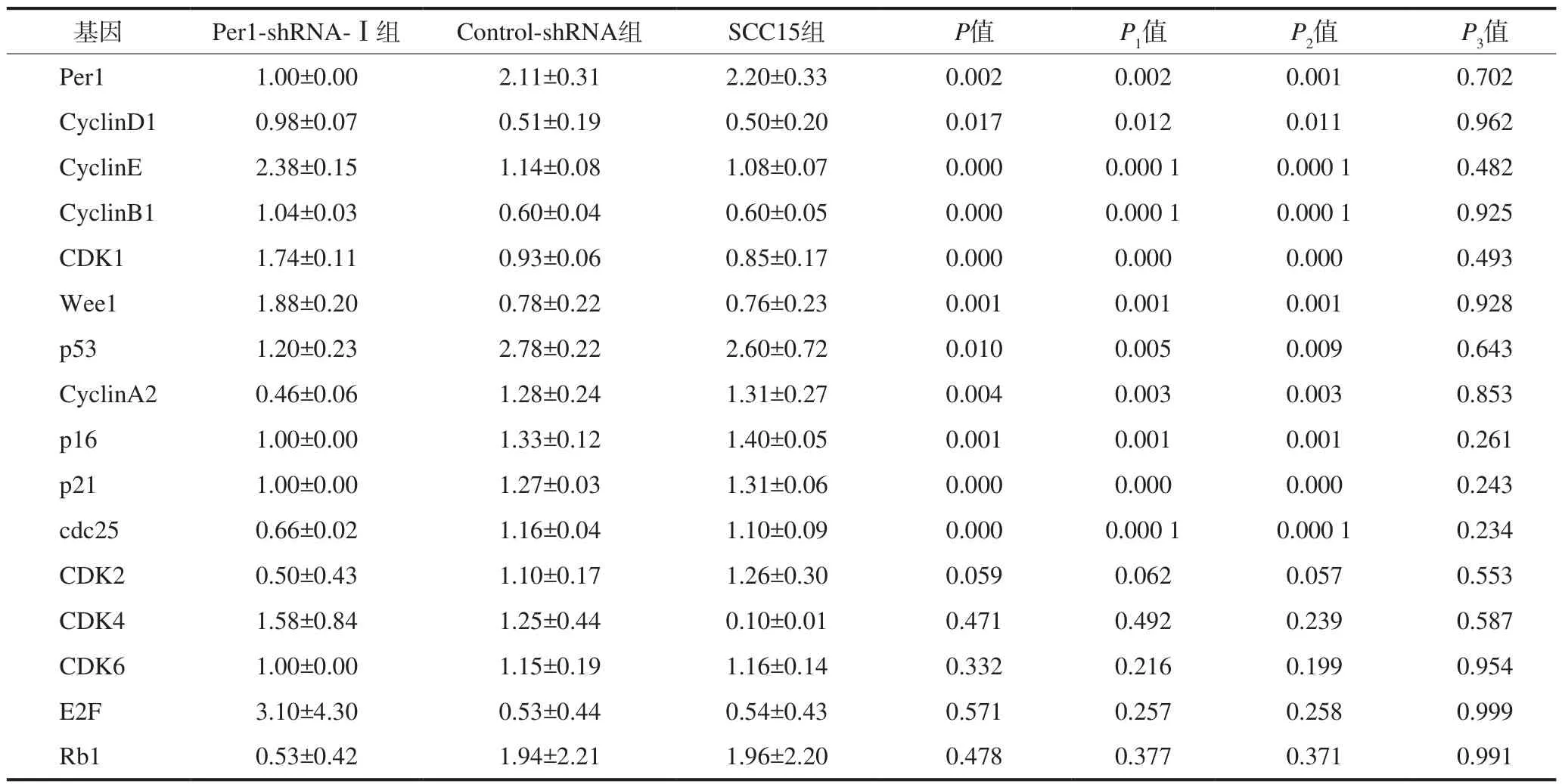
表2 Per1-shRNA-Ⅰ、Control-shRNA和SCC15組中細(xì)胞周期相關(guān)基因mRNA的表達(dá)Tab 2 The mRNA expressions of cell cycle related genes in Per1-shRNA-Ⅰ, Control-shRNA, and SCC15 groups
2.4Per1基因沉默后SCC15細(xì)胞的細(xì)胞周期分布、增殖、凋亡的變化
流式細(xì)胞儀檢測(cè)結(jié)果見表3。Per1-shRNA-Ⅰ組中細(xì)胞增殖指數(shù)高于Control-shRNA組和SCC15組(P<0.05),凋亡指數(shù)降低(P<0.05),S期細(xì)胞數(shù)降低(P<0.05),G2/M期細(xì)胞數(shù)增加(P<0.05)。Control-shRNA組和SCC15組細(xì)胞的增殖指數(shù)、凋亡指數(shù)和細(xì)胞周期分布無統(tǒng)計(jì)學(xué)差異(P>0.05)。
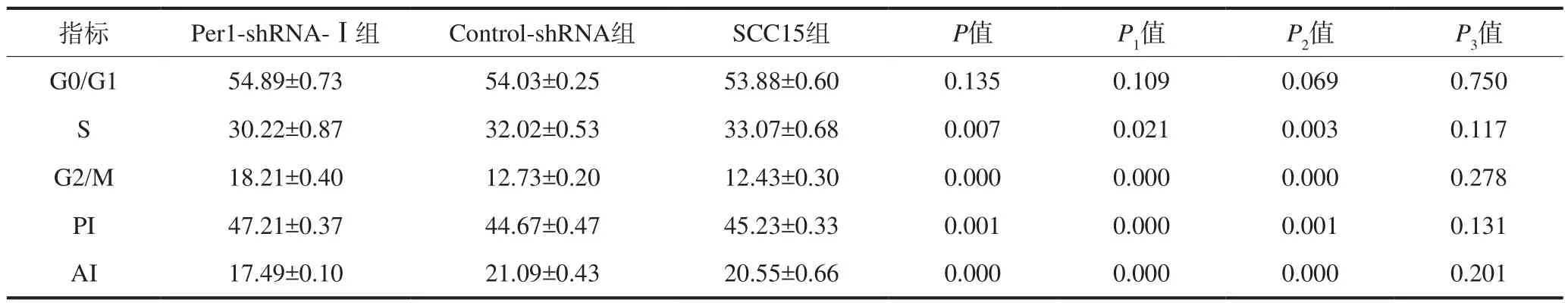
表3 Per1-shRNA-Ⅰ、Control-shRNA和SCC15組中細(xì)胞周期分布、細(xì)胞增殖、細(xì)胞凋亡Tab3 The cell cycle distribution, cell proliferation, and apoptosis in Per1-shRNA-Ⅰ, Control-shRNA, and SCC15 groups
2.5體內(nèi)成瘤
體內(nèi)成瘤結(jié)果見圖3:Per1-shRNA-Ⅰ組和SCC15組腫瘤的體積分別為(0.25±0.09)cm3和(0.08± 0.02)cm3,質(zhì)量分別為(0.50±0.16)g和(0.18± 0.04)g,均具有統(tǒng)計(jì)學(xué)差異(P<0.05)。表明Per1-shRNA-Ⅰ組細(xì)胞體內(nèi)成瘤能力顯著增強(qiáng)(P<0.05)。
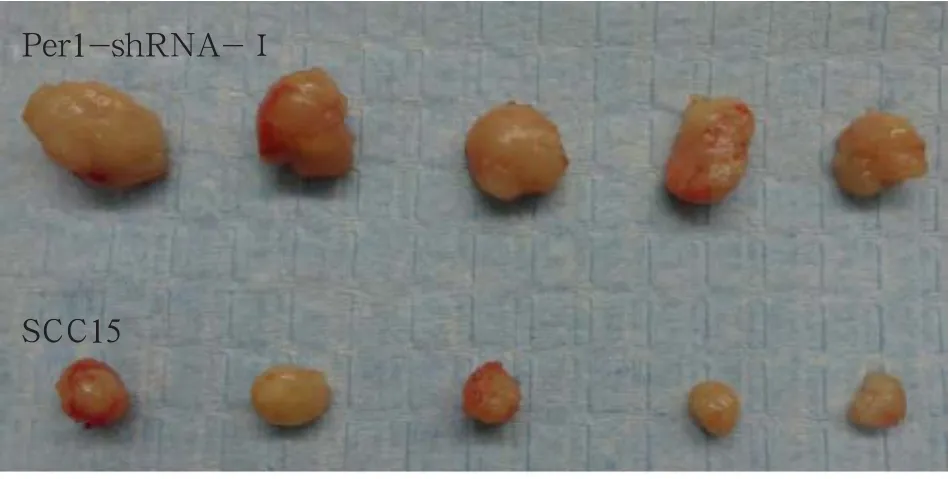
圖3 Per1-shRNA-Ⅰ和SCC15組細(xì)胞的體內(nèi)成瘤能力Fig 3 The tumorigenic ability in vivo of Per1-shRNA-Ⅰand SCC15 group cells
3 討論
細(xì)胞周期紊亂導(dǎo)致細(xì)胞增殖和凋亡的平衡改變,這是腫瘤發(fā)生的主要原因[4,10]。正常細(xì)胞在Cyclins-CDKs-CKIs細(xì)胞周期分子網(wǎng)絡(luò)系統(tǒng)的精確調(diào)控下按嚴(yán)格的時(shí)間順序沿著G1-S-G2-M時(shí)相運(yùn)轉(zhuǎn)[4,10,14]。在Cyclins-CDKs-CKIs網(wǎng)絡(luò)中,CDKs是調(diào)控細(xì)胞周期的核心,在細(xì)胞周期的不同時(shí)相,不同的Cyclins與不同的CDKs結(jié)合形成Cyclins/CDKs復(fù)合物,激活CDKs的功能,促進(jìn)細(xì)胞由一個(gè)細(xì)胞周期時(shí)相向另一個(gè)細(xì)胞周期時(shí)相有序轉(zhuǎn)換,而不同的CKIs可通過與相應(yīng)的CDKs或Cyclins/CDKs復(fù)合物結(jié)合,抑制CDKs的活性,從而抑制細(xì)胞周期時(shí)相轉(zhuǎn)換[10-11]。如果正性調(diào)節(jié)因子Cyclins和負(fù)性調(diào)節(jié)因子CKIs的協(xié)同調(diào)控作用發(fā)生異常,細(xì)胞增殖和凋亡的平衡就會(huì)發(fā)生改變,最終導(dǎo)致腫瘤的發(fā)生。
研究[7-8]證明:Per1異常表達(dá)與腫瘤的發(fā)生發(fā)展密切相關(guān),其機(jī)理是Per1能調(diào)控下游許多重要細(xì)胞周期蛋白。Yang等[7]報(bào)道,下調(diào)Per1基因表達(dá)導(dǎo)致小鼠乳腺癌細(xì)胞中Cyclin D1和Cyclin E的表達(dá)水平增高,細(xì)胞增殖及生長能力增加;過表達(dá)Per1導(dǎo)致結(jié)腸癌細(xì)胞HCT116中Cyclin B1和Wee1的表達(dá)水平降低,從而誘導(dǎo)細(xì)胞凋亡,抑制細(xì)胞增殖;Cao等[8]也發(fā)現(xiàn),過表達(dá)Per1基因能抑制前列腺癌細(xì)胞增殖和促進(jìn)細(xì)胞凋亡。但在Cyclins-CDKs-CKIs細(xì)胞周期分子網(wǎng)絡(luò)系統(tǒng)中,Per1對(duì)CDKs和CKIs這兩方面的調(diào)控作用目前還了解甚少。
G1期的啟動(dòng)是啟動(dòng)細(xì)胞周期運(yùn)行的關(guān)鍵,Cyclin D1與CDK4或CDK6結(jié)合,激活CDK4或CDK6的活性,啟動(dòng)G1期,然后Cyclin E與CDK2結(jié)合,激活CDK2的活性,使細(xì)胞從G1期向S期過渡[10,14],同時(shí)CKIs中屬于Ink4家族的p16能與Cyclin D1競(jìng)爭(zhēng)與CDK4 或CDK6結(jié)合,特異性抑制CDK4或CDK6活性,而CKIs中屬于Cip/Kip家族的p21具有廣泛激酶抑制活性,能抑制各種Cyclins-CDKs復(fù)合物的活性,因而在G1期對(duì)CyclinE/CDK2復(fù)合體活性有抑制作用,從而阻滯細(xì)胞周期進(jìn)程[10-11,15]。本研究顯示:SCC15細(xì)胞中沉默Per1基因后,雖然CDK2、CDK4和CDK6的表達(dá)水平無顯著改變,但Cyclin D1和Cyclin E表達(dá)顯著增高,激活了CDK2、CDK4或CDK6的活性,同時(shí)p16和p21表達(dá)顯著降低,對(duì)CDK2、CDK4和CDK6活性的抑制能力減弱,從而增強(qiáng)了細(xì)胞G1期的啟動(dòng)能力,促進(jìn)細(xì)胞由G1期向S期加速過渡,促進(jìn)細(xì)胞增殖。本研究也證明:Per1基因沉默后,SCC15細(xì)胞體內(nèi)成瘤能力增強(qiáng)。
在細(xì)胞周期的S期,Cyclin A2與CDK2結(jié)合激活CDK2,Cyclin A2/CDK2復(fù)合物驅(qū)動(dòng)細(xì)胞延續(xù)S期,并促進(jìn)細(xì)胞從S期向G2期轉(zhuǎn)化[10,14]。本研究顯示:沉默Per1基因后,SCC15細(xì)胞中CDK2表達(dá)無明顯改變,但Cyclin A2表達(dá)顯著降低,同時(shí)抑制Cyclin A2/ CDK2復(fù)合物活性的p21表達(dá)也顯著降低,導(dǎo)致Cyclin A2/CDK2復(fù)合物對(duì)細(xì)胞S期的延續(xù)作用減弱,因而S期進(jìn)程縮短,從而導(dǎo)致S期細(xì)胞數(shù)降低。
在G2期,Cyclin B1與CDK1結(jié)合激活CDK1,CDK1/Cyclin B1復(fù)合體驅(qū)動(dòng)細(xì)胞從G2期向M期轉(zhuǎn)化。cdc25能增強(qiáng)CDK1的活性,促進(jìn)G2期向M期的轉(zhuǎn)化進(jìn)程[10,14],Weel能抑制CDK1的活性,導(dǎo)致M期延遲或?qū)⒓?xì)胞周期阻滯在G2/M期[10]。本研究顯示:沉默Per1基因后,SCC15細(xì)胞中CDK1、Cyclin B1和Wee1表達(dá)顯著增高,而p21和cdc25表達(dá)顯著降低,提示沉默Per1基因后,Cyclins-CDKs-CKIs網(wǎng)絡(luò)調(diào)控系統(tǒng)中的三方面均發(fā)生了改變。同時(shí)本研究結(jié)果還顯示,細(xì)胞在G2/M期數(shù)量增多,證明Wee1對(duì)Cyclin B1/CDK1復(fù)合體的負(fù)性調(diào)控作用占有優(yōu)勢(shì)。
在Cyclins-CDKs-CKIs網(wǎng)絡(luò)調(diào)控的細(xì)胞周期運(yùn)行中,p53是G1/S細(xì)胞周期檢查點(diǎn)的重要因子。在G1/S細(xì)胞周期檢查點(diǎn),有DNA損傷的細(xì)胞通過激活p53使細(xì)胞周期停滯進(jìn)行DNA修復(fù),不能修復(fù)者誘導(dǎo)其凋亡[10]。本研究結(jié)果顯示:沉默Per1基因后SCC15細(xì)胞中p53表達(dá)降低,導(dǎo)致細(xì)胞對(duì)損傷DNA的修復(fù)能力減弱和誘導(dǎo)細(xì)胞凋亡能力下降,增加了受損DNA未經(jīng)修復(fù)的細(xì)胞進(jìn)入S期,使細(xì)胞基因組的完整性和穩(wěn)定性受到影響,促進(jìn)細(xì)胞惡性轉(zhuǎn)化。本研究也證明:Per1基因沉默后,SCC15細(xì)胞增殖指數(shù)顯著增高,凋亡指數(shù)顯著下降,并且體內(nèi)成瘤能力增強(qiáng)。
本研究證明:生物鐘基因Per1是重要的抑癌基因。Per1沉默后,SCC15癌細(xì)胞中CDK2、CDK4、CDK6、E2F和Rb1 mRNA表達(dá)水平無顯著改變,但Cyclin D1、Cyclin E、Cyclin B1、CDK1和Wee1 mRNA的表達(dá)水平顯著增高,而p53、Cyclin A2、p16、p21和cdc25 mRNA的表達(dá)水平顯著降低,最終導(dǎo)致細(xì)胞增殖水平增高,凋亡水平下降,細(xì)胞周期進(jìn)程改變,細(xì)胞體內(nèi)成瘤能力增強(qiáng)。本研究從轉(zhuǎn)錄水平證明了生物鐘基因Per1對(duì)Cyclins-CDKs-CKIs細(xì)胞周期分子網(wǎng)絡(luò)中的三方面及細(xì)胞周期G1/S檢查點(diǎn)均有重要的調(diào)控作用。在此基礎(chǔ)上對(duì)Per1的深入研究有可能進(jìn)一步明確晝夜節(jié)律與細(xì)胞周期兩大周期活動(dòng)之間的相互作用以及與癌變發(fā)生的關(guān)系,為癌癥的治療提供新的有效分子靶點(diǎn)。
[參考文獻(xiàn)]
[1]Kelleher FC, Rao A, Maguire A.Circadian molecular clocks and cancer[J].Cancer Lett, 2014, 342(1):9-18.
[2]Eismann EA, Lush E, Sephton SE.Circadian effects in cancerrelevant psychoneuroendocrine and immune pathways[J].Psychoneuroendocrinology, 2010, 35(7):963-976.
[3]Zieker D, Jenne I, Koenigsrainer I, et al.Circadian expression of clock- and tumor suppressor genes in human oral mucosa[J].Cell Physiol Biochem, 2010, 26(2):155-166.
[4]Borgs L, Beukelaers P, Vandenbosch R, et al.Cell circadian cycle: new role for mammalian core clock genes[J].Cell Cycle, 2009, 8(6):832-837.
[5]Zheng B, Albrecht U, Kaasik K, et al.Nonredundant roles of the mPer1 and mPer2 genes in the mammalian circadian clock[J].Cell, 2001, 105(5):683-694.
[6]Bae K, Jin X, Maywood ES, et al.Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock[J].Neuron, 2001, 30(2):525-536.
[7]Yang X, Wood PA, Ansell CM, et al.The circadian clock gene Per1 suppresses cancer cell proliferation and tumor growth at specific times of day[J].Chronobiol Int, 2009, 26(7):1323-1339.
[8]Cao Q, Gery S, Dashti A, et al.A role for the clock gene per1 in prostate cancer[J].Cancer Res, 2009, 69(19):7619-7625.
[9]Cadenas C, van de Sandt L, Edlund K, et al.Loss of circadian clock gene expression is associated with tumor progression in breast cancer[J].Cell Cycle, 2014, 13(20):3282-3291.
[10]Soták M, Sumová A, Pácha J.Cross-talk between the circadian clock and the cell cycle in cancer[J].Ann Med, 2014, 46(4):221-232.
[11]Lim S, Kaldis P.Cdks, cyclins and CKIs: roles beyond cell cycle regulation[J].Development, 2013, 140(15):3079-3093.
[12]Wilson RC, Doudna JA.Molecular mechanisms of RNA interference[J].Annu Rev Biophys, 2013, 42:217-239.
[13]Pfaffl MW.A new mathematical model for relative quantification in real-time RT-PCR[J].Nucleic Acids Res, 2001, 29(9):e45.
[14]Hochegger H, Takeda S, Hunt T.Cyclin-dependent kinases and cell-cycle transitions: does one fit all[J].Nat Rev Mol Cell Biol, 2008, 9(11):910-916.
[15]Williams RT, Barnhill LM, Kuo HH, et al.Chimeras of p14ARF and p16: functional hybrids with the ability to arrest growth[J].PLoS ONE, 2014, 9(2):e88219.
(本文編輯李彩)
·臨床研究·
Effects and mechanism of the circadian clock gene Per1 on the proliferation, apoptosis, cycle, and tumorigenicity in vivo of human oral squamous cell carcinoma
Fu Xiaojuan, Yang Kai, Li Hanxue, Zhao Qin, Chen Dan.(Dept.of Oral and Maxillofacial Surgery, The First Affiliated Hospital of Chongqing Medical University, Chongqing 400016, China)
[Key words]circadian clock;Per1;cell cycle;gene;oral carcinoma
[Abstract]Objective To determine the regulatory effects of the circadian clock gene Per1 on cell cycle-related genes and its influence on the proliferation, apoptosis, cycle, and tumorigenicity in vivo of human oral squamous cell carcinoma SCC15 cells.MethodsThree groups of the short hairpin RNA (shRNA) of lentivirus recombinant plasmids were designed against the RNA of Per1 and then transfected to the SCC15 cells.The optimum interference group was screened through Western blot and quantitative real-time PCR (qRT-PCR) and assigned as the experimental group.The transfected lentivirus plasmid without an interference effect on any gene was set as the control group (Control-shRNA).Untreated SCC15 cells were set as the blank group.The mRNA expressions of cell cycle-related genes, namely, Per1, p53, Cyclin D1, Cyclin E, Cyclin A2, Cyclin B1, CDK1, CDK2, CDK4, CDK6, p16, p21, Wee1, cdc25, E2F, and Rb1, in each group were detectedthrough qRT-PCR.The cell proliferation, apoptosis, and cell cycle distribution in each group were evaluated through flow cytometry.The cells of the experimental group andthe blank group were subcutaneously inoculated in nude mice to observe tumorigenesis.ResultsThree groups of Per1-shRNA lentivirus plasmids were constructed successfully.Among the groups, the Per1-shRNA-Ⅰgroup exhibited the highest interference effect, as indicated by qRT-PCR and Western blot analysis.As such, this group was set as the experimental group.The mRNA expression levels of CyclinD1, CyclinE, CyclinB1, CDK1, and Wee1 gene in the Per1-shRNA-Ⅰgroup were significantly higher than those in the Control-shRNA group and the SCC15 group (P<0.05).By contrast, the mRNA expression levels of p53, Cyclin A2, p16, p21, and cdc25 in the Per1-shRNA-Ⅰgroup were significantly lower than those in the Control-shRNA group and the SCC15 group (P<0.05).The mRNA expression levels of each gene between the Control-shRNA group and the SCC15 group did not significantly differ (P>0.05).The mRNA expression levels of CDK2, CDK4, CDK6, E2F, and Rb1 did not significantly differed in the three groups (P>0.05).The proliferation index of the Per1-shRNA-Ⅰgroup was significantly higher than those of the Control-shRNA group and the SCC15 group (P<0.05).The apoptosis index of the Per1-shRNA-Ⅰgroup was significantly lower than those of the Control-shRNA group and the SCC15 group (P<0.05).The number of S-phase cells in the Per1-shRNA-Ⅰgroup was significantly lower than those of S-phase cells in the Control-shRNA group and the SCC15 group (P<0.05).The number of G2/M-phase cells in the Per1-shRNA-Ⅰgroup was significantly higher than those of G2/M-phase cells in the Control-shRNA group and the SCC15 group (P<0.05).Conversely, the proliferation index, apoptotic index, and cell cycle distribution of the cells in the Control-shRNA group did not significantly differ from those of the SCC15 group (P>0.05).The tumorigenic ability in vivo was significantly enhanced in the Per1-shRNA-Ⅰgroup (P<0.05).ConclusionPer1 is an important tumor suppressor gene.Perl can regulate a large number of downstream cell cycle-related genes.The alteration of its expression can affect cell cycle progression, proliferation, apoptosis imbalance, and tumorigenic ability in vivo.Further studies on Per1 may elucidate cancer development and provide novel effective molecular targets for cancer treatment.
[中圖分類號(hào)]R 739.8
[文獻(xiàn)標(biāo)志碼]A [doi]10.7518/hxkq.2016.03.008
[收稿日期]2015-10-16; [修回日期]2016-02-28
[作者簡(jiǎn)介]付小娟,碩士,E-mail:466133758@qq.com
[通信作者]楊凱,教授,博士,E-mail:cqfyyk@aliyun.com

- 華西口腔醫(yī)學(xué)雜志的其它文章
- 一種顯微精準(zhǔn)定深孔牙體預(yù)備技術(shù)
- 牙齦卟啉單胞菌合成環(huán)二腺苷酸的高效液相色譜-串聯(lián)質(zhì)譜法定性分析
- 變異鏈球菌磷酸轉(zhuǎn)移酶系統(tǒng)ptxA、ptxB基因?qū)?xì)菌生長能力的影響
- 針對(duì)變異鏈球菌的人源特異性靶向抗菌肽C16LL-37的生物學(xué)特性
- 脈沖超聲及電磁場(chǎng)對(duì)大鼠骨髓間充質(zhì)干細(xì)胞體外成軟骨分化過程中細(xì)胞外基質(zhì)溢出的影響
- 降鈣素基因相關(guān)肽通過Hippo通路調(diào)控小鼠骨髓間充質(zhì)干細(xì)胞成骨分化的實(shí)驗(yàn)研究