MRSA對巨噬細胞系RAW264.7自噬的影響
2016-06-20 02:58:34朱軍吳本權楊洋朱家馨張天托
新醫學 2016年5期
關鍵詞:研究
朱軍 吳本權 楊洋 朱家馨 張天托
510630廣州,中山大學附屬第三醫院呼吸內科,內科ICU
?
MRSA對巨噬細胞系RAW264.7自噬的影響
朱軍吳本權楊洋朱家馨張天托
510630廣州,中山大學附屬第三醫院呼吸內科,內科ICU
【摘要】目的探討耐甲氧西林金黃色葡萄球菌(MRSA)感染對于巨噬細胞自噬的影響。方法以小鼠單核巨噬細胞系RAW264.7為研究對象,以革蘭陽性菌MRSA感染3 h組作為實驗組,設立空白對照組(對照組),另設MRSA感染0、30、60、120、150、180 min組。以MRSA感染細胞后,熒光定量PCR檢測自噬相關基因5(ATG5)、ATG7、ATG12的表達情況,蛋白免疫印跡法檢測自噬相關蛋白Beclin1及微管相關蛋白輕鏈3(LC3)的表達情況。結果感染MRSA 3 h后,實驗組RAW264.7細胞中ATG5、ATG7、ATG12的mRNA表達水平均高于對照組(P均<0.01),LC3蛋白相對表達量也比對照組增加(P<0.01),2組間Beclin1蛋白相對表達量比較差異無統計學意義(P>0.05)。ATG7、ATG12 mRNA和LC3及Beclin1蛋白相對表達量均在感染MRSA 180 min組最高,ATG5 mRNA在感染MRSA 150 min組最高。結論MRSA可以誘導巨噬細胞發生自噬,自噬基因激活存在先后順序,自噬可能為巨噬細胞吞噬胞外菌MRSA的另一種免疫滅活形式。
【關鍵詞】耐甲氧西林金黃色葡萄球菌;巨噬細胞;自噬
自噬是與真核生物關系密切的基礎生理過程,通過細胞自我消化累積的蛋白質細胞器等各類復雜的細胞質成分,從而調節控制真核生物細胞內成分的數量與質量。現今,已有自噬與各種疾病的關系研究[1]。隨著近年研究的逐步深入,巨噬細胞的自噬對于清除胞內病原體有何重要作用逐漸受到關注[2-3]。自噬的作用機制在肺部疾病中,尤其在COPD、特發性肺纖維化、肺結核等疾病中已有較透徹的研究[4]。關于自噬與肺部感染性疾病,如胞外菌感染的研究仍較少。近年,耐甲氧西林金黃色葡萄球菌(MRSA)已成為引起醫院內和社區相關性感染的重要致病菌之一,其治療形式較為嚴峻[5]。已有研究發現,自噬可以清除某些胞內菌,如李斯特菌、沙門菌和結核分枝桿菌[6]。對于胞外菌MRSA感染細胞,巨噬細胞除吞噬外能否產生自噬的相關研究仍較少。本研究以小鼠單核巨噬細胞系RAW264.7為對象,觀察自噬在MRSA肺部感染中的作用,旨在探討MRSA感染巨噬細胞對其自噬功能的影響。
材料與方法
一、材料
1. 細胞系及菌株
小鼠單核巨噬細胞系RAW264.7購于中國醫學科學院基礎醫學研究所基礎醫學細胞中心;MRSA菌株為由我院呼吸科實驗室分離鑒定的臨床菌株。
2. 主要試劑及儀器
DMEM高糖培養基、胎牛血清、青霉素和鏈霉素購于美國Life公司,Syber Green Mix實時定量PCR 檢測試劑盒購于美國Life公司。LC3多克隆兔抗體購于美國Sigma公司;Beclin1單克隆抗體購于美國Cell Signaling Technology公司,內參抗體β-actin兔一抗購于中國谷歌生物公司,辣根過氧化物酶(HRP) 標記的羊抗兔二抗購于廣州威佳科技有限公司,增強化學發光(ECL)檢測試劑盒購于廣州聚研生物科技有限公司,細胞培養6孔板購于美國Corning公司。凱基全蛋白提取試劑盒及凱基BCA法蛋白濃度檢測試劑盒購于廣州杰特偉生物科技有限公司,蛋白質印跡法配膠成分30%聚丙烯酰胺、三羥甲基氨基甲烷-鹽酸(Tris-HCl)緩沖液、10%十二烷基硫酸鈉(SDS)溶液、過硫酸銨(APS)購于廣州威佳科技有限公司,蛋白電泳轉膜系統購于美國Bio-Rad公司。
二、方法
1. 細胞培養
RAW264.7細胞用含10%胎牛血清、100 kU/L青霉素和100 mg/L鏈霉素的DMEM高糖培養基培養于25 ml的透氣蓋培養瓶內,將培養瓶置于37℃、5%CO2培養箱內,根據細胞數目形態,當細胞融合至60%~70%時將細胞傳代,平均約2~3 d傳代1次,保持實驗用細胞處于對數生長期,細胞狀態良好,在普通光學顯微鏡下折光度透亮,未見大量顆粒及細胞碎片。
2. 細菌培養
將凍存于-80℃的MRSA菌種在室溫下解凍后,接種于血瓊脂培養基,置于37℃溫箱培養24 h,然后測定其在600 nm處吸光度(OD600) ,當 OD600=0.6,即約1×109/ml時,置于16%甘油在-80℃凍存。
3. MRSA感染細胞
將處于對數生長期的RAW264.7細胞按1×106/ml的密度接種于6孔板,每孔加入含10%胎牛血清的DMEM高糖培養液2 ml,培養過夜,待每孔細胞數增至2×106時開始感染,根據預實驗結果,按細菌∶細胞=10∶1將MRSA加入培養板中感染細胞。第一部分的實驗分為2組:空白處理作為對照組,MRSA感染作為實驗組。第二部分的實驗為MRSA刺激時間點檢測:將細胞均勻種于6孔板中,共分7組,第1組為對照組、第2組加入MRSA作用30 min、第3組加入MRSA作用60 min,組間相隔30 min,以此類推,第7組MRSA作用時間為180 min。每組實驗重復3次,取均值作為結果。
4. ATG mRNA表達水平的檢測
采用熒光定量PCR(qPCR)法:根據Gen Bank數據庫的ATG基因序列,分別設計ATG5、ATG7、ATG12特異性引物,由美國Life公司合成,引物序列見表1。收集上述各組細胞,在6孔板每孔加入1 ml Trizol提取細胞總RNA并進行模板DNA第一鏈合成,然后加入10 μl Syber Green混合物,反應體系總體積20 μl,采用ABI 7500系統進行PCR反應。反應條件為:50℃ 2 min,95℃ 2 min預變性,95℃15 s變性,60℃1 min退火,共40個循環,95℃15 s,60℃1 min,95℃30 s,60℃15 s延伸。反應后得到溶解曲線,記錄目的基因與內參基因達到某一閾值的Ct值,將目的基因與內參基因Ct值進行比較后再與對照組比較,計算ATG mRNA相對表達量:ATGmRNA=2-ΔΔCt。
5. 自噬相關蛋白Beclin1及微管相關蛋白輕鏈3(LC3)表達水平的檢測
MRSA感染細胞后,收集細胞加入現配的蛋白裂解液(每孔50 μl),用細胞刮刮取蛋白,收集混合液于4℃以12 000 r/min離心15 min,收集上清液,于562 nm波長下用二喹啉甲酸(BCA)法檢測蛋白濃度,加入5倍體積蛋白上樣緩沖液混勻,煮沸10 min,-80℃保存。采用十二烷基硫酸鈉聚丙烯酰胺(SDS-PAGE)凝膠電泳法,下層12%分離膠,上層5%濃縮膠,蛋白上樣量20 μg,85 V恒壓電泳,待蛋白標記物分出條帶后,將電壓調為110 V,當電泳至條帶接近底端時,停止電泳,260 mA恒流轉膜1.5 h將蛋白轉至聚偏氟乙烯(PVDF)膜上,根據預染蛋白標記物,剪下不同的目的蛋白于濕盒內孵育,加入5%脫脂奶粉室溫封閉1 h,TBST緩沖液洗膜3次,每次洗10 min,一抗稀釋比例LC3為1∶1 000、Beclin1為1∶2 000、β-actin為1∶2 000,4℃過夜后,TBST緩沖液洗膜3次,每次10 min,室溫孵育二抗1 h(稀釋比例為1∶2 000) ,TBST緩沖液洗膜3次,每次15 mim,ECL發光液A液、B液等體積混合后敷于蛋白膜上,于Chemi Scope化學發光成像系統顯影,運用Image J軟件分析LC3(取LC-Ⅰ條帶)及Beclin1的蛋白灰度值,作為其蛋白相對表達量。

表1 引物序列
三、 統計學處理
結果
一、感染MRSA 對RAW264.7細胞中ATG表達的影響
感染MRSA 3 h后,實驗組RAW264.7細胞的ATG5、ATG7、ATG12的mRNA表達水平均高于對照組(P均<0.01),見表1。
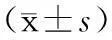
表1 感染MRSA 3 h后RAW264.7細胞中
二、感染MRSA不同時間點RAW264.7細胞中的ATG表達情況
感染MRSA不同時間點組的RAW264.7細胞中,ATG5、ATG7、ATG12 mRNA表達水平比較差異均有統計學意義(F分別為36.551、41.073、19.662,P均<0.001)。其中ATG7、ATG12 mRNA相對表達量在感染MRSA 180 min組最高,ATG5 mRNA相對表達量在感染MRSA 150 min組最高。180 min組的ATG7 mRNA表達水平為4.07±0.54,高于90 min組的2.08±0.01(t=9.025,P<0.001);180 min組的ATG12 mRNA表達水平為3.54±0.46,高于90 min組的1.66±0.19(t=9.253,P<0.001);150 min組的ATG5 mRNA表達水平為2.49±0.14,高于90 min組的1.83±0.25(t=2.650,P=0.024),見圖1。

圖1 感染MRSA不同時間點后
三、 感染MRSA對RAW264.7細胞中LC3及Beclin1蛋白表達的影響
感染MRSA 3 h后,RAW264.7細胞中LC3蛋白相對表達量在實驗組為48 523±12 692、對照組為7 450±1 550,2組比較差異有統計學意義(t=14.692,P<0.001); Beclin1蛋白相對表達量在實驗組為13 765±2 267、對照組為12 522±3 347,2組比較差異無統計學意義(t=0.533,P=0.619),見圖2。
四、感染MRSA不同時間點RAW264.7細胞中LC3及Belin1蛋白的表達情況
感染MRSA不同時間點組的RAW264.7細胞中,LC3及Belin1蛋白相對表達量比較差異均有統計學意義(F分別為5.235、3.938,P均<0.05)。LC3、Beclin1蛋白相對表達量均在感染MRSA 180 min組最高。其中LC3蛋白相對表達量在180 min組為57 010±3 978,在60 min組為45 383±6 273,2組比較差異有統計學意義(t=3.210,P=0.009)。Beclin1蛋白相對表達量在180 min組為86 440±5 024、60 min組為70 685±2 776,2組比較差異有統計學意義(t=6.273,P<0.001),見圖3。

圖2 感染MRSA后RAW264.7細胞中LC3及Beclin1的表達情況
A:蛋白免疫印跡法結果;B:定量分析結果

圖3 感染MRSA不同時間點RAW264.7細胞中LC3及Belin1蛋白的表達情況
A:蛋白免疫印跡法結果;B:定量分析結果
討論
巨噬細胞是固有免疫系統重要組成部分,在外來病原體入侵機體時,可以通過吞噬等非特異性免疫作用來殺傷病原體,并且可以通過抗原提呈誘導特異性免疫進一步殺死病原體。肺部感染性疾病中,巨噬細胞對于肺部的宿主防御功能必不可少,其能夠監視暴露的氣道,調節固有和適應性免疫[7]。
自噬是真核生物細胞內回收利用細胞質成分的一種降解機制。自噬過程包括將長壽命蛋白及胞內較大體積的細胞器成分隔離進入雙層膜囊泡狀結構,然后再運送至溶酶體進行降解,降解后的成分重新用來供應細胞能量及合成大分子的需要。因此,自噬被視為胞內回收機制[8]。最早對于自噬分子機制的研究從酵母菌開始,隨后發現大多數自噬基因出現于高等真核生物,表明自噬是一種進化高度保守的過程。在酵母菌中,自噬主要涉及適應饑餓,在多細胞生物體內,該途徑涉及多條通路,例如程序性細胞死亡、胞內破損細胞器及沉積蛋白質的清除等[9]。從整體上而言,自噬與多種病理生理過程相關,包括腫瘤、神經退行性病變、感染等[10]。隨著研究的逐步深入,學者們發現自噬與疾病的治療相關,已有研究證明自噬與腫瘤治療相關,可以提高腫瘤細胞對化學治療藥物的敏感性[11]。與哺乳動物自噬相關的基因統一命名為ATG。在哺乳動物細胞自噬泡形成過程中,由ATG3、ATG5、ATG7、ATG10、ATG12和LC3參與組成的2條泛素樣蛋白加工修飾通路,在自噬過程起著至關重要的作用。LC3是酵母ATG8的類似物,當LC3前體形成后,首先加工成胞漿可溶性形式LC3-I,并暴露出其羧基末端的甘氨酸殘基,然后LC3-I被ATG7活化,轉移并被修飾成膜結合形式LC3-Ⅱ[12-13]。本實驗中LC3-Ⅰ較LC3-Ⅱ表達明顯增多,考慮是否由于自噬屬于動態過程,LC3-Ⅱ已逐漸被溶酶體降解,故取LC3-Ⅰ條帶作定量分析。
近幾年,關于自噬與感染性疾病的研究逐漸增多[14]。金黃色葡萄球菌是可以引起人體從表皮感染至嚴重血源性感染等侵襲性感染的胞外菌,其感染率和患者病死率極高[15]。其中MRSA感染更為當前臨床治療的難題。雖然經典的研究證實,金黃色葡萄球菌是胞外菌,但其可逃避吞噬細胞的殺傷作用而存活于多種吞噬細胞中。更為嚴重的是,有研究顯示其可存活于單核細胞、巨噬細胞、甚至中性粒細胞中[16]。β-內酰胺類和糖肽類抗菌藥均難以進入上述免疫細胞,故治療失敗率高。因此,研究自噬對于胞內生存的MRSA有何作用及其與吞噬的關系,對闡明胞外病原體滅活及其免疫逃避機制均有重要意義。
已有研究表明,在肺泡上皮細胞A549受到結核分枝桿菌、A型鏈球菌和銅綠假單胞菌感染時,自噬可以清除病原體,對機體起到清除病原體的保護作用[17]。另外,自噬對于結核分枝桿菌亦具有殺傷作用[18]。目前,對金黃色葡萄球菌入侵單核細胞系MEF及Hela 細胞系產生自噬已有相關研究[19]。本研究主要觀察MRSA感染巨噬細胞對自噬的影響,結果顯示巨噬細胞在感染MRSA后,ATG5、ATG7、ATG12 mRNA及LC3蛋白相對表達升高,從基因水平及蛋白質水平證明感染MRSA可以激活巨噬細胞自噬,參與自噬過程的相關基因表達水平達峰時間不完全一致,表明自噬是一動態過程,基因激活存在先后順序。但對于該類自噬的具體信號通路機制仍然未明,其是否與革蘭陰性菌內毒素成分脂多糖誘導自噬過程類似,還是通過MRSA表面細胞壁某種成分誘導產生自噬,對此仍需要進一步研究[20]。另外, MRSA是否能夠逃避自噬,自噬能否起到免疫作用,對肺炎預后有何影響,還需進一步實驗研究[21]。
參考文獻
[1]Deretic V, Levine B.Autophagy, immunity, and microbial adaptations.Cell Host Microbe,2009,5(6):527-549.
[2]Chow A, Brown BD, Merad M.Studying the mononuclear phagocyte system in the molecular age.Nat Rev Immunol,2011,11(11):788-798.
[3]Araya J, Hara H, Kuwano K.Autophagy in the pathogenesis of pulmonary disease.Intern Med,2013,52(20):2295-2303.
[4]Weiss G, Schaible UE.Macrophage defense mechanisms against intracellular bacteria.Immunol Rev,2015,264(1):182-203.
[5]Otto M. MRSA virulence and spread.Cell Microbiol,2012,14(10):1513-1521.
[6]Huang J, Brumell JH. Autophagy in immunity against intracellular bacteria.Curr Top Microbiol Immunol,2009,335:189-215.
[7]Vlahos R, Bozinovski S.Role of alveolar macrophages in chronic obstructive pulmonary disease.Front Immunol,2014,5:435.
[8]Glick D, Barth S, Macleod KF.Autophagy: cellular and molecular mechanisms.J Pathol,2010,221(1):3-12.
[9]He C, Klionsky DJ.Regulation mechanisms and signaling pathways of autophagy.Annu Rev Genet,2009,43:67-93.
[10]Bialik S, Kimchi A.Autophagy and tumor suppression: recent advances in understanding the link between autophagic cell deathpathways and tumor development.Adv Exp Med Biol,2008,615:177-200.
[11]劉穗玲,李小毛. 自噬與紫杉醇敏感性. 新醫學,2014,45(4):218-222.
[12]Tal MC, Iwasaki A.Autophagy and innate recognition systems.Curr Top Microbiol Immunol,2009,335:107-121.
[13]Ryter SW, Cloonan SM, Choi AM.Autophagy: a critical regulator of cellular metabolism and homeostasis.Mol Cells,2013,36(1):7-16.
[14]Smeekens SP, Malireddi RK, Plantinga TS, Buffen K, Oosting M, Joosten LA, Kullberg BJ, Perfect JR, Scott WK, van de Veerdonk FL, Xavier RJ, van de Vosse E, Kanneganti TD, Johnson MD, Netea MG.Autophagy is redundant for the host defense against systemic Candida albicans infections.Eur J Clin Microbiol Infect Dis,2014,33(5):711-722.
[15]O’Keeffe KM, Wilk MM, Leech JM, Murphy AG, Laabei M, Monk IR, Massey RC, Lindsay JA, Foster TJ, Geoghegan JA, McLoughlin RM.Manipulation of autophagy in phagocytes facilitates Staphylococcus aureus bloodstream infection.Infect Immun,2015,83(9):3445-3457.
[16]Garzoni C, Kelley WL.Staphylococcus aureus: new evidence for intracellular persistence.Trends Microbiol,2009,17(2):59-65.
[17]Li P, Shi J, He Q, Hu Q, Wang YY, Zhang LJ, Chan WT, Chen WX.Streptococcus pneumoniae induces autophagy through the inhibition of the PI3K-I/Akt/mTOR pathway and ROS hypergeneration in A549 cells.PLoS One,2015,10(3):e0122753.
[18]Ní Cheallaigh C, Keane J, Lavelle EC, Hope JC, Harris J.Autophagy in the immune response to tuberculosis: clinical perspectives.Clin Exp Immunol,2011,164(3):291-300.
[19]Schnaith A, Kashkar H, Leggio SA, Addicks K, Kr?nke M, Krut O.Staphylococcus aureus subvert autophagy for induction of caspase-independent host cell death.J Biol Chem,2007,282(4):2695-2706.
[20]Xu Y, Jagannath C, Liu XD, Sharafkhaneh A, Kolodziejska KE, Eissa NT.Toll-like receptor 4 is a sensor for autophagy associated with innate immunity.Immunity,2007,27(1):135-144.
[21]Mizushima N, Levine B, Cuervo AM, Klionsky DJ.Autophagy fights disease through cellular self-digestion.Nature,2008,451(7182):1069-1075.
Effect of MRSA on the autophagy of RAW264.7 macrophage cell line
ZhuJun,WuBenquan,YangYang,ZhuJiaqxin,ZhangTiantuo.
DepartmentofRespiratory,ICU,theThirdAffiliatedHospitalofSunYat-senUniversity,Guangzhou510630,ChinaCorrespondingauthor,WuBenquan,E-mail:zswbq@163.com
【Abstract】ObjectiveTo evaluate the effect of methicillin-resistant staphylococcus aureus (MRSA) infection upon the autophagy of macrophages. MethodsMouse monocyte-macrophage cell line RAW264.7 was used. In the experimental groups, RAW264.7 cells were infected with Gram-positive MRSA for different time points including 0, 30, 60, 90, 120, 150 and 180 min. In the control group, the cells were left untreated. After MRSA infection, the expression of ATG5, ATG7 and ATG12 was measured by fluorescence quantitative PCR, and the expression levels of autophagy-related protein (Beclin1) and microtubule-associated protein light chain 3 (LC3) were measured by Western blot. ResultsAt 3 h after MRSA infection, the expression levels of ATG5, ATG7 and ATG12 mRNA in the experimental group were significantly higher compared with those in the control group (all P<0.01). The relative expression of LC3 in the experimental group was considerably higher than that in the control group (P<0.01), where the relative expression of Beclin1 did not significantly differ between two groups (P>0.05). The relative expression levels of ATG7, ATG12 mRNA, LC3 and Beclin1 proteins peaked at 180 min following MRSA infection, and the level of ATG5 mRNA achieved the peak at 150 min after MRSA infection. ConclusionsMRSA could induce autophagy in macrophages. The autophagy genes were activated in a certain sequence. Autophagy probably acted as an alternative immune process of phagocytosis of MRSA by macrophages.
【Key words】Methicillin-resistant staphylococcus aureus; Macrophage; Autophagy
DOI:10.3969/j.issn.0253-9802.2016.05.008
基金項目:國家自然科學基金面上項目(81170004)
(收稿日期:2015-11-22)(本文編輯:林燕薇)
·基礎研究論著·
通訊作者,吳本權,E-mail:zswbq@163.com
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
