黏著斑激酶信號通路在細菌侵入非吞噬細胞中的作用
2016-06-23 13:49:52賈曉陽付玉王瀟楠王志鋼郝慧芳
生物技術通報 2016年7期
賈曉陽 付玉 王瀟楠 王志鋼 郝慧芳
(內蒙古大學生命科學學院微生物實驗室,呼和浩特 010021)
黏著斑激酶信號通路在細菌侵入非吞噬細胞中的作用
賈曉陽 付玉 王瀟楠 王志鋼 郝慧芳
(內蒙古大學生命科學學院微生物實驗室,呼和浩特 010021)
細菌對宿主細胞的黏附和侵襲是引發傳染病的重要步驟。細菌在黏附的過程中,其表面結構或特殊黏附分子和宿主細胞表面受體相互作用激活黏著斑激酶(focal adhesion kinase,FAK),通過FAK/Src-Cortactin-Arp2/3通路和FAK/PI3K-Rac通路調控細胞骨架重排,促進細菌侵入非吞噬細胞。 為了深入探討細菌侵入非吞噬細胞的整個過程及調控機制,就細菌對非吞噬細胞的黏附、侵入以及細胞FAK信號通路在此過程中的調節作用進行綜述。
細菌;黏附;入侵;黏著斑激酶;信號通路
細菌廣泛存在于人類生活的環境當中,人們身體內外也存在著大量的共生菌群。一些細菌會導致人類嚴重的疾病,甚至死亡。黏附是細菌入侵的第一步,細菌已進化出一些表面分子和結構促使其黏附到宿主細胞表面,如菌毛、鞭毛和黏附素等。細菌也會產生一些毒素、侵襲素,還有細菌的III型分泌系統,在這些因子的共同作用下,細菌能夠順利地與宿主細胞結合,并且將入侵的信息傳遞到宿主細胞當中,促使宿主細胞中的激酶發生磷酸化,傳遞細胞信號調節細胞骨架運動,最終促進細菌的入侵。在一些細菌入侵宿主細胞的過程中,細胞黏著斑激酶起重要的調節作用。細菌首先會誘導細胞黏著斑激酶自身的磷酸化,同時激活下游信號通路,調節肌動蛋白聚合,引起細胞骨架重排,最終導致細胞膜的內陷,細菌侵入宿主細胞。為了深入探討細菌侵入非吞噬細胞的整個過程及調控機制,本文就細菌對非吞噬細胞的黏附、侵入以及細胞黏著斑激酶信號通路在此過程中的調節作用進行綜述。
1 細菌對宿主細胞的黏附
黏附是病原菌入侵宿主細胞的第一步,這個過程需要病原菌的黏附因子和宿主細胞表面受體的特異性結合,進而有助于病原菌在宿主細胞表面的定植。細菌已進化出特殊的表面分子和結構促使其能黏附到宿主細胞的表面。細菌有大量的能夠黏附到宿主細胞表面的相關結構,如菌毛、纖毛、莢膜和鞭毛等[1]。菌毛是革蘭氏陰性菌表面的重要結構其主要功能是使細菌黏附到宿主細胞上。在革蘭氏陰性菌中,與黏附相關最典型的是I型菌毛、P菌毛和IV型菌毛。I型菌毛和P菌毛主要通過伴侶—引領的途徑形成[2]。例如,引發腎盂腎炎的大腸桿菌P菌毛,就是通過這一途徑完成菌毛的裝配,其分為三步:(1)菌毛子集通過II型分泌系統,到達細胞膜周質;(2)膜周質中的伴侶分子與菌毛子集結合,并且將菌毛子集運送到細胞外膜上的引領分子;(3)由引領分子對菌毛子集進行裝配[3]。其P菌毛通過PapG蛋白黏附到尿路細胞上糖脂的α-D-吡喃半乳糖-(1-4)糖脂-β-D-吡喃半乳糖苷[4]。IV型菌毛的形成則與I型菌毛和P菌毛的形成過程有所不同。IV型菌毛在細胞質膜內首先形成一個完整的菌毛,然后通過擠壓到達細胞膜外[5]。以上3種類型的菌毛均在黏附過程中起重要的作用。細菌表達的黏附相關的蛋白為黏附素,黏附素不僅分布在菌毛、鞭毛這些結構的末端、或軸上,也分布在細菌細胞表面。例如,磷壁酸是革蘭氏陽性細菌細胞壁組成成分,金黃色葡萄球菌、表皮葡萄球菌和化膿性鏈球菌的黏附過程與磷壁酸有關。金黃色葡萄球菌表面有許多黏附素,如纖連蛋白結合蛋白(fibronectinbinding proteins,FnBPs)、膠原蛋白結合蛋白(collagen adhesin,Cna)、聚集因子(clumping factors,Clfs)等。其中,纖連蛋白結合蛋白與細胞外基質結合,細胞外基質與宿主細胞結合,從而起到黏附的作用[6]。除了細菌細胞表面的黏附素,細菌(主要是革蘭氏陰性菌)自身分泌產生的蛋白到達細胞外膜,黏附到細胞外基質或者宿主細胞表面[7],同時促進細菌的聚集,如引發小腸結腸炎的耶爾森菌中的YadA[8]。
宿主細胞膜表面有些糖類、蛋白等可作為細菌黏附的主要受體。哺乳動物的宿主細胞表面有許多黏附分子,這些黏附受體主要分為四大類:整合素、鈣黏蛋白、免疫蛋白超家族和選擇素[9]。整合素普遍存在于細胞與細胞、細胞與細胞外基質的黏附過程中,它是宿主細胞表面的一個異二聚體,由α和β兩個不同的亞單位,按照不同的組合構成,其中β1整合素家族通常是細胞外基質的受體。有整合素參與的黏附,要與細胞外基質結合共同完成黏附作用(這種黏附方式叫做間接黏附),細胞外基質包括纖連蛋白(fibronectin,Fn)、層黏連蛋白、膠原蛋白。由整合素介導產生結合的細菌有小腸結腸炎耶爾森菌[10]、金黃色葡萄球菌、化膿性鏈球菌等。整合素不僅能介導病原菌與宿主細胞的黏附,并且與細胞信號轉導具有密切的聯系,有研究表明,耶爾森菌外膜蛋白侵襲素促進β1整合素的聚合,激活酪氨酸激酶,引起細胞骨架中肌動蛋白的重排,從而促進病原菌的入侵[11,12]。
病原菌與宿主細胞特異性黏附機制有兩種:一是直接黏附,即細菌自身表面的黏附素可直接與宿主細胞表面的受體結合,如大腸桿菌、李斯特菌[13]、耶爾森菌[14]和淋病奈瑟菌[15]。二是間接黏附,即宿主細胞自身向胞外產生的一些大分子物質,如細胞外基質(extracellular matrix,ECM),這些大分子物質能促進宿主細胞與病原菌的結合,從而達到黏附目的。這些細菌具有與細胞外基質結合的能力,一般是一些革蘭氏陽性菌,如鏈球菌和葡萄球菌[16]。
細菌與宿主細胞發生黏附是一個復雜的過程,不僅僅通過細菌與宿主細胞的一些表面結構來完成。例如,腸致病性大腸桿菌(enteropathogenic Escherichia coli,EPEC)和腸出血性大腸桿菌(enterohemorrhagic Escherichia coli,EHEC)。EPEC/EHEC的束狀菌毛對宿主細胞進行黏附,然后經過III型分泌系統分泌的細菌效應物Tir嵌入到宿主細胞膜上,與細菌表面的外膜蛋白緊密素(Intimin)相互結合,促進細菌與宿主細胞的進一步結合,與此同時,結合的部位會形成一個基座。基座的形成是通過Tir與intimin相互作用,被磷酸化了的Tir誘導蛋白質適配器NCK,使得神經綜合征蛋白(neuronal wiskottaldrich syndrome protein,N- WASP)和Arp2 / 3復合物(actin-related protein 2/3)募集到細菌與宿主細胞結合的部位,導致黏附部位的肌動蛋白聚合從而形成一個基座[17]。肌動蛋白結合蛋白如踝蛋白會被募集到基座,用來穩定其的結構。
2 細菌侵襲進入宿主細胞
細菌入侵宿主細胞的過程中會伴隨著肌動蛋白細胞骨架的重新排列。細菌進入非吞噬細胞的機制有拉鏈式機制和觸發式機制[18]:(1)拉鏈式機制指細菌通過與宿主細胞發生黏附,調節宿主細胞內的一系列信號通路,從而導致細胞骨架發生重排,細胞表面的膜形成偽足,包裹于細菌周圍,細菌內陷于細胞膜[19]。此過程不發生大規模的細胞骨架重排。依靠這種入侵機制,典型的細菌有耶爾森菌、產單核細胞李斯特菌和金黃色葡萄球菌[12,20]。(2)觸發式機制與前者相比,此過程會引起大規模的細胞骨架重排,在細菌與宿主細胞表面結合的過程中,宿主細胞膜上會在臨近入侵細菌的表面形成大型片狀或者偽足樣結構,將細菌卷入到細胞內[18]。這種入侵機制的典型細菌有志賀氏菌和沙門氏菌。細菌入侵細胞伴隨著細胞骨架的重排,細胞骨架發生重排,則是由于細菌與宿主細胞發生黏附的過程中,模擬了宿主信號傳遞最終導致細胞骨架重排。黏著斑激酶(focal adhesion kinase,FAK)、GTP酶通過對相關蛋白的酪氨酸磷酸化,產生一系列對下游分子的級聯反應,最終導致細胞骨架發生重排。例如,GTP酶Rho家族中有Rac、Cdc42、Rho,它們是調控肌動蛋白的主要成員。細胞外基質可誘導Rac活化,調節片狀偽足的形成,Cdc42誘導遷移細胞前沿肌動蛋白絲狀偽足的形成。細菌激活宿主的蛋白激酶不僅僅依賴黏附的過程,有時,病原菌分泌的毒力因子可以直接進入到非吞噬細胞中,與宿主細胞中的蛋白激酶發生作用,從而控制與調節細胞骨架的重排。沙門氏菌產生一些易位效應蛋白,這些效應物可以促進細菌的初始入侵。易位蛋白SipC經過III型分泌系統,進入宿主細胞,促使肌動蛋白聚合并且與F-actin結合;同時SopE激活Rho GTP酶,促進肌動蛋白聚合和膜皺褶形成[21]。志賀氏菌通過III型分泌系統分泌的蛋白OspE作用在整合素的相關酶上,可以促進整合素和黏著斑的表達量上調,并且表達黏著斑的運動能力降低,從而使得黏附細胞外基質的能力提高[22]。
3 細胞黏著斑激酶與細菌入侵
FAK是一類胞質非受體蛋白酪氨酸激酶,可以整合許多細胞外信號,如整合素和機械牽拉等。FAK是相關聯的非受體和非膜蛋白酪氨酸激酶(protein- tyrosine kinase,PTK)[23],其不含有與Src蛋白的SH2(Src homology 2)或SH3(Src homology 3)相互作用的結構域。與FAK有關的PTK已經被分離,包括黏著斑激酶β(cell adhesion kinaseβ,CAKβ),蛋白酪氨酸激酶2(proline-rich tyrosine kinase 2,PYK2),相關黏附斑酪氨酸激酶(related adhesion focal tyrosine kinase,RAFTK),鈣依賴型酪氨酸激酶(calcium-dependent protein-tyrosine kinase,CADTK)和 黏 著 斑 激 酶2(focal adhesion kinase 2,FAK-2)[24-27]。FAK主要包含3個結構域:N末端結構域含有FERM區域,它是整合素的作用位點。當病原菌與宿主細胞發生黏附之后,隨之整合素被激活,整合素β亞基的胞內部分與FAK結合,進而通過Src、磷脂酰肌醇3激酶(phosphtidylinositol 3-kinase,PI3K)等通路將胞外基質的信號導入胞內,引起肌動蛋白的聚合。中間的激酶結構域(PTK),可以激活下游的通路。FAK至少含有6個酪氨酸位點,分別 是Tyr-397、Tyr-407、Tyr-576、Tyr-577、Tyr-861和Tyr-925[28,29]。Tyr-397是FAK主要的磷酸化部位,可以發生自體磷酸化[30]。磷酸化的Tyr-397可以與Src的SH2結構域結合[31],形成復合體,并且激活下游通路。FAK的C末端結構域(FAT)約100個氨基酸,起到黏著斑靶向定位的功能,FAT可以與樁蛋白(Paxillin)[32]和裸蛋白(Talin)[33]結合,從而引起肌動蛋白(Actin)的聚合。在激酶結構域和FAT結構域之間,含有兩個脯氨酸基序,其中靠近C末端結構域的脯氨酸基序能夠與GTPase的SH3結構域結合[34,35]。GRAF是RhoGTPases的激活蛋白,RhoGTPases參與細胞骨架重建并在細胞外信號轉導到細胞內發揮著重要的作用。前述我們知道細菌與宿主細胞相互作用,模擬信號轉導機制,引起局部肌動蛋白絲聚合,促進細菌內化。FAK在這一過程中發揮著關鍵的調節作用。
FAK參與細菌入侵的過程,是由整合素介導的細菌引起的細胞內化。例如,尿路致病性大腸桿菌入侵膀胱細胞(圖1)。尿路致病性大腸桿菌的菌毛是I型菌毛,編碼菌毛的基因是fim基因,I型菌毛首先對宿主細胞進行黏附,整合素α3β1是宿主細胞的黏附受體。菌毛FimH通過識別整合素N端鏈接的甘露糖聚糖殘基,并逐個地與整合素子集α3和β1進行固定[36],與此同時,整合素和F-actin不斷地被募集到細菌的黏附部位。在細菌黏附部位,整合素募集并激活宿主細胞內多種適配器,從而引起細胞信號轉導,產生細胞骨架的重排,進一步促進細菌的入侵。整合素能夠募集和激活FAK、Src和Rho家族的GTP酶[37]。活化的FAK與PI3K形成瞬時復合物,產生一些磷脂酰磷酸鹽(Phosphotidylinositides,PIPs),這些磷脂酰磷酸鹽作為第二信號,可以調節肌動蛋白的動力,一些紐蛋白和α輔肌動蛋白參與到細胞骨架的重排[38-40]。與此同時,Rho家族GTP酶中的Rac1也促進細菌入侵位點的細胞骨架重排。
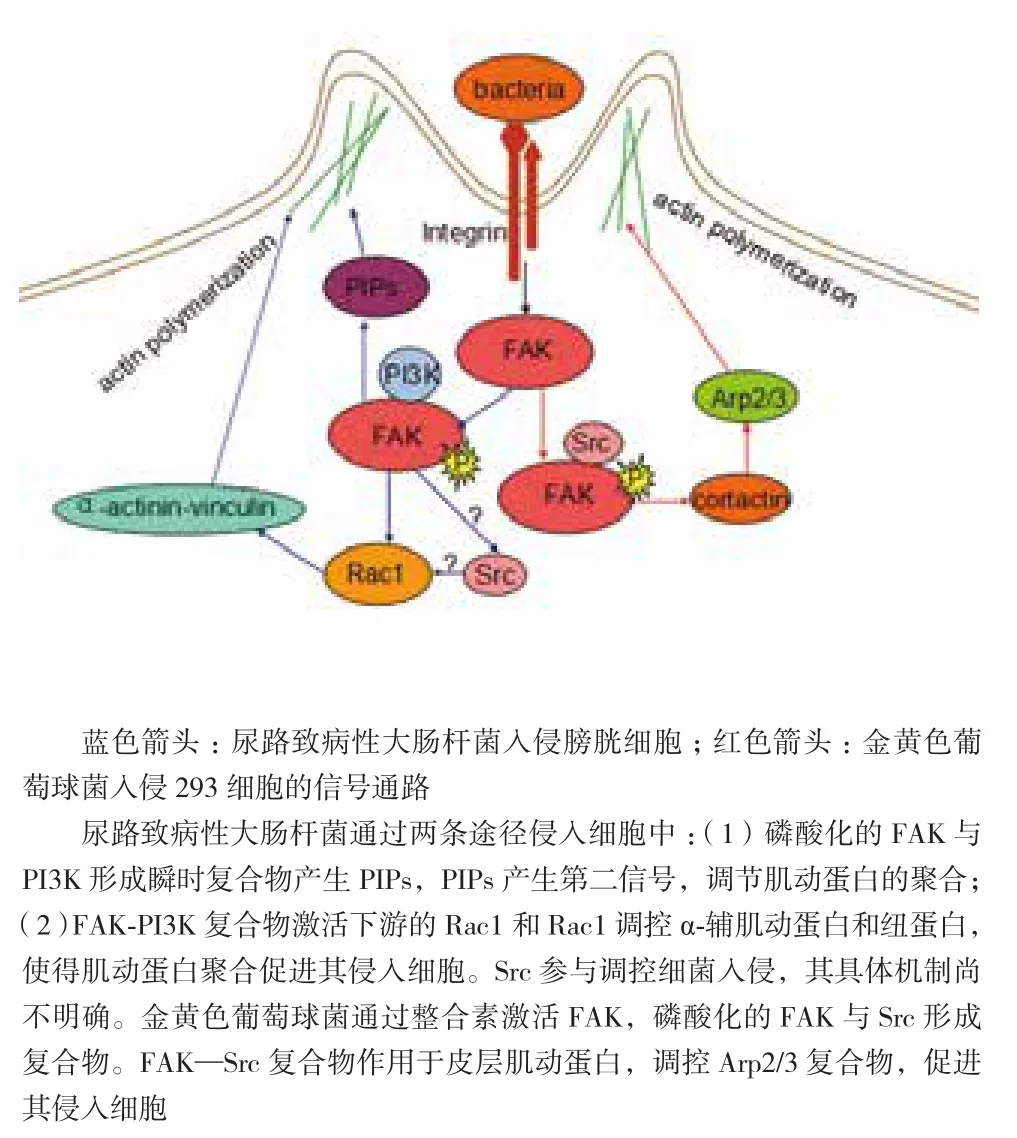
圖1 細菌入侵非吞噬細胞的信號通路
在金黃色葡萄球菌入侵非吞噬細胞的過程(圖1),金黃色葡萄球菌表面表達的纖連蛋白結合蛋白FnBPs,也稱作金黃色葡萄球菌的黏附素,以纖連蛋白Fn為橋梁,將細菌表面的FnBPs和α5β1整合素連接起來[41]。研究表明,缺失FnBPs的菌株突變體會喪失入侵的功能。還有一種熱休克蛋白,可以與整合素形成共受體,通過Fn為橋梁,將細菌與宿主細胞進行連接,產生細胞的內化[42],但是這個作用的機制在真核細胞中還不清楚。在金黃色葡萄球菌入侵293T細胞時,由整合素介導的細菌入侵,不僅可以促進FAK在入侵位點的聚集,還能募集一些相關的蛋白,如張力蛋白、紐蛋白等。有研究表明,缺陷型的FAK[43]和缺陷型的Src[44]細胞可以抵制金黃色葡萄球菌的入侵。這表明,FAK在細菌入侵宿主細胞的過程中發揮重要的作用。被整合素募集并激活的FAK在其397位點上發生自體磷酸化,并與Src結合形成復合物,FAK和Src復合物可以使肌動蛋白的相關蛋白產生磷酸化,從而調控下游的信號通路。皮層肌動蛋白是Src的一個重要底物,在細菌入侵過程中的作用尤為重要。皮層肌動蛋白是肌動蛋白的結合蛋白,它是許多信號轉導的樞紐,并參與Arp2 / 3復合物的聚集(皮層肌動蛋白的NTA(N-terminal acidic motif)結構域能夠與Arp2 / 3復合物結合[45],被激活的Arp2 / 3復合物能使肌動蛋白生成支細絲,并保持70度角[46]),Arp2 / 3復合物調控肌動蛋白的聚合,肌動蛋白能夠控制細胞骨架的重排,從而促進病原菌或病毒的入侵。另一條控制肌動蛋白的通路是,皮層肌動蛋白的SH3結構域結合并激活N-WASP[47,48],N-WASP激活Arp2/3復合物,使得肌動蛋白進行聚合。
已有許多實驗證明,FAK調控細菌入侵細胞的過程。李斯特菌、假結核耶爾森菌、大腸桿菌、引發腦膜炎的奈瑟氏菌等,在這些細菌入侵的過程中,FAK均起到重要的作用。總而言之,細菌入侵細胞會伴隨著胞內黏著斑激酶、Src酶、GTP酶、磷脂酸-3-激酶等產生細胞信號通路轉導,最終引起肌動蛋白聚合,細胞骨架發生重排,促進細菌的入侵。
4 展望
細菌與宿主之間的相互作用機制是目前研究的熱點。細菌改變并利用宿主細胞信號轉導系統,引起肌動蛋白細胞骨架重排,使其侵入到非吞噬細胞中,FAK起著關鍵的調控作用。但是,是否還有其他的蛋白分子也參與調控細菌入侵的過程;這些蛋白分子參與調控細菌入侵的過程是獨立于FAK還是與FAK協同調控?這些問題還需要進一步的探索。細菌侵入非吞噬細胞可以逃避免疫攻擊,再加上抗生素抗藥性的問題給有效治療細菌感染增加了難度,細菌在侵入非吞噬細胞中的調控機制研究為抗菌治療提供了新的思路,除了利用抗生素還可通過抑制其侵入和對宿主細胞功能的影響來治療胞內菌的感染。深入研究細菌在侵入非吞噬細胞中的分子機制為尋找新藥物靶點和探索新抗菌藥奠定理論基礎。
[1]郭曉奎, 童善慶. 細胞微生物學[M]. 上海:第二軍醫大學出版社, 2004:94-96.
[2]Kline KA, F?lker S, Dahlberg S, et al. Bacterial adhesins in hostmicrobe interactions[J]. Cell Host Microbe, 2009, 6:580-592.
[3]Pizarro-Cerdá J, Cossart P. Bacterial adhesion and entry into host cells[J]. Cell, 2006, 124(4):720-727.
[4]Hultgren SJ, Normark S, Abraham SN. Chaperoneassisted assembly and molecular architecture of adhesive pili[J]. Annu Rev Microbiol, 1991, 45:383-415.
[5]Wolfgang M, van Putten JP, Hayes SF, et al. Components and dynamics of fiber formation define a ubiquitous biogenesis pathway for bacterial pili[J]. EMBO J, 2000, 19(23):6408-6418.
[6]Foster TJ, Geoghegan JA, Ganesh VK, et al. Adhesion, invasion and evasion:The many functions of the surface proteins of Staphylococcus aureus[J]. Nat Rev Microbiol, 2014, 1:49-62.
[7]Stones DH, Krachler AM. Fatal attraction:how bacterial adhesins affect host signaling and what we can learn from them[J]. Int J Mol Sci, 2015, 16(2):2626-2640.
[8]B?lin I, Wolf-Watz H. Molecular cloning of the temperature-inducible outer membrane protein 1 of Yersinia pseudotuberculosis[J]. Infect Immun, 1984, 43(1):72-78.
[9]Juliano RL. Signal transduction by cell adhesion receptors and the cytoskeleton:functions of integrins, cadherins, selectins, and immunoglobulin-superfamily members[J]. Annu Rev Pharmacol Toxicol, 2002, 42:283-323.
[10]El Tahir Y, Skurnik M. YadA, the multifaceted Yersinia adhesion[J]. Int J Med Microbiol, 2001, 291(3):209-218.
[11]Isberg RR, Barnes P. Subversion of integrins by enteropathogenic Yersinia[J]. J Cell Sci, 2001, 114(1):21-28.
[12]Boyle EC, Finlay BB. Bacterial pathogenesis:exploiting cellular adherence[J]. Curr Opin Cell Biol, 2003, 15(5):633-639.
[13]Pizarro-Cerdá J, et al. Entry of Listeria monocytogenes in mammalian epithelial cells:an updated view[J]. Cold Spring Harb Perspect in Med, 2012, 2(11). pii:a010009.
[14]Isberg RR, Leong JM. Multiple beta 1 chain integrins are receptors for invasin, a protein that promotes bacterial penetration into mammalian cells[J]. Cell, 1990, 60(5):861-871.
[15]N?gele V, Heesemann J, Schielke S, et al. Neisseria meningitids adhesin NadA targets β1 integrins:functional similarity to Yersinia invasin[J]. J Biol Chem, 2011, 286(23):20536-20546.
[16]Hoffmann C, et al. Integrin-mediated uptake of fibronectin-binding bacteria[J]. Eur J Cell Biol, 2011, 90(11):891-896.
[17]Campellone KG, Leong JM. Nck-independent actin assembly is mediated by two phosphorylated tyrosines within enteropathogenic Escherichia coli Tir[J]. Mol Microbiol, 2005, 56(2):416-432.
[18]張湘燕, 郭曉奎, 劉晶星, 等. 細菌利用宿主肌動蛋白細胞骨架進入非吞噬細胞的機制[J]. 細胞生物學雜志, 2002, 24(3):155-158.
[19]Swanson JA, Baer SC. Phagocytosis by zippers and triggers[J]. Trends Cell Biol. 1995, 5(3):89-93.
[20]Alva-Murillo N, López-Meza JE, Ochoa-Zarzosa A. Nonprofessional Phagocytic Cell Receptors Involved in Staphylococcus aureus Internalization[J]. Biomed Res Int, 2014, 2014:538-546.
[21]Hardt WD, Chen LM, Schuebel KE, et al. S. typhimurium encodes an activator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells[J]. Cell, 1998, 5:815-826.
[22]Kim M, Ogawa M, Mimuro H, et al. Reinforcement of epithelial cell adhesion to basement membrane by a bacterial pathogen as a new infectious stratagem[J]. Virulence, 2010, 1(1):52-55.
[23]Parsons JT, Parsons SJ. Src family protein tyrosine kinases:cooperating with growth factor and adhesion signaling pathways[J]. Curr Opin Cell Biol, 1997, 9(2):187-192.
[24]Sasaki H, Nagura K, Ishino M, et al. Cloning and kinase characterization of cell adhesion kinase beta, a novel proteintyrosine kinase of the focal adhesion subfamily[J]. J Biol Chem, 1995, 270(36):21206-21219.
[25]Lev S, Moreno H, Martinez R, et al. Protein tyrosine kinase PYK2 involved in Ca2+-induced regulation of ion channel and MAP kinase functions[J]. Nature, 1995, 376(6543):737-745.
[26]Avraham S, London R, Fu Y, et al:Identification and characterization of a novel related adhesion focal tyrosine kinase(RAFTK)from megakaryocytes and brain[J]. J Biol Chem, 1995, 270:27742-27751.
[27]Yu H, Li X, Marchetto GS, et al. Activation of a novel calciumdependent protein-tyrosine kinase. Correlation with c-Jun N-terminal kinase but not mitogen-activated protein kinase activation[J]. J Biol Chem, 1996, 271(47):29993-29998.
[28]Calalb MB, et al. Tyrosine phosphorylation of focal adhesion kinase at sites i n the catalytic domain regulates kinase activity:a role for Src family kinases[J]. Mol Cell Biol, 1995, 2:954-963.
[29]Schlaepfer DD, Hunter T. Evidence for in vivo phosphorylation of the Grb2 SH2-domain binding site on focal adhesion kinase by Srcfamily protein-tyrosine kinases[J]. Mol Cell Biol, 1996, 16(10):5623-5633.
[30]Burgaya F, Toutant M, Studler JM, et al. Alternatively spliced focal adhesion kinase in rat brain with increased autophosphorylation activity[J]. J Biol Chem, 1997, 272(45):28720-28725.
[31]Schwartz MA, et al. Integrins:emerging paradigms of signal transduction[J]. Annu Rev Cell Dev Biol, 1995, 11:549-599.
[32]Brown MC, et al. Identification of LIM3 as the principal determinant of paxillin focal adhesion localization and characterization of a novel motif on paxillin directing vinculin and focal adhesion kinase binding[J]. J Cell Biol, 1996, 135(4):1109-1123.
[33]Chen HC, Appeddu PA, Parsons JT, et al. Interaction of focal adhesion kinase with cytoskeletal protein talin[J]. J Biol Chem, 1995, 270(28):16995-16999.
[34]Hildebrand JD, Taylor JM, Parsons JT. An SH3 domain-containing GTPase-activating protein for Rho and Cdc42 associates with focal adhesion kinase[J]. Mol Cell Biol, 1996, 16(6):3169-3178.
[35]Ohba T, Ishino M, Aoto H, et al. Interaction of two proline-rich sequences of cell adhesion kinase beta with SH3 domains of p130Cas-related proteins and a GTPase-activating protein[J]. Biochem J, 1998, 330(Pt 3):1249-1254.
[36]Eto DS, Jones TA, Sundsbak JL, et al. Integrin-mediated host cell invasion by type 1-piliated uropathogenic Escherichia coli[J]. PLoS Pathog, 2007, 3(7):e100.
[37]Martinez JJ, Hultgren SJ. Requirement of Rho-family GTPases in the invasion of Type 1-piliated uropathogenic Escherichia coli[J]. Cell Microbiol, 2002, 4(1):19-28.
[38]Shoelson SE, Sivaraja M, Williams KP, et al. Specific phosphopeptide binding regulates a conformational change in the PI 3-kinase SH2 domain associated with enzyme activation[J]. EMBO J, 1993, 12(2):795-802.
[39]Yin HL, Janmey PA. Phosphoinositide regulation of the actin cytoskeleton[J]. Annu Rev Physiol, 2003, 65:761-789.
[40]Hartwig JH, Bokoch GM, Carpenter CL, et al. Thrombin receptor ligation and activated Rac uncap actin filament barbed ends through phosphoinositide synthesis in permeabilized human platelets[J]. Cell, 1995, 82(4):643-653.
[41]Joh D, Wann ER, Kreikemeyer B, et al. Role of fibronectin-binding MSCRAMMs in bacterial adherence and entry into mammalian cells[J]. Matrix Biol, 1999, 18(3):211-223.
[42]Dziewanowska K, Carson AR, Patti JM, et al. Staphylococcal fibronectin binding protein interacts with heat shock protein 60 and integrins:role in internalization by epithelial cells[J]. Infect Immun, 2000, 68(11):6321-6328.
[43]Agerer F, Lux S, Michel A, et al. Cellular invasion by Staphylococcus aureus reveals a functional link between focal adhesion kinase and cortactin in integrin-mediated internalization[J]. J Cell Sci 2005, 118:2189-2200.
[44]Agerer F, Lux S, Michel A, et al. Cellular invasion by Staphylococcus aureus reveals a functional link between focal adhesion kinase and cortactin in integrin-mediated internalisation[J]. J Cell Sci, 2005, 118(10):2189-2200.
[45]Martinez-Quiles N, et al. Erk/Src phospho-rylation of cortactin acts as a switch on-switch off mechanism that controls its ability to activate N-WASP[J]. Mol Cell Biol, 2004, 12:5269-5280.
[46]Pollard TD, Cooper JA. Actin, a central player in cell shape and movement[J]. Science, 2009, 326(5957):1208-1212.
[47]Kurisu S, Takenawa T. The WASP and WAVE family proteins[J]. Genome Biol, 2009, 10(6):226.
[48]Martinez-Quiles N, Ho HY, Kirschner MW, et al. Erk/Src phosphorylation of cortactin acts as a switch on switch off mechanism that controls its ability to activate N-WASP[J]. Mol Cell Biol, 2004, 24(12):5269-5280.
(責任編輯 狄艷紅)
The Role of Focal Adhesion Kinase Signaling Pathway in the Bacterial Invasion of Non-phagocytic Cells
JIA Xiao-yang FU Yu WANG Xiao-nan WANG Zhi-gang HAO Hui-fang
(Microbiology Lab of College of Life Science in Inner Mongolia University,Hohhot 010021)
The adhesion and invasion into eukaryotic cells are major steps in bacterial pathogenesis. In order to further study the entire process and molecular mechanisms of bacteria invading non-phagocytic cells,we reviewed the bacterial adhesion and invasion of nonphagocytic cells and the regulatory mechanisms of focal adhesion kinase(FAK)in this process. During the adhesion process,the interaction of surface adhesive molecules and special structures of bacteria with receptors of host cells activates the FAK to regulate the cytoskeletal rearrangement through FAK/Src-Cortactin-Arp2/3 and FAK/PI3K-Rac pathways,which promotes the bacterial invasion of non-phagocytic cells.
bacteria;adhesion;invasion;focal adhesion kinase;signaling pathway
10.13560/j.cnki.biotech.bull.1985.2016.07.004
2015-10-08
內蒙古自治區高等學校科學研究項目(NJZZ002),內蒙古大學高層次人才引進科研項目(135130)
賈曉陽,女,碩士研究生,研究方向:微生物;E-mail:740405724@qq.com
郝慧芳,女,博士,研究方向:生物病理學;E-mail:haohf@life.imu.edu.cn
