基于COⅠ基因序列對我國部分巴貝斯蟲分類的研究
2016-07-14 00:42:49茍惠天薛慧文孫曉林羅建勛
畜牧獸醫學報 2016年6期
茍惠天,薛慧文,殷 宏,孫曉林*,羅建勛*
(1甘肅農業大學動物醫學院,蘭州 730070;2中國農業科學院蘭州獸醫研究所,蘭州 730046)
?
基于COⅠ基因序列對我國部分巴貝斯蟲分類的研究
茍惠天1,薛慧文1,殷宏2,孫曉林1*,羅建勛2*
(1甘肅農業大學動物醫學院,蘭州 730070;2中國農業科學院蘭州獸醫研究所,蘭州 730046)
為了確定巴貝斯蟲中國分離株的分類地位,對我國報道的7種巴貝斯蟲11個地方分離株的COⅠ基因序列進行測定;并與GenBank中其他巴貝斯蟲COⅠ和相應18S rRNA基因序列分別構建系統發生樹,比較基于不同基因的分類結果。結果顯示:11株巴貝斯蟲的COⅠ基因大小在935~999 bp,與18S rRNA基因比較,COⅠ基因序列含有較多的變異位點及簡約信息位點;基于兩個基因的系統發育樹,對于牛巴貝斯蟲的分類結果基本一致。但也存在如下差異:18S rRNA無法將泰勒蟲與巴貝斯蟲進行區分,而COⅠ基因可明顯區分;羊巴貝斯蟲新疆未定種的分類地位在基于COⅠ基因的分類中更具合理性。來源于COⅠ和18S rRNA的信息都說明我國莫氏巴貝斯蟲的不同地方株間可能存在亞種關系。該研究為巴貝斯蟲的分子分類提供了候選基因。
巴貝斯蟲;分類;系統發育樹;COⅠ基因
巴貝斯蟲(Babesiaspp.)屬于頂復門、梨形蟲綱、巴貝斯蟲科、巴貝斯蟲屬,是一類能引起人和動物巴貝斯蟲病的蜱傳性血液原蟲。該病呈全球分布,給公共健康和畜牧業發展帶來重大影響[1]。在中國引起牛巴貝斯蟲病的病原有雙芽巴貝斯蟲(Babesiabigemina)、牛巴貝斯蟲(B.bovis)、卵形巴貝斯蟲(B.ovata)、大巴貝斯蟲(B.major)、東方巴貝斯蟲(B.orientalis)和巴貝斯蟲未定種喀什分離株(BabesiaU sp.Kashi)。已證實微小扇頭蜱、長角血蜱、刻點血蜱、鐮形扇頭蜱、小亞璃眼蜱及其他一些硬蜱可以傳播這些巴貝斯蟲[2-3]。引起羊巴貝斯蟲病的病原有綿羊巴貝斯蟲(B.ovis)、莫氏巴貝斯蟲(B.motasi)和巴貝斯蟲未定種新疆分離株(Babesiasp.Xinjiang);已知青海血蜱、長角血蜱和小亞璃眼蜱是以上三種巴貝斯蟲的傳播媒介[4-5]。
近年來,18S rRNA作為分子標記在巴貝斯蟲的研究中得到了廣泛應用[6]。但是由于物種多樣性以及單個標記基因很難真實反映物種間的進化關系。所以一些新的標記基因也被用于此類研究,諸如轉錄間隔區(ITS)及主要表面蛋白基因(MPSP)的應用[7-8]。細胞色素C氧化酶基因Ⅰ(COⅠ)作為線粒體基因組的重要組成部分,較之于核糖體基因,更能真實地反映物種的遺傳信息,因此近年來被廣泛地用于物種的系統進化分析及生物條形碼研究中[9-10]。但迄今為止,有關以COⅠ作為標記基因對巴貝斯蟲的進化分析的研究,在國內外還未見有報道。筆者試圖通過比較基于兩個分類基因COⅠ和18S rRNA在巴貝斯蟲中的進化關系,進一步了解巴貝斯蟲不同蟲株之間的遺傳進化關系[11]。
1 材料與方法
1.1蟲株和實驗動物
本試驗涉及的11株巴貝斯蟲均分離自中國。包括六株牛巴貝斯蟲和五株羊巴貝斯蟲。以上蟲株在實驗室液氮保存,詳細資料見表1。
6~12月齡的黃牛和綿羊購自無巴貝斯蟲病報道的地區。在試驗前30 d所有的實驗動物摘除脾。在試驗前10 d,所有的動物取耳尖血制備涂片,再次檢查是否有巴貝斯蟲感染。只有檢查結果為陰性的動物,方可用于后續試驗。
1.2DNA提取、目的基因擴增及測序
對6只黃牛和5只綿羊分別感染含有不同巴貝斯蟲分離株的蟲血各10 mL。當染蟲率大于5%時,靜脈采血,分離紅細胞。使用DNA提取試劑盒(QIAGEN,德國)進行巴貝斯蟲基因組的提取,方法按說明書操作,分光光度計測算DNA的濃度。以未感染梨形蟲的動物血液作為陰性對照。
利用GenBank已公布的巴貝斯蟲COⅠ基因的序列,設計兩對引物。BaF1(ORF第91位):5′-ATAGGATTCTATATGAGTAT-3′,BaR1(ORF第1 318位):5′-ATAATCAGGTATTCTCCTTGG-3′;BaF2(ORF第190位):5′-TCTCTTCATGGTTTAATTATGATAT-3′,BaR2(ORF第1 315位):5′-TA-GCTCCAATTGATAAAACAAAGTG-3′。PCR擴增按照50 μL體系進行,其中基因組DNA 100 ng,上下游引物各1 pmol·L-1,dNTP 160 μmol·L-1,TaqDNA聚合酶1 U,MgCl21.75 mmol·L-1,最后用滅菌水補足50 μL,以上試劑均購自大連寶生物公司。PCR反應條件:94 ℃預變性5 min;94 ℃變性1 min,63 ℃或58 ℃退火1 min,72 ℃延伸3 min,35個循環;72 ℃終延伸8 min,最后4 ℃保存。取5 μL PCR產物再進行電泳,觀察結果。
切膠純化PCR產物,然后將目的片段克隆到pGEM-T easy載體(Promega,美國)。為確保結果的準確性,每株巴貝斯蟲送5個鑒定為陽性的克隆到測序公司,用ABI 3730進行序列測定。另外,對測序巴貝斯蟲的相應18S rRNA基因序列,分別從GenBank中進行下載。
1.3序列比對和進化分析
使用Lasergene軟件包中的SeqMan對每一條測序結果進行拼接,并將拼接結果在NCBI中作BLAST比對,檢測是否混雜有其他DNA。同時以兩株泰勒蟲(Theileriaparva和Theileriasergenti)的COⅠ基因序列作為外群。使用MEGA4中的多序列比對方法,分別對COⅠ和18S rRNA兩組序列進行比對,使用K2-P方法計算遺傳距離[12]。用最大簡約法(MP)和貝葉斯法(BI)分別針對COⅠ和18S rRNA序列構建系統發育樹。在最大簡約法中,使用TBR啟發式搜索的方法對數據進行分析,空位被視為缺失,顛換和轉換具有相等的權重,自展分析次數為1 000[13]。在貝葉斯法中,分析采用GTR+I+Γ模型,四條鏈分別進行計算,COⅠ和18S rRNA各運算2.0×105代后數據得到收斂[14]。最后將兩種方法產生的系統發育樹用Tree View軟件顯示,并進一步編輯。
2 結 果
2.1COⅠ基因的擴增
從11株巴貝斯蟲基因組中均擴增到COⅠ基因序列,大小為935~999 bp,經Blast N比對無誤后登錄GenBank,序列號見表1。
表1本研究中所用到COⅠ 和18S rRNA的序列信息
Table 1Origins of the piroplasma and GenBank accession numbers for theCOⅠ and 18S rRNA gene used in this study
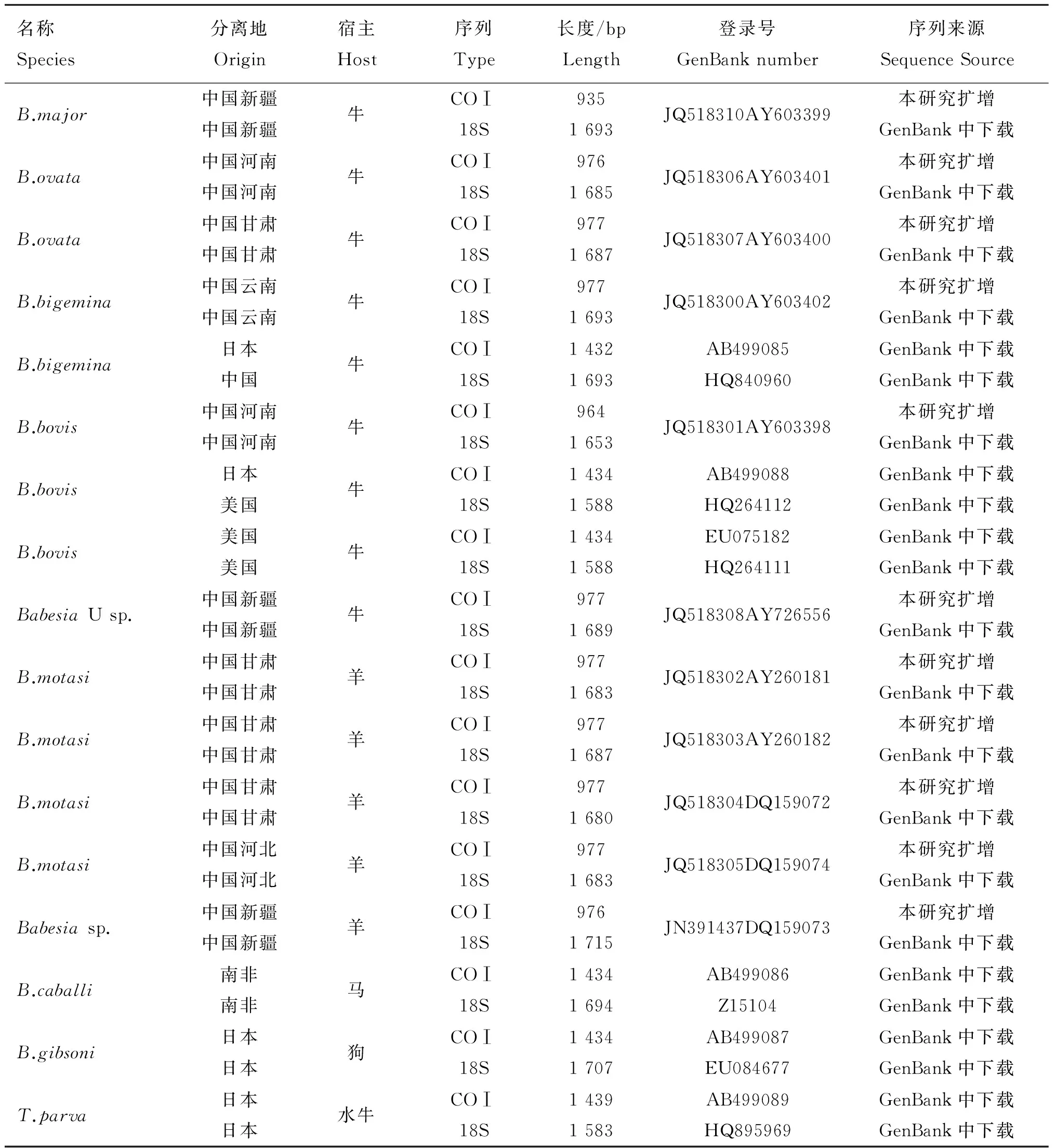
名稱Species分離地Origin宿主Host序列Type長度/bpLength登錄號GenBanknumber序列來源SequenceSourceB.major中國新疆中國新疆牛COⅠ18S9351693JQ518310AY603399本研究擴增GenBank中下載B.ovata中國河南中國河南牛COⅠ18S9761685JQ518306AY603401本研究擴增GenBank中下載B.ovata中國甘肅中國甘肅牛COⅠ18S9771687JQ518307AY603400本研究擴增GenBank中下載B.bigemina中國云南中國云南牛COⅠ18S9771693JQ518300AY603402本研究擴增GenBank中下載B.bigemina日本中國牛COⅠ18S14321693AB499085HQ840960GenBank中下載GenBank中下載B.bovis中國河南中國河南牛COⅠ18S9641653JQ518301AY603398本研究擴增GenBank中下載B.bovis日本美國牛COⅠ18S14341588AB499088HQ264112GenBank中下載GenBank中下載B.bovis美國美國牛COⅠ18S14341588EU075182HQ264111GenBank中下載GenBank中下載BabesiaUsp.中國新疆中國新疆牛COⅠ18S9771689JQ518308AY726556本研究擴增GenBank中下載B.motasi中國甘肅中國甘肅羊COⅠ18S9771683JQ518302AY260181本研究擴增GenBank中下載B.motasi中國甘肅中國甘肅羊COⅠ18S9771687JQ518303AY260182本研究擴增GenBank中下載B.motasi中國甘肅中國甘肅羊COⅠ18S9771680JQ518304DQ159072本研究擴增GenBank中下載B.motasi中國河北中國河北羊COⅠ18S9771683JQ518305DQ159074本研究擴增GenBank中下載Babesiasp.中國新疆中國新疆羊COⅠ18S9761715JN391437DQ159073本研究擴增GenBank中下載B.caballi南非南非馬COⅠ18S14341694AB499086Z15104GenBank中下載GenBank中下載B.gibsoni日本日本狗COⅠ18S14341707AB499087EU084677GenBank中下載GenBank中下載T.parva日本日本水牛COⅠ18S14391583AB499089HQ895969GenBank中下載GenBank中下載
2.2COⅠ基因和18S rRNA基因的序列比對
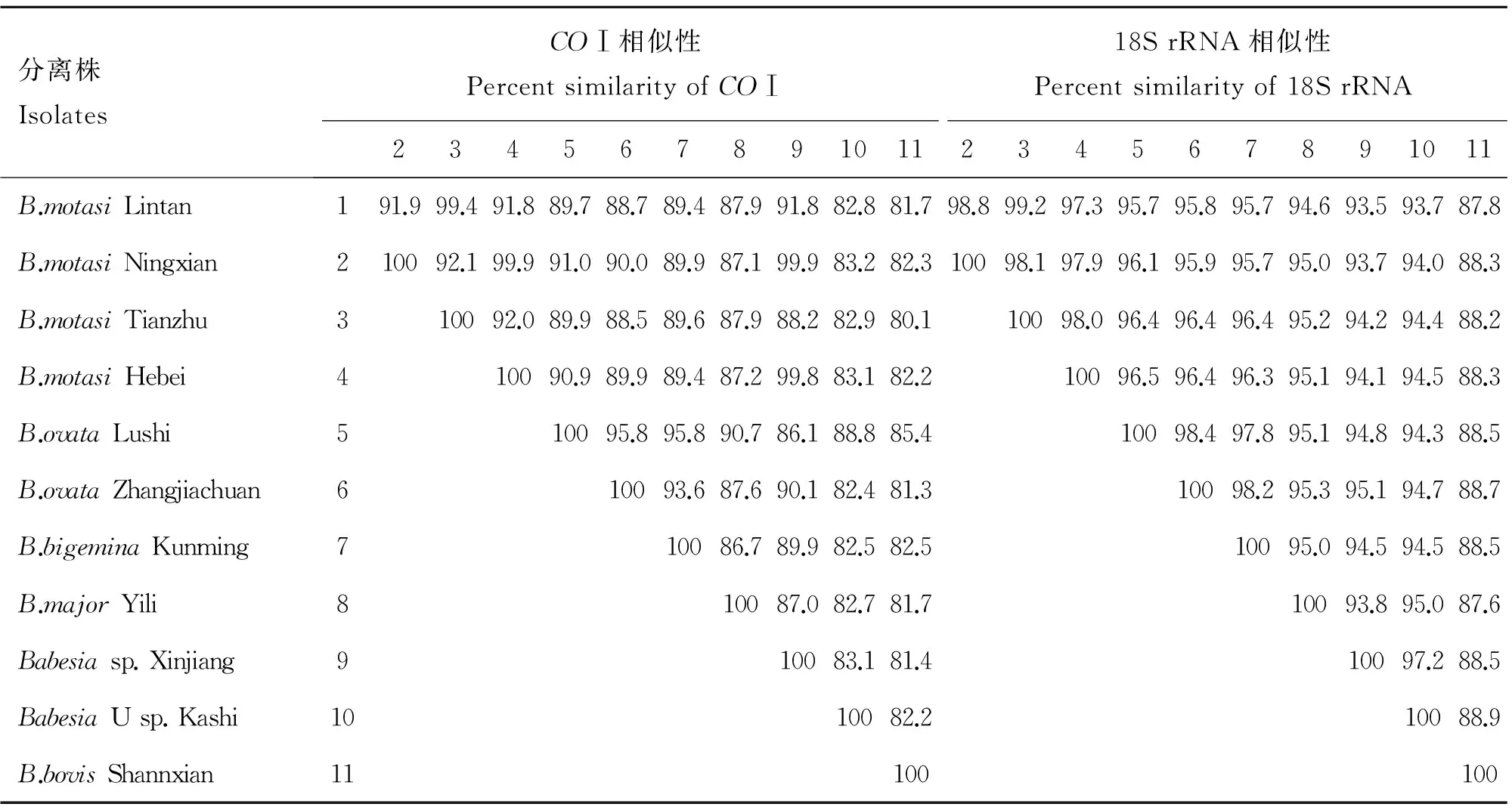
從GenBank中下載到18S rRNA基因的大小為1 583~1 715 bp。所有序列經Claustal W比對后,得出18S rRNA基因含有1 563個信息位點,其中變異位點308個,248個為簡約信息位點;COⅠ基因中含有923個信息位點,其中變異位點354個,簡約信息位點281個。基于兩個基因不同種之間,同一種不同分離株之間的序列相似性見表2。
2.3系統進化分析
為了確定巴貝斯蟲的系統發生關系,分別對COⅠ和18S rRNA基因序列用MP法和BI法構建了系統發育樹。由于每一個基因其MP樹和BI樹的拓撲結構幾乎一致,所以用合一樹來表示。圖1中A、B分別代表COⅠ和18S rRNA的合一樹。通過比較,基于兩個基因的系統發育樹,都能真實反應牛巴貝斯蟲間的分類地位。但也存在如下差異:發現18S無法將泰勒蟲與巴貝斯蟲進行很好區分,而COⅠ基因可明顯區分;羊巴貝斯蟲新疆未定種的分類地位在兩個發育樹中存在較大差異。
表2巴貝斯蟲中國分離株COⅠ和18S rRNA基因序列的相似性比對
Table 2Sequence similarity ofCOⅠand 18S rRNA ofBabesiaspp.from China isolates
%
3 討 論
國內外有關巴貝斯蟲的分子分類學研究主要依靠18S rRNA基因。但近年來越來越多的研究顯示,18S rRNA作為分類基因在科或更高階元的研究時,顯示出一定優勢;在本研究中,18S rRNA變異位點主要集中在少數幾個區域,如:第157—216位,第568—625位,第701—733位,第1 233—1 259位,其余部位則為高度保守。由于其序列的高度保守性,在區分親緣關系很近的物種(屬以下)時,其結果不甚理想。線粒體基因組,特別是COⅠ基因,因其獨立的進化機制,在很大程度上保證了物種遺傳信息的真實性。另外,具有適度的序列變異性,使其在任何分類階元中都可使用。因此,國內外越來越多的學者將其作為新的分類基因。國際DNA條形碼聯盟更是推薦COⅠ為首選基因,進行物種的分子分類。與18S rRNA比較,COⅠ基因的變異位點比例更高,但整段序列并沒有明顯的保守區和變異區,而是變異位點和保守位點交替排列。因此,基于COⅠ基因的巴貝斯蟲分類,能夠更清楚地反映出巴貝斯蟲與泰勒蟲的分類地位,不同宿主巴貝斯蟲之間、同一種巴貝斯蟲不同分離株間的關系[15]。
有關羊的巴貝斯蟲在之前的研究中主要有兩種:莫氏巴貝斯蟲(B.motasi)和羊巴貝斯蟲(B.ovis),其傳播媒介分別是刻點血蜱和囊性扇頭蜱,隨后的研究發現粗糙巴貝斯蟲(B.crassa)也可感染羊。有關莫氏巴貝斯蟲的分類,有學者認為其存在亞種。Uilenberg提出至少可以將歐洲的巴貝斯蟲分為高致病性和低致病性兩個類群[16-17]。筆者實驗室在過去的研究中,從不同地區綿羊和山羊的血液樣品中曾分離到幾株大型巴貝斯蟲[4,18]。這些巴貝斯蟲后來被命名為莫氏巴貝斯蟲臨潭株(B.motasiLintan)、莫氏巴貝斯蟲寧縣株(B.motasiNingxian)、莫氏巴貝斯蟲天祝株(B.motasiTianzhu)和莫氏巴貝斯蟲河北株(B.motasiHebei)。通過COⅠ系統發育樹,可以看到臨潭株和天祝株落在一個分枝中,河北株和寧縣株在另一分枝中。該結果與18S rRNA的進化樹不太一致,但是與ITS分析的結果相一致。寧縣株是從甘肅省東部地區的綿羊血液之中分離得到的,對綿羊和山羊具有高致病性,其形狀與歐洲的莫氏巴貝斯蟲非常相似,長角血蜱是已知該巴貝斯蟲的傳播媒介。臨潭株可被青海血蜱和長角血蜱傳播,并且具有低致病性。河北株和天祝株的生物學特性尚不十分清楚,但是它們都分離自血蜱屬分布的地區[17]。此外,以臨潭株體外培養物經純化后作為抗原,進行的ELISA試驗結果顯示,與寧縣株和河北株的陽性血清不發生交叉反應,但卻可以與天祝株的陽性血清具有很強的交叉反應[19]。以上結果說明我國莫氏巴貝斯蟲的不同地方株間很可能存在亞種關系。
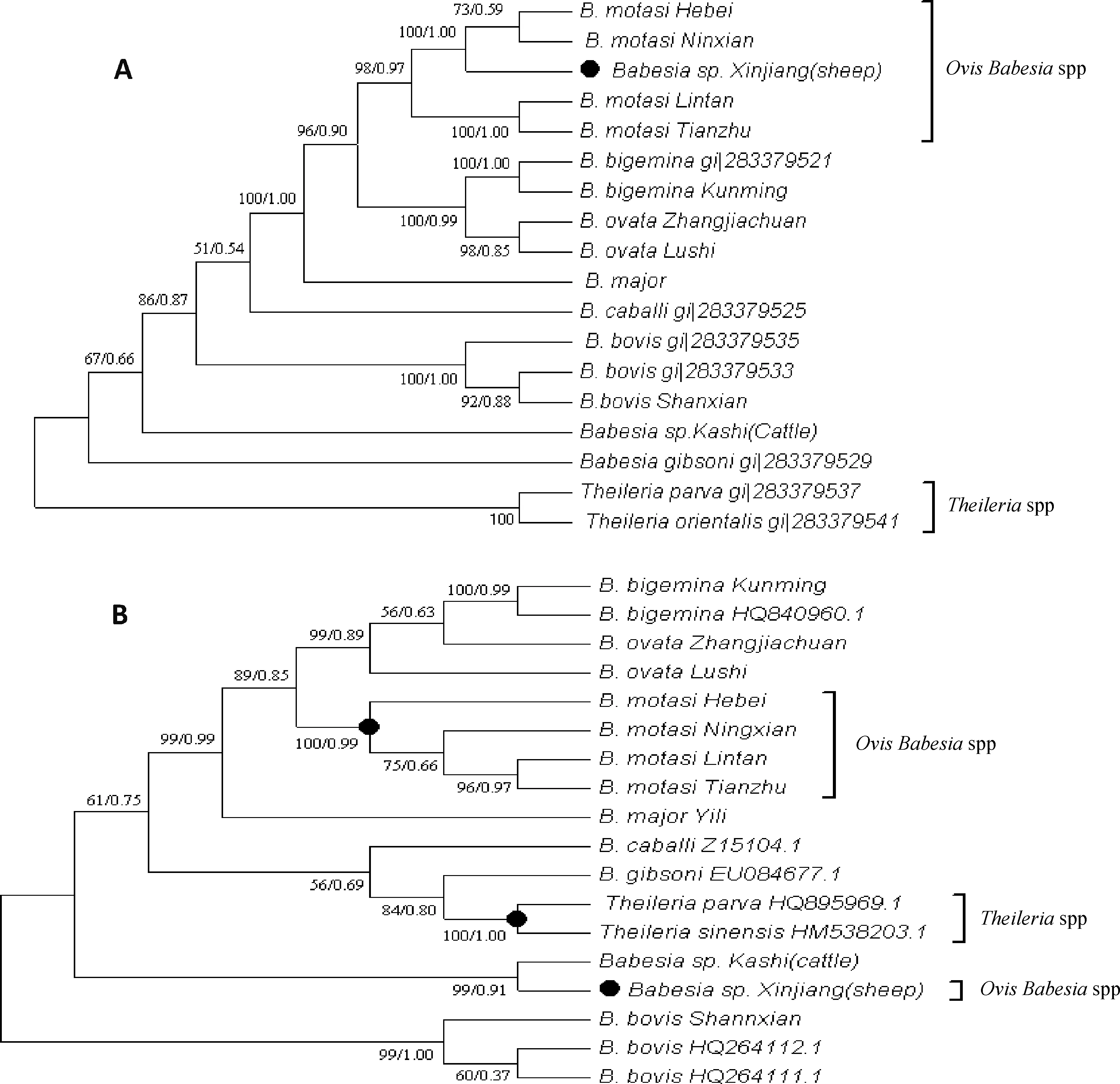
A.COⅠ基因序列;B.18S rRNA基因序列;系統發育樹的構建采用MP和BI兩種計算方法,節點數字分別表示自展值和后驗概率;兩株泰勒蟲作為外群These trees were calculated using the BI and MP methods.Trees were constructed with COⅠ sequences (A) and 18S rRNA sequences (B).The bootstrap values supporting and posterior probability each node are shown.Two Theileria isolates were usedas outgroup圖1 基于COⅠ和18S rRNA基因的巴貝斯蟲系統發育樹Fig.1 Phylogenetic trees of Babesia isolates based on their COⅠand 18S rRNA genes
巴貝斯蟲新疆未定種(Babesiasp.Xinjiang)是筆者實驗室于2001年從新疆采集的血紅扇頭蜱和小亞璃眼蜱,感染綿羊后,經由羊血中分離得到的一種巴貝斯蟲。綜合形態學觀察,傳播試驗等生物學特性,確定該巴貝斯蟲是獨立于莫氏巴貝斯蟲、綿羊巴貝斯蟲、粗糙巴貝斯蟲之外的一個新種[17]。本研究中,基于18S rRNA的系統發育樹中,Babesiasp.Xinjiang與其他四株羊巴貝斯蟲的位置較遠,反而與一株同樣分離自新疆,感染牛的巴貝斯蟲(Babesiasp.Kashi)歸為一支。該結果的出現可能與構建系統發育樹所用的序列相關。而在基于COⅠ的系統發育樹中,該未定種與莫氏巴貝斯蟲歸為一支,并位于致病性不同的四株莫氏巴貝斯蟲之間,顯示與莫氏巴貝斯蟲存在很近的遺傳關系。綜合以上18S rRNA和COⅠ的結果,都未能真實反應該巴貝斯蟲未定種的分類地位。因此,對于該未定種的命名還需更多生物學數據進行佐證[11]。
總之,與之前基于18S rRNA和其他基因的研究結果比較,COⅠ基因的應用,可以幫助更好地解釋巴貝斯蟲各種之間的進化關系。但由于COⅠ基因在巴貝斯蟲上研究非常少,以致基因數據庫中能用到的數據量還很有限。因此,通過本研究,希望能引起廣大研究者的興趣,在以后的分類研究中引入COⅠ基因,推動巴貝斯蟲甚至梨形蟲的分類研究。
[1]TELFORD S R 3rd,SPIELMAN A.Reservoir competence of white-footed mice forBabesiamicroti[J].JMedEntomol,1993,30(1):223-227.
[2]LUO J,YIN H,GUAN G,et al.A comparison of small-subunit ribosomal RNA gene sequence of bovineBabesiaspecies transmitted byHaemaphysalisspp. in China[J].ParasitolRes,2005,95(2):145-149.
[3]YIN H,LU W,LUO J,et al.Experiments on the transmission ofBabesiamajorandBabesiabigeminabyHaemaphysalispunctata[J].VetParasitol,1996,67(1-2):89-98.
[4]GUAN G Q,YIN H,LUO J X,et al.Transmission ofBabesiasp to sheep with field-collectedHaemaphysalisqinghaiensis[J].ParasitolRes,2002,88(13 Suppl 1):22-24.
[5]NIU Q,LUO J,GUAN G,et al.Differentiation of two ovineBabesiabased on the ribosomal DNA internal transcribed spacer (ITS) sequences[J].ExpParasitol,2009,121(1):64-68.
[6]AHMED J S,LUO J,SCHNITTGER L,et al.Phylogenetic position of small-ruminant infecting piroplasms[J].AnnNYAcadSci,2006,1081:498-504.
[7]LIU J,YIN H,LIU G,et al.Discrimination ofBabesiamajorandBabesiaovatabased on ITS1-5.8S-ITS2 region sequences of rRNA gene[J].ParasitolRes,2008,102(4):709-713.
[8]LIU A,GUAN G,LIU Z,et al.Detecting and differentiatingTheileriasergentiandTheileriasinensisin cattle and yaks by PCR based on major piroplasm surface protein (MPSP)[J].ExpParasitol,2010,126(4):476-481.
[9]MATZEN DA SILVA J,CREER S,DOS SANTOS A,et al.Systematic and evolutionary insights derived from mtDNA COI barcode diversity in the decapoda (Crustacea:Malacostraca)[J].PLoSOne,2011,6(5):e19449.
[10]DERYCKE S,VANAVERBEKE J,RIGAUX A,et al.Exploring the use of cytochrome oxidase c subunit 1 (COI) for DNA barcoding of free-living marine nematodes[J].PLoSOne,2010,5(10):e13716.
[11]ARMSTRONG P M,KATAVOLOS P,CAPORALE D A,et al.Diversity ofBabesiainfecting deer ticks (Ixodesdammini)[J].AmJTropMedHyg,1998,58(6):739-742.
[12]TAMURA K,DUDLEY J,NEI M,et al.MEGA4:molecular evolutionary genetics analysis (MEGA) software version 4.0[J].MolBiolEvol,2007,24(8):1596-1599.
[13]SWOFFORD D.PAUP 4.0 b10:Phylogenetic analysis using parsimony[M].Sunderland,MA,USA:Sinauer Associates,2002.
[14]RONQUIST F,HUELSENBECK J P.MrBayes 3:Bayesian phylogenetic inference under mixed models[J].Bioinformatics,2003,19(12):1572-1574.
[15]MEDINA M,COLLINS A G,SILBERMAN J D,et al.Evaluating hypotheses of basal animal phylogeny using complete sequences of large and small subunit rRNA[J].ProcNatlAcadSciUSA,2001,98(17):9707-9712.
[16]UILENBERG G.Babesia—a historical overview[J].VetParasitol,2006,138(1-2):3-10.
[17]BAI Q,LIU G,LIU D,et al.Isolation and preliminary characterization of a largeBabesiasp.from sheep and goats in the eastern part of Gansu Province,China[J].ParasitolRes,2002,88(13 Suppl 1):S16-S21.
[18]LIU A H,YIN H,GUAN G Q,et al.At least two genetically distinct largeBabesiaspecies infective to sheep and goats in China[J].VetParasitol,2007,147(3-4):246-251.
[19]GUAN G,MOREAU E,LIU J,et al.Babesiasp.BQ1 (Lintan):Molecular evidence of experimental transmission to sheep byHaemaphysalisqinghaiensisandHaemaphysalislongicornis[J].ParasitolInt,2010,59(2):265-267.
(編輯白永平)
Toxonomic Study of SomeBabesiaspp.Based onCOⅠGene in China
GOU Hui-tian1,XUE Hui-wen1,YIN Hong2,SUN Xiao-lin1*,LUO Jian-xun2*
(1.CollegeofVeterinaryMedicine,GansuAgriculturalUniversity,Lanzhou730070,China;2.LanzhouVeterinaryResearchInstituteofChineseAcademyofAgriculturalScience,Lanzhou730046,China)
In order to clarify the taxonomic status ofBabesiaspp,theCOⅠgene were sequenced from 11 isolates of 7 species reported in China;The phylogenetic tree were constructed usingCOⅠand 18S rRNA sequences downloaded from GenBank and this study,and also the two phylogenetic trees were compared.As result shown,the length ofCOⅠgene was 935-999 bp,and theCOⅠgene had more variation and informative sites than 18S rRNA.The taxonomic status of bovisBabesiaspp.based onCOⅠand 18S rRNA shared high similarity.However,there were also some differences from two phylogenetic trees.Such as,theCOⅠgene can differentiateBabesiaspp andTheileriaspp clearly.To 18S rRNA,it was more difficult.The taxonomic status based onCOⅠgene ofBabesiasp.from Xinjiang was more reasonable.The information of two phylogenetic tree suggested that theBabesiamotasiisolated from different region may represent different subspecies.This study supported the candidate gene of molecular taxonomy ofBabesiaspp.
Babesiaspp.;toxonomy;phylogenetic tree;COⅠgene
10.11843/j.issn.0366-6964.2016.06.028
2015-11-16
國家自然科學基金(31560700);甘肅省自然科學基金(145RJZA172);蘭州市科技創新人才計劃(2014-2-11);甘肅農業大學盛彤笙創新基金(GSAU-STS-1422)
茍惠天(1980-),甘肅臨洮人,講師,博士,主要從事獸醫公共衛生研究工作,E-mail:gouht@gsau.edu.cn
羅建勛,男,研究員,E-mail:vectorparasit@126.com;孫曉林,男,教授,E-mail:sunxl@gsau.edu.cn
S852.72
A
0366-6964(2016)06-1293-06
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
數學小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中學生數理化·七年級數學人教版(2019年4期)2019-05-20 10:06:32
中學生數理化·七年級數學人教版(2018年6期)2018-06-26 08:36:06
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
