滇龍膽乙酰CoA轉移酶基因的克隆與表達分析
2016-07-25 18:03:06張曉東李彩霞王連春王元忠
江蘇農業科學 2016年6期
張曉東+李彩霞+王連春+王元忠

摘要:根據滇龍膽轉錄組乙酰CoA酰基轉移酶基因GrAACT序列設計引物,使用逆轉錄PCR(RT-PCR)技術從滇龍膽幼葉擴增該基因,并進行序列分析、原核表達和組織特異性分析。結果表明:GrAACT基因(登錄號KJ917163)開放閱讀框長 1 215 bp,編碼404個氨基酸;GrAACT蛋白相對分子質量41.41 ku,pI值為6.25,屬于AACT蛋白家族成員,無信號肽,為疏水穩定蛋白,主要由α-螺旋、無規則卷曲構成;GrAACT蛋白具有AACT蛋白保守結構域:硫解酶N端結構域(第12~272位氨基酸)、硫解酶C端結構域(第282~402位氨基酸)、類硫解酶結構域(第13~403位氨基酸);在硫解酶C端結構域中,包含硫解酶保守位點(第350~366位氨基酸)、硫解酶活性位點(第385~398位氨基酸);滇龍膽GrAACT蛋白與長春花CrAACT蛋白親緣關系最近;GrAACT基因在大腸桿菌中表達的重組蛋白相對分子質量約為6741 ku;組織特異性表達分析結果表明,GrAACT基因主要在莖、根中表達。
關鍵詞:滇龍膽;乙酰CoA酰基轉移酶;基因克隆;生物信息學分析;表達分析
中圖分類號: S184;R282.71;Q785文獻標志碼: A文章編號:1002-1302(2016)06-0094-05
收稿日期:2015-05-27
基金項目:國家自然科學基金(編號:81260608);國家科技支撐計劃(編號:2011BAI13B02-04);云南省教育廳科學研究基金重點項目(編號:2015Z171)。
作者簡介:張曉東(1980—),男,河南新鄭人,博士,副教授,從事植物代謝基因工程研究。E-mail:zxd95@126.com。
通信作者:王元忠,碩士,助理研究員,從事藥用植物資源評價與利用的研究。E-mail:boletus@126.com。滇龍膽是傳統中藥龍膽的來源植物之一,也是云南省重要道地藥材,主要分布于中國西南部,尤其是云南省山區地帶;其根入藥,用于治療各種炎癥、肝炎、風濕病和膽囊炎等[1]。目前,醫用龍膽需求量每年遞增約10%,而現有滇龍膽產量低、品質不高,且野生滇龍膽資源遭到人為大肆采挖[2]。為選育優質、高產、高抗新品種,2013年6月云南省已啟動龍膽草航天育種工程[3]。從中醫藥現代化角度來看,滇龍膽主要藥效成分為龍膽苦苷,要從根本上解決龍膽草藥源問題并保護野生滇龍膽,除篩選新品種之外,還必須弄清龍膽苦苷的生物合成途徑及其調控機理,為通過基因工程手段生產龍膽苦苷奠定基礎。
龍膽苦苷為裂環烯醚萜類化合物,在植物中主要通過質體2-C-甲基-D-赤蘚糖醇-4-磷酸(MEP)、胞質甲羥戊酸(MVA)途徑合成[4]。乙酰CoA酰基轉移酶(又稱乙酰乙酰CoA硫解酶,AACT,EC 2.3.1.9)是MVA途徑中的第1個限速酶,能夠催化2分子乙酰CoA形成1分子乙酰乙酰CoA[5]。目前,乙酰CoA酰基轉移酶基因AACT已從擬南芥[5]、丹參[6]、胡蘿卜[7]、白木香[8]、油茶[9]等許多植物中分離。AACT基因表達具有組織特異性,并被生物、非生物因素誘導。擬南芥中存在2個平行進化的同源基因,其中AtAACT1主要在維管系統中表達,AtAACT2則在根尖、幼葉、莖上部和花藥中大量表達,二者存在功能冗余;AtAACT2的作用是為胞質定位、MVA起源的異戊烯萜類生物合成途徑合成大量乙酰乙酰CoA前體[5]。在生物誘導劑(100 mg/mL酵母提取物)、非生物誘導劑(30 mmol/L Ag+)共同誘導下,丹參SmAACT基因在誘導后12 h的表達量達到最高值[10]。
目前,由于龍膽基因組資源極度匱乏[11],導致龍膽苦苷生物合成途徑并不清楚[3]。本研究根據筆者實驗室滇龍膽轉錄組數據庫中的GrAACT基因序列,設計特異性引物,通過逆轉錄PCR(RT-PCR)技術成功從滇龍膽幼葉中擴增到GrAACT基因,并進行序列分析、原核表達和組織表達特異性分析,以期為闡明滇龍膽龍膽苦苷生物合成途徑及其調控機理奠定基礎。
1材料與方法
1.1材料
滇龍膽組培苗、盆栽苗均采自玉溪師范學院分子生物學實驗室,經云南省農業科學院藥用植物研究所金航研究員鑒定為滇龍膽(Gentiana rigescens Franch. ex Hemsl)。基因克隆所用材料為滇龍膽無菌苗幼葉,基因組織特異性表達分析所用材料為盆栽3年生滇龍膽的根、莖、葉,采樣日期為2014年5月17日。
1.2方法
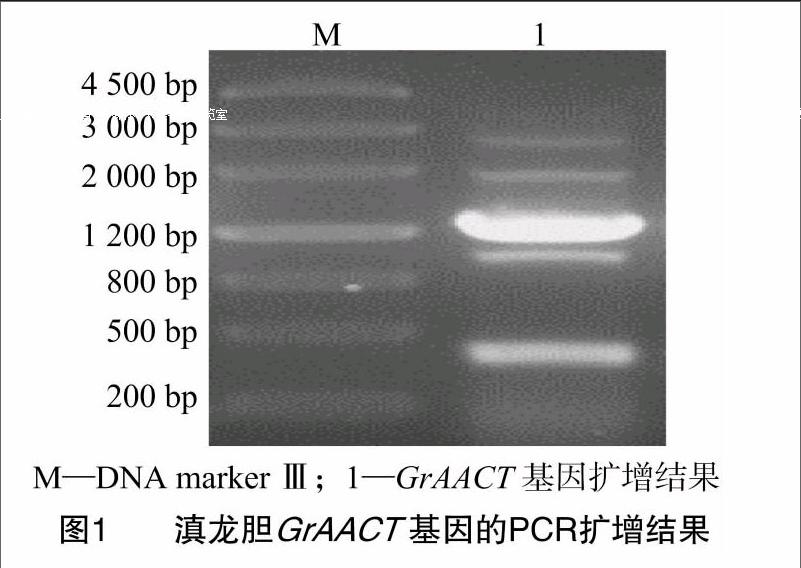
按照多糖植物組織提取試劑RNAiso(TaKaRa,大連)說明書提取滇龍膽幼葉總RNA;按照逆轉錄試劑盒(TaKaRa,大連)說明書合成cDNA。根據原核表達載體pGEX-4T-1多克隆酶切位點和筆者所在實驗室前期測序的滇龍膽轉錄組GrAACT基因序列,設計1對特異引物GrAACT BamHⅠ-F、GrAACT XhoⅠ-R(表1)。以cDNA為模板進行PCR擴增,反應體系為:0.5 μL TransTaq Taq DNA Polymerase High Fidelity(2.5 U/μL,北京全式金生物技術有限公司),5 μL 10×TransTaq HiFi Buffer Ⅱ,4 μL dNTP(2.5 mmol/L,TaKaRa,大連),2 μL模板,各1 μL正反向引物(10 μmol/L),加dd H2O補足至50 μL。PCR反應條件為:94 ℃ 3 min;94 ℃ 30 s,55.5 ℃ 30 s,72 ℃ 75 s,30個循環;72 ℃ 10 min。PCR產物經1.0%瓊脂糖凝膠電泳分離,然后割膠,使用膠回收試劑盒(Qiagen,德國)對目的片段進行回收,將其連接到pMD19-T載體(TaKaRa,大連)。轉化大腸桿菌DH5α(TaKaRa,大連)后進行藍白斑篩選,挑取12個白斑搖菌;使用試劑盒提取質粒(北京百泰克生物技術有限公司),經酶切檢測正確后進行測序[生工生物工程(上海)股份有限公司],獲得重組質粒pMD19-GrAACT。
1.2.1GrAACT基因的生物信息學分析用NCBI網站的BLAST程序進行序列比對,用Genetyx 6.1.8進行翻譯,用DNAMAN 7進行多序列比對,用Clustal X2.1進行比對,然后用MEGA 6.0軟件內置的NJ法構建系統進化樹,設置Bootstrap=1 000;利用在線數據庫(http://molbiol. edu.ru/eng/scripts/01_11.html)進行稀有密碼子分析。用ChloroP服務器v1.1進行葉綠體轉運肽預測;用Interpro軟件進行保守結構域預測;用ProtScalee軟件進行疏水性分析;用PredictProtein對二級結構進行預測;用Phyre2(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)對三級結構進行預測;用Expasy中的TMHMM工具預測蛋白的跨膜螺旋區;用在線工具WOLF PSORT預測蛋白的亞細胞定位情況。
1.2.2GrAACT基因的熒光實時定量分析分別取3年生滇龍膽的根、莖、葉,提取總RNA,用DNase I處理除去基因組DNA。用反轉錄試劑盒(TaKaRa)合成第1鏈cDNA。以轉錄組中GrGAPDH基因(GenBank登錄號KM061807)作為內參設計特異性引物(表1),PCR條件:95 ℃ 3 min,95 ℃ 15 s,60 ℃ 31 s。根據GrAACT基因開放閱讀柜(ORF)序列設計特異性引物(表1),使用SuperReal PreMix Plus試劑盒[天根生化科技(北京)有限公司]進行qPCR,擴增條件為:95 ℃ 3 min,95 ℃ 15 s,60 ℃ 31 s;每個反應重復3次。反應在ABI7000熒光定量PCR儀(Applied Biosystems)上進行擴增,擴增曲線、溶解曲線、標準曲線由定量PCR儀軟件自動生成。使用內參基因GrGAPDH表達校準后,計算根、莖、葉中GrAACT基因的相對表達量。采用比較CT值的“2-ΔΔCT”的方法進行定量數據的分析處理。
2結果與分析
2.1滇龍膽GrAACT基因的克隆
以滇龍膽幼葉cDNA為模板,使用表1中的基因特異性引物GrAACTBamHI-F、GrAACTBamHI-R擴增出約 1 200 bp 的片段,詳見圖1。通過TA克隆獲得重組質粒 pMD19-GrAACT。
2.2GrAACT基因的生物信息學分析
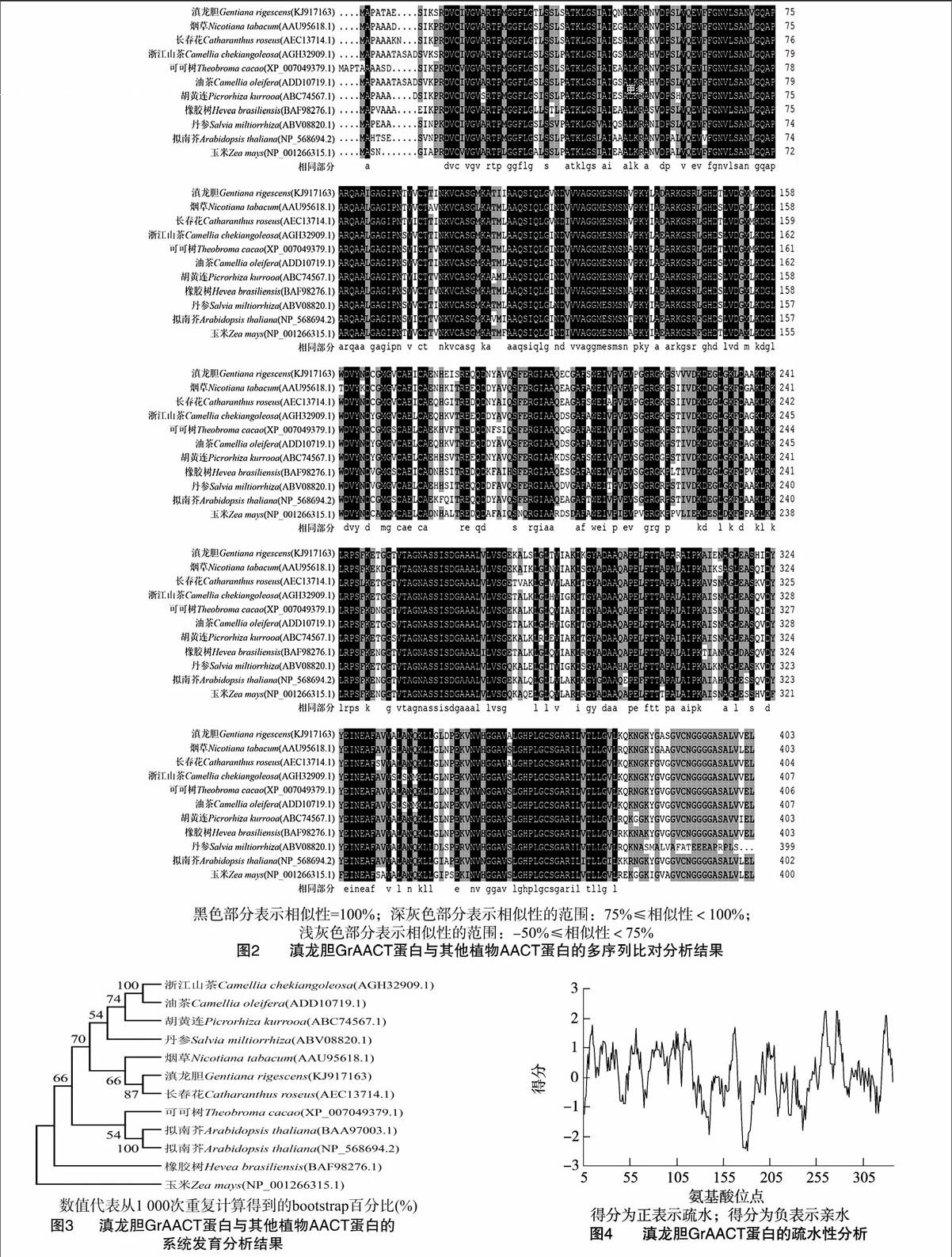
2.2.1GrAACT基因的ORF分析和多序列比對分析利用Genetyx軟件對GrAACT基因的ORF序列進行分析,結果顯示:該基因(GenBank登錄號KJ91716)長1 215 bp,編碼404個氨基酸。利用GenBank數據庫的BLASTp程序對GrAACT蛋白進行比對分析,結果表明:滇龍膽GrAACT蛋白與長春花CrAACT蛋白的序列相似性最高(88.61%),與單子葉植物玉米ZmAACT蛋白的相似性(78.55%)稍低。利用DNAMAN 7將GrAACT蛋白序列與NCBI中相似性較高的部分序列進行多序列比對分析,結果表明:GrAACT蛋白與已知蛋白序列相比保守性較高(圖2)。利用Mega 6.0將GrAACT氨基酸序列與NCBI中相似性較高的部分序列進行系統發育分析,結果顯示:滇龍膽GrAACT蛋白與長春花CrAACT蛋白處于同一進化支(圖3),表明二者親緣關系較近。
2.2.2GrAACT蛋白的理化特性分析使用ExPASy ProtParam tool對GrAACT蛋白進行分析,結果表明:GrAACT蛋白單體相對分子質量為41 410 u,pI值為6.25,這與丹參SmAACT蛋白[6]、油茶CoAACT蛋白[9]、擬南芥ATAACT2蛋白[5]類似;帶正電氨基酸殘基(Arg+Lys)數為36個,帶負電氨基酸殘基(Asp+Glu)數為38個,化學式為:C1812H2959N509O562S17;不穩定指數為27.92,屬穩定蛋白;脂肪指數為9856,總平均疏水性(GRAVY)為0.178,為疏水蛋白(圖4)。GrAACT蛋白含20種基本氨基酸,其中丙氨酸含量最高,為13.9%;其次是甘氨酸、纈氨酸,含量分別為11.9%、9.9%;色氨酸含量最低,為0.5%。
2.2.3GrAACT蛋白的二級結構、三級結構分析利用SSpro方法對GrAACT蛋白進行二級結構分析,結果表明:該蛋白二級結構中α-螺旋(H)占39.11%,無規則卷曲(C)占40.10%,延伸帶(E)占20.79%。利用Swiss-Model Workspace的自動模式預測GrAACT蛋白的三級結構,結果見圖5,該模型以人線粒體乙酰CoA酰基轉移酶[2f2s.1.C]為模板,在第13—404位氨基酸處建模,序列相似度為52.73%。
2.2.4GrAACT蛋白保守結構域分析使用InterPro在線工具對GrAACT蛋白的保守結構域進行分析,結果表明:GrAACT
蛋白包含3個保守結構域,它們分別為硫解酶N端結構域(IPR020616,第12—272位氨基酸)、硫解酶C端結構域(IPR020617,第282—402位氨基酸)、類硫解酶結構域(IPR016039,第13—403位氨基酸)(圖6)。在硫解酶C端結構域中,包含硫解酶保守位點(THIOLASE_2,第350—366位氨基酸)、硫解酶活性位點(THIOLASE_3,第385—398位氨基酸)。
2.2.5GrAACT蛋白信號肽分析利用ExPASy SignalP 4.1服務器分析GrAACT蛋白,未發現信號肽,表明該蛋白為非分泌型蛋白。利用TMHMM工具預測GrAACT蛋白的跨膜螺旋區,結果表明:GrAACT蛋白不含有跨膜區域(圖7)。
2.2.6GrAACT蛋白亞細胞定位分析使用WoLF PSORT軟件進行亞細胞定位分析,結果表明,GrAACT蛋白在細胞質、葉綠體、過氧化物酶體的定位系數分別為6.0、4.0、3.0。
2.2.7GrAACT基因稀有密碼子分析使用在線軟件對GrAACT基因進行稀有密碼子分析,結果表明:GrAACT基因中稀有密碼子占1.48%,且無二聯或三聯稀有密碼子連續出現的情況,因此可選用大腸桿菌表達菌BL21或Rosetta(DE3)進行原核表達。
2.3GrAACT基因原核表達載體的構建
使用酶切檢測重組質粒pGEX-4T-1-GrAACT,結果表明:BamHⅠ、XhoⅠ雙酶切可切出目的基因和載體,且二者長度之和等于XhoⅠ單酶切產物長度(圖8),表明GrAACT基因已成功插入載體pGEX-4T-1。
2.4GrAACT基因的原核表達
將重組質粒pGEX-4T-1-GrAACT轉化大腸桿菌Rosetta(DE3)后,在37 ℃、1 mmol/L IPTG(終濃度)下,分別
誘導表達0、2、4、6 h后,提取細菌總蛋白進行SDS-PAGE分析。結果表明:與對照(第1~3泳道)相比,pGEX-4T-1-GrAACT轉化菌經IPTG誘導后,在相對分子質量67 410 u(含GST蛋白26 000 u)左右有1條蛋白條帶,并且隨誘導時間增加,其蛋白含量逐漸升高(圖9),表明重組質粒pGEX-4T-1-GrAACT在大腸桿菌Rosetta(DE3)中成功誘導表達了GrAACT蛋白。當溫度為37 ℃、誘導時間為6 h時,蛋白表達量最大(圖9),細胞破碎后可直接用于蛋白純化。
2.5GrAACT基因的組織表達分析
取3年生滇龍膽的根、莖、葉,提取總RNA,反轉錄成cDNA后,通過實時熒光定量RT-PCR分析GrAACT基因在根、莖、葉中的表達情況。圖10結果表明:GrAACT基因在莖中表達量最高,分別約為葉、根的13.17、1.36倍,表達量最低的是葉。
3討論
滇龍膽藥效成分龍膽苦苷是通過MVA、MEP途徑合成的,在前期筆者已經克隆了MEP途徑關鍵酶1-脫氧-D-木酮糖-5-磷酸還原異構酶基因GrDXR[12]。為闡明龍膽苦苷生物合成途徑及其調控機理,本研究克隆了MVA途徑中的GrAACT基因。AACT是MVA途徑的第1個催化酶,因此滇龍膽中GrAACT基因的表達情況可以直接或間接影響龍膽苦苷的生物合成。研究表明,植物中如丹參[6]、白木香[8]、油茶[9]AACT蛋白均包含硫解酶N端結構域、C端結構域和保守位點。本研究從滇龍膽幼葉中克隆獲得GrAACT基因,其編碼蛋白序列比對分析結果表明,GrAACT蛋白屬于AACT家族蛋白,具有硫解酶N端、C端結構域,并且在其C端具有保守位點:NVHGGAVALGHPLGCSG(第350—366位氨基酸)。這些結果表明,所克隆基因的確為滇龍膽乙酰CoA酰基轉移酶基因。
根據底物鏈長特異性,硫解酶被分為2類:第I類是降解型硫解酶(EC 2.3.1.16),定位于線粒體、過氧化物酶體,在脂肪酸氧化中催化3-酮脂酰CoA斷裂產生乙酰乙酰CoA、縮短的酰基CoA;第Ⅱ類是合成型硫解酶(EC 2.3.1.9),定位于細胞質,能夠催化2分子乙酰CoA形成1分子乙酰乙酰CoA[13]。生物信息學分析結果表明,GrAACT蛋白定位于細胞質,屬于Ⅱ型硫解酶,參與胞質中的MVA途徑。在植物中,AACT蛋白的定位還存在爭議。在擬南芥中,參與MVA生物合成途徑的AtAACT2蛋白定位于細胞質[14];而對擬南芥過氧化物酶體蛋白質組研究結果表明,該蛋白也存在于過氧化物酶體[15]。在向日葵(Helianthus annuus)乙醛酸循環體中,HaAACT能夠催化朝向乙酰乙酰CoA方向的反應[16],說明過氧化物酶體中也存在AACT。GrAACT蛋白三維模型預測結果表明,該蛋白的活性形式為四聚體。在本研究中,GrAACT單體相對分子質量為41 410 u,其活性形式可能是單體,也可能是四聚體,這需要通過非變性SDS-PAGE進行進一步檢測。
在擬南芥中,參與MVA途徑的AtAACT2基因在根尖、幼葉、莖上部和花藥中大量表達[5]。組織表達特異性檢測結果表明,GrAACT基因在根中的表達量較高,因此推測:滇龍膽根部為龍膽苦苷生物合成的主要器官。滇龍膽用藥部分為根,因為與葉、莖相比,根中龍膽苦苷含量最高[17-18],這也為上述假說提供了證據。在擬南芥中,參與MVA途徑的AtAACT2基因是多效基因。首先,AtAACT2對于胚胎形成和正常的雄性配子傳遞是必需的,AtAACT2突變體不能存活;其次,AtAACT2基因的RNAi株系表現出多效的表型,包括減弱的頂端優勢、延長的生活周期和開花時間、雄性不育、矮化、種子產量降低、根長變短和植物甾醇類質和量的改變等;再次,當外源添加甲羥戊酸后,AtAACT2基因的RNAi株系以上的表型和生化改變都可逆轉[5]。本研究中的GrAACT蛋白與AtAACT2蛋白相似性高達83.87%,GrAACT基因是否也為多效基因,需要進一步試驗驗證。
本研究為滇龍膽的分子生物學研究提供基因資源,并為GrAACT基因功能的解析奠定基礎。下一步將對GrAACT蛋白進行純化和酶活分析,通過互補試驗或過表達研究GrAACT基因的功能,從而為龍膽苦苷生物合成途徑的闡明奠定基礎。
參考文獻:
[1]Suyama Y,Kurimoto S I,Kawazoe K,et al. Rigenolide A,a new secoiridoid glucoside with a cyclobutane skeleton,and three new acylated secoiridoid glucosides from Gentiana rigescens Franch[J]. Fitoterapia,2013,91:166-172.
[2]金航,張霽,張金渝,等. 滇龍膽[M]. 昆明:云南科技出版社,2013.
[3]Zhang X,Allan A C,Li C,et al. De novo assembly and characterization of the transcriptome of the Chinese medicinal herb,Gentiana rigescens[J]. International Journal of Molecular Sciences,2015,16(5):11550-11573.
[4]Hua W,Zheng P,He Y,et al. An insight into the genes involved in secoiridoid biosynthesis in Gentiana macrophylla by RNA-seq[J]. Molecular Biology Reports,2014,41(7):4817-4825.
[5]Jin H,Song Z,Nikolau B J. Reverse genetic characterization of two paralogous acetoacetyl CoA thiolase genes in Arabidopsis reveals their importance in plant growth and development[J]. The Plant Journal,2012,70(6):1015-1032.
[6]崔光紅,王學勇,馮華,等. 丹參乙酰CoA酰基轉移酶基因全長克隆和SNP分析[J]. 藥學學報,2010,45(6):785-790.
[7]Bach T J,Boronat A,Campos N,et al. Mevalonate biosynthesis in plants[J]. Critical Reviews in Biochemistry & Molecular Biology,1999,34(2):107-122.
[8]劉娟,徐艷紅,楊勇,等. 白木香乙酰乙酰基輔酶A硫解酶基因(AsAACT)的克隆與表達分析[J]. 中國中藥雜志,2014,39(6):972-980.
[9]張琳,譚曉風,胡姣,等. 油茶乙酰CoA酰基轉移酶基因 cDNA 克隆及序列特征分析[J]. 中南林業科技大學學報,2011,31(8):108-112.
[10]Gao W,Sun H X,Xiao H,et al. Combining metabolomics and transcriptomics to characterize tanshinone biosynthesis in Salvia miltiorrhiza[J]. BMC Genomics,2014,15(1):1-14.
[11]Nakatsuka T,Yamada E,Saito M,et al. Construction of the first genetic linkage map of Japanese gentian (Gentianaceae)[J]. BMC Genomics,2012,13(1):1-15.
[12]張曉東,趙靜,李彩霞,等. 滇龍膽1-脫氧-D-木酮糖-5-磷酸還原異構酶基因GrDXR的克隆、序列分析與原核表達[J]. 中草藥,2014,45(16):2378-2384.
[13]Meng Y,Li J. Cloning,expression and characterization of a thiolase gene from Clostridium pasteurianum[J]. Biotechnology Letters,2006,28(16):1227-1232.
[14]Carrie C,Murcha M W,Millar A H,et al. Nine 3-ketoacyl-CoA thiolases (KATs) and acetoacetyl-CoA thiolases (ACATs) encoded by five genes in Arabidopsis thaliana are targeted either to peroxisomes or cytosol but not to mitochondria[J]. Plant Molecular Biology,2007,63(1):97-108.
[15]Reumann S,Babujee L,Ma C,et al. Proteome analysis of Arabidopsis leaf peroxisomes reveals novel targeting peptides,metabolic pathways,and defense mechanisms[J]. The Plant Cell,2007,19(10):3170-3193.
[16]Oeljeklaus S,Fischer K,Gerhardt B. Glyoxysomal acetoacetyl-CoA thiolase and 3-oxoacyl-CoA thiolase from sunflower cotyledons[J]. Planta,2002,214(4):597-607.
[17]楊美權,張金渝,沈濤,等. 不同栽培模式對滇龍膽中龍膽苦苷含量的影響[J]. 江蘇農業科學,2011(1):287-289.
[18]王彩云,張曉東,沈濤,等. 龍膽苦苷生物合成途徑研究進展[J]. 江蘇農業科學,2014,42(3):4-10.
