低溫對不同萌發狀態裸燕麥種子生長生理特性的影響
2016-07-25 14:42:14張麗賈志國
江蘇農業科學 2016年6期
張麗++賈志國

摘要:將3種萌發狀態(吸脹、萌動、發芽)的裸燕麥種子分別于0 ℃下處理24、48、72 h,測定裸燕麥種子發芽及各項生長、生理指標。結果表明:隨低溫脅迫時間的延長,發芽率、發芽勢、發芽指數及活力指數均顯著降低。與對照相比,低溫脅迫顯著抑制了根長、芽長、鮮質量、干質量及根冠比,其中48 h時間處理下,根長、鮮質量、干質量及根冠比受抑制程度最小。各處理下的相對電導率、丙二醛含量及可溶性蛋白含量表現為不同程度的增加,脯氨酸含量顯著降低。綜上所述,裸燕麥對24、48 h的低溫處理有一定的忍耐能力,3種萌發狀態的裸燕麥種子抗寒能力從強到弱依次為發芽種子> 吸脹種子> 萌動種子。
關鍵詞:裸燕麥;低溫脅迫;萌發;生理指標;抗寒性
中圖分類號: S512.601文獻標志碼: A文章編號:1002-1302(2016)06-0161-04
收稿日期:2015-04-23
基金項目:河北北方學院自然科學研究計劃。
作者簡介:張麗(1979—),女,博士,講師,從事作物與經濟林栽培育種工作。E-mail:yingrizl@126.com。裸燕麥(Avena nuda L.)別稱莜麥、鈴鐺麥,禾本科燕麥屬一年生草本植物[1]。裸燕麥性喜涼爽,耐干旱、抗鹽堿、生長周期短,是我國華北、西北等高寒地區的主要糧飼兼用作物[2],其營養價值高,富含人體必需的8種氨基酸,是北方地區人們喜愛的特色食物,其秸稈也是優質的牧草[3]。近年來研究發現,裸燕麥具有降脂、降糖的功效,被作為一種營養保健功能食品加以開發利用[4],因此裸燕麥是一種多用途、高經濟價值的作物。
我國海拔較高的高寒地區為裸燕麥的適宜種植區。裸燕麥是禾本科中抗寒性較強的作物,萌芽期是其生長發育過程中最關鍵的時期,此時由于其抗寒能力強,能抵抗一定的低溫脅迫,但當溫度過低,超過其所能忍受的極限低溫時便會造成脅迫,影響種子發芽率和發芽勢,造成出苗不齊,從而影響其最終產量。前人關于裸燕麥抗旱性研究較多[5-7],對其抗寒機理的研究較少。本試驗研究了同一低溫不同處理時間脅迫對不同萌發狀態裸燕麥種子的影響,從萌發、生長、生理3個方面探討了裸燕麥對低溫脅迫的響應,揭示了裸燕麥抗寒生理機制,為我國北方嚴寒地區裸燕麥高產栽培及進一步推廣提供理論依據。
1材料與方法
1.1試驗材料與設計
選擇裸燕麥常規品種花早2號為試材。挑選大小一致、籽粒飽滿的種子,分成36組,每組100粒,經清水浸種24 h后,在20 ℃進行萌發試驗,將種子處理為吸脹、萌動、發芽3種萌發狀態,分別進行低溫脅迫發芽試驗。將不同萌發狀態的裸燕麥種子置于0 ℃低溫下,分別處理24、48、72 h,3個時間處理低溫脅迫結束后,轉移至20 ℃繼續培養;對照始終保持20 ℃。重復3次。
1.2試驗及測定方法
1.2.1發芽試驗方法種子低溫脅迫萌發試驗根據顏啟傳的方法[8]進行改進。取足量種子放入大燒杯中浸泡24 h,然后放入經過消毒的大瓷盤中,用濕布覆蓋,放在陰蔽處培養,供試驗選用。首先于15 cm培養皿內鋪2層濾紙作為發芽床,選取浸泡后的均勻飽滿種子12組,每組100粒,均勻擺放,作為吸脹組試驗材料。將培養皿置于0 ℃光照培養箱分別處理24、48、72 h,每天補充適量的蒸餾水,以保持滲透勢不變,分別測定各項生理指標。隨著試驗的進行,再于瓷盤中分別挑選處于萌動和發芽狀態的種子進行上述試驗,每處理重復3次。
1.2.2發芽指標的測定從上述3種萌發狀態的種子中選擇萌動處理組為代表,進行標準發芽試驗,每天記錄發芽數,于試驗后4 d計算發芽勢,8 d后試驗結束,計算發芽率、發芽指數及活力指數。
1.2.3生長指標的測定將“1.2.2”節發芽結束后的各處理在20 ℃下培養15 d,利用常規方法測定裸燕麥幼苗生長初期根長、苗高、根鮮質量、苗鮮質量、干質量,計算根冠比。
1.2.4生理指標的測定相對電導率采用電導率法測定。稱取0.5 g樣品,加入10 mL水,浸泡24 h,測定其浸出液電導率,煮沸15 min,冷卻后再次測定其電導率,將2次電導率代入公式,記算結果;膜傷害指標丙二醛(MDA)含量采用硫代巴比妥酸(TBA)比色法[9]測定;游離脯氨酸含量采用酸性茚三酮顯色法[9]測定;可溶性蛋白含量采用考馬斯亮藍 G-250 染色法[10]測定。
1.3數據分析
應用Microsoft Excel軟件進行數據整理和繪圖,SPSS 160軟件進行數據統計分析,用單因素方差分析(One-Way ANOVA)和最小顯著差異法(LSD)比較不同處理間的差異。
2結果與分析
2.1低溫脅迫對裸燕麥種子發芽的影響
由表1可以看出,隨著低溫脅迫時間的延長,裸燕麥種子的發芽率、發芽勢、發芽指數及活力指數均呈現逐漸降低的趨勢,與20 ℃對照相比差異顯著。24、48、72 h的低溫脅迫后發芽率分別降低為對照的90.0%、61.0%、46.6%,且處理間差異顯著。發芽勢的變化規律與發芽率基本一致,72 h的低溫脅迫導致發芽勢降低為對照的39.6%,表明相比發芽率低溫脅迫對發芽勢的影響更大。24 h的低溫處理也顯著降低了發芽指數。活力指數則表現為極顯著差異(P<0.01),隨著脅迫時間的延長,48、72 h脅迫下,發芽指數和活力指數分別降低為對照的57.1%、34.0%和31.4%、14.9%,說明低溫脅迫一定程度上抑制了裸燕麥的萌發,活力指數的變化可以看出即使能夠萌發的那部分種子,其繼續生長的活力也受到了明顯的影響。
2.2低溫脅迫對裸燕麥幼苗苗高、根長、鮮質量、干物質及根冠比的影響
溫度是植物種子萌發出苗的基本條件之一,低溫脅迫導致萌動發芽后的種子生長緩慢甚至停止生長[11]。本研究結果(表2)表明,裸燕麥幼苗的根長、芽長均受到了低溫抑制,與對照相比均顯著降低,隨著低溫脅迫時間的延長,根長表現為先升后降的趨勢,48 h的低溫脅迫反而使根長增加,與24 h處理相比,48 h處理下芽長變化不顯著,72 h的低溫處理對芽長的抑制作用進一步加強。3種低溫時間處理下,根鮮質量顯著低于對照,24、72 h低溫處理后的芽鮮質量也顯著降低,相比而言,低溫脅迫對根的抑制作用更明顯。隨低溫脅迫時間的增加,48 h處理后的根鮮質量和芽鮮質量均略有增加,72 h處理又繼續下降,但處理間差異不顯著。裸燕麥幼苗干物質受到低溫的顯著抑制,0 ℃處理72 h后的種子萌發后的幼苗干質量僅為對照的45.5%。與對照相比,根冠比與干質量的變化規律基本一致,均低于對照,72 h處理后降低為對照的36.4%,3個時間處理之間對比發現,48 h處理后的干質量和根冠比的平均值最大,與對照不顯著,表明裸燕麥對低溫脅迫有一定的適應抵抗能力。
2.3不同低溫脅迫時間對裸燕麥種子生理指標的影響
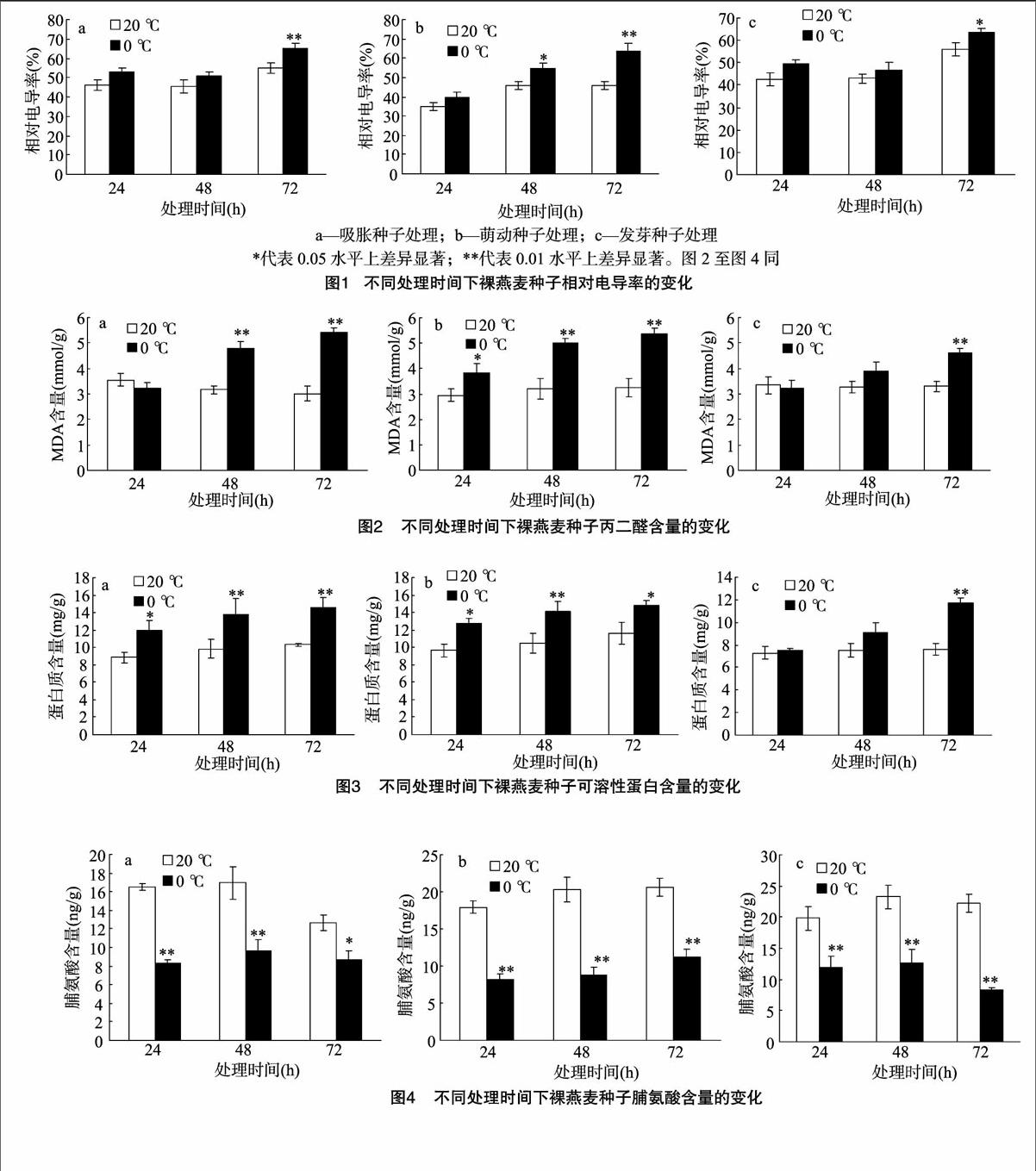
2.3.1不同低溫脅迫時間下裸燕麥種子相對電導率的變化由圖1可知,與對照相比,低溫處理后不同萌發狀態的裸燕麥種子相對電導率均有所增加。圖1-a所示低溫處理72 h的吸脹種子相對電導率極顯著增加,而24、48 h處理下增加不明顯。隨著0 ℃低溫脅迫時間的延長,萌動種子相對電導率逐漸增加(圖1-b),與各自對照相比,48 h處理達顯著差異,72 h處理差異極顯著。發芽種子在低溫處理72 h后也顯著增加(圖1-c)。吸脹、萌動、發芽3種萌發狀態的裸燕麥種子72 h處理后電導率的相對增加量分別為10.2%、18.1%、7.6%,表明發芽種子抗低溫能力較強,而萌動種子作為種子發芽的敏感時期對低溫抵抗能力最弱。
2.3.2不同低溫脅迫時間下裸燕麥種子丙二醛含量的變化植物在逆境脅迫過程中產生過剩的自由基,引發膜脂過氧化作用,膜脂過氧化的中間產物自由基和最終產物MDA都會造成膜系統的損傷[12]。由圖2可以看出,隨低溫處理時間的延長,不同萌發狀態的裸燕麥種子丙二醛含量也會增加。與各自對照相比,48、72 h處理后的吸脹和萌動種子差異極顯著(圖2-a、圖2-b)。圖2-c所示,24、48 h的低溫處理時間下發芽狀態種子的丙二醛含量差異不顯著,當脅迫時間延長至72 h時其含量顯著增加。24 h低溫脅迫后吸脹和發芽種子丙二醛含量無明顯變化,說明裸燕麥對短時間的低溫脅迫有一定的忍耐能力。
2.3.3不同低溫脅迫時間下裸燕麥種子可溶性蛋白含量的變化可溶性蛋白作為滲透調節物質、能量物質和信息傳遞物質在植物抵御逆境過程中起著重要的作用[13]。由圖3可知,經過低溫脅迫處理的裸燕麥種子中可溶性蛋白含量均高于對照,說明低溫脅迫造成種子中可溶性蛋白含量升高。3種低溫處理時間下,處于吸脹和萌動時期的裸燕麥種子可溶性蛋白含量均顯著高于對照(圖3-a、圖3-b)。發芽狀態的裸燕麥種子在處理時間為72 h時,可溶性蛋白含量顯著高于對照,24、48 h處理比對照略有增加,但差異不顯著。72 h 低溫脅迫處理下,吸脹、萌動、發芽3種萌發狀態可溶性蛋白增加量分別為各自對照的1.41、1.24、1.54倍。萌動時期蛋白增加量最低,表明此時期的裸燕麥種子對低溫更敏感,抗寒性較弱。
2.3.4不同低溫脅迫時間下裸燕麥種子脯氨酸含量的變化由圖4可知,與各自對照相比,3種萌發狀態下的裸燕麥種子低溫脅迫后的脯氨酸含量均顯著降低。不同萌發狀態種子處理之間對比發現,24、48 h低溫脅迫后,發芽狀態下脯氨酸積累最多,其平均值分別為吸脹、萌動種子的1.37、1.46倍,表明發芽狀態的裸燕麥種子在一定時間的低溫脅迫范圍內,脯氨酸可以迅速積累以抵抗逆境條件。
3結論與討論
各種作物種子對發芽要求一定的溫度,過低的溫度會抑制種子的萌發[14-15]。本研究中低溫脅迫抑制了裸燕麥種子萌發,隨著低溫處理時間的延長,發芽勢逐漸下降,發芽總數也顯著減少。不同低溫處理時間下裸燕麥種子的發芽率、發芽勢、發芽指數及活力指數均低于對照,但活力指數下降的最快,表明低溫脅迫在一定程度上抑制了萌發,而仍能萌發的那部分種子活力也大大降低了。裸燕麥芽苗期的生長指標也受到低溫的影響,持續低溫阻礙了芽和根的伸長,48 h低溫處理后的各生長指標的平均值均高于其他2種時間處理,表明恢復正常溫度后,裸燕麥仍能正常生長,低溫處理48 h后的種子在適宜條件下生長最快,處理裸燕麥對持續低溫48 h有一定的忍耐能力,溫度回升后尚能恢復,但當低溫脅迫時間持續超過72 h,種子發芽生長已比較困難,只能勉強維持生命活動。本試驗還發現,24 h低溫脅迫下的生長恢復能力低于 48 h 處理,對于抗寒性較強的裸燕麥來說可能存在一個低溫鍛煉的問題,48 h的持續低溫使幼苗體內積累了更多的干物質,從而保持了一定的生長量。前人關于小麥的研究中發現,干旱脅迫雖然導致了不同萌發狀態種子的傷害,但并不喪失其自身的修復和恢復生長的能力[16]。
在低溫冷害條件下,植物由于細胞膜內大量溶質外滲導致了其浸出液的電導率增加,表明細胞膜遭受了破壞,電導率增加越少,表明抗寒性越強[17]。經過24、48 h低溫處理的裸燕麥膜透性增加的程度較小,尤其是吸脹種子和發芽種子其數值雖略有增加,但與對照差異不顯著,溫度回升后靠自身的修復能力可以恢復正常。經過72 h低溫處理的裸燕麥膜透性增加程度較大,細胞內電解質大量外滲,電導率顯著增加。分析電導率指標得出,裸燕麥可以忍耐24、48 h的持續低溫,且發芽種子的耐寒能力大于吸脹種子和萌動種子。低溫處理后,3種不同萌發狀態的裸燕麥種子丙二醛、可溶性蛋白含量表現為不同程度的增加,經分析得出與電導率指標基本一致的結果,其抗寒能力從強到弱依次為發芽種子>吸脹種子>萌動種子。萌動狀態的種子抵御低溫脅迫的能力較差,該時期對外界環境條件非常敏感,各種生理活性物質及代謝反應開始趨于旺盛,持續低溫干擾了萌發的進程,其受傷害程度也較大。
脯氨酸含量作為抗寒性指標前人已有大量報道[18-19]。一般認為,脯氨酸積累越多其抗寒性越強,但也有不一致的觀點,認為脯氨酸與抗寒性關系不大[20]。本試驗發現了與前人研究不一致的結果,各處理組脯氨酸含量均顯著低于對照,可能是由于裸燕麥種子蛋白質含量豐富,20 ℃正常萌發時,蛋白質大量分解為可供幼苗利用的游離氨基酸,而0 ℃處理抑制了蛋白水解酶的活性,從而導致對照組脯氨酸含量大于處理組,抵消了低溫脅迫后脯氨酸的積累作用。如果忽略對照組,單純分析低溫脅迫處理發現,發芽狀態的脯氨酸積累量大于吸脹和萌動的種子,與上述指標分析結果一致。因此,脯氨酸含量可以作為裸燕麥抗寒性的指標。
綜上所述,裸燕麥萌發時期抗低溫能力較強,且發芽種子>吸脹種子>萌動種子。從吸脹、萌動到發芽不同萌發狀態的裸燕麥種子,遇到短時間的低溫脅迫后,并不會喪失恢復能力,在適宜的溫度條件下仍能重新恢復生長,比延遲播種爭得了更寶貴的生長時間,為裸燕麥生產中播種期的確定提供了科學依據。
參考文獻:
[1]楊海鵬. 中國燕麥[M]. 北京:農業出版社,1989.
[2]鄭克寬,韓冰,于海峰,等. 裸燕麥新品種的經濟性狀和實用價值的研究[J]. 內蒙古農業大學學報:自然科學版,2002,23(1):61-65.
[3]崔海,王加啟,卜登攀,等. 燕麥飼料在動物生產中的應用[J]. 中國畜牧獸醫,2010,37(6):214-217.
[4]王雁麗,林鳳仙,趙英. 淺論高寒地區裸燕麥的開發及綜合利用[J]. 吉林農業科學,2007,32(3):57-59.
[5]趙海超,劉景輝,任永峰,等. 水分脅迫對內農大莜1號裸燕麥品種生長指標的影響[J]. 作物雜志,2010(1):73-76.
[6]李威. 六種裸燕麥品種苗期抗旱性比較[J]. 青海大學學報:自然科學版,2008,26(4):44-48.
[7]牛瑞明,王燕,吳桂麗,等. 裸燕麥種子萌發對模擬干旱脅迫的響應及其耐旱性綜合評價[J]. 麥類作物學報,2011,31(4):753-756.
[8]顏啟傳. 種子學[M]. 北京:中國農業出版社,2001:425-432.
[9]王學奎. 植物生理生化實驗原理和技術[M]. 2版.北京:高等教育出版社,2006:278-280.
[10]李合生. 植物生理生化實驗原理和技術[M]. 北京:高等教育出版社,2000.
[11]韓春梅. 不同溫度處理對豇豆種子萌發及抗氧化酶活性的影響[J]. 長江蔬菜,2010(10):29-30.
[12]呂優偉,賀佳圓,白小明,等. 9個野生早熟禾對低溫脅迫的生理響應及苗期抗寒性評價[J]. 草地學報,2014,22(2):326-333.
[13]李春燕,陳思思,徐雯,等. 苗期低溫脅迫對揚麥16葉片抗氧化酶和滲透調節物質的影響[J]. 作物學報,2011,37(12):2293-2298.
[14]張榮萍,吉瓊芳. 低溫脅迫對粳型巨胚稻西巨胚一號種子萌發的影響[J]. 西昌學院學報:自然科學版,2014,28(4):5-7.
[15]谷英楠,楊克軍,王玉鳳,等. 低溫對寒地玉米種子萌發及幼苗抗氧化酶活性的影響[J]. 安徽農業科學,2014,42(16):5092-5096.
[16]朱云集,崔金梅,彭文博,等. 不同萌發狀態的小麥種子在干旱逆境下的生理特性及恢復能力[J]. 華北農學報,1999,14(2):73-78.
[17]王建強,鄧永成. 低溫脅迫對銀葉菊和礬根抗寒生理指標的比較研究[J]. 中國農學通報,2014,30(7):224-227.
[18]曹慧明,史作民,周曉波,等. 植物對低溫環境的響應及其抗寒性研究綜述[J]. 中國農業氣象,2010,31(2):310-314, 319.
[19]吳惠俐,李雪平,高健,等. 禾本科植物抗寒機理的研究進展[J]. 竹子研究匯刊,2014,33(1):7-11.
[20]宋正熊,朱列書,尹佳,等. 低溫脅迫對煙草幼苗生化指標的影響及相關性分析[J]. 江西農業學報,2014,26(2):99-101, 105.
