西南石漠化地區2種巖生優勢樹種的光合生理
2016-07-25 00:16:35周文龍趙衛權蘇維詞張凡袁
江蘇農業科學 2016年6期
周文龍++趙衛權++蘇維詞++張凡++袁俊++易武英++謝元貴

摘要:利用光合儀測定了貴州石漠化治理區巖生優勢種青岡、火棘的光合生理生態特性,并比較它們的凈光合作用速率、蒸騰速率等生理生態指標的日變化特征。結果表明:青岡具有低光合速率、低蒸騰速率的特點,火棘具有高光合速率、高蒸騰速率的特點;青岡在石灰巖喀斯特石漠化區表現出的凈光合速率曲線較為平緩,未出現明顯的光合“午休”現象,火棘光合“午休”現象則相對明顯;青岡凈光合速率主要受氣孔性限制因素的影響,火棘凈光合速率主要受非氣孔性限制因素的影響。相關分析表明,在相關環境因子相關性顯著的情況下,環境因子對青岡的各項光合生理特性影響的顯著性不明顯,而火棘各項光合生理參數則受環境影響較大;植物自身的生理因子、環境生態因子相互作用,共同影響青岡、火棘的光合生理機能。
關鍵詞:貴州省;石漠化地區;巖生優勢樹種;青岡;火棘;光合生理
中圖分類號: S718.3文獻標志碼: A文章編號:1002-1302(2016)06-0477-03
收稿日期:2015-05-19
基金項目:貴州省科學技術基金(編號:黔科合J字[2012]2216號);貴州省軟科學項目(編號:黔科合R字[2014]2026號);貴州省社會發展攻關(編號:黔科合SY字[2012]3008號);貴州省社會發展攻關項目(編號:黔科合SY字[2014]3060號);貴州省科技創新人才團隊建設項目(編號:黔科合人才團隊[2014]4014號)。
作者簡介:周文龍(1984—),男,湖北鄂州人,碩士,助理研究員,研究方向為喀斯特環境資源與3S技術應用。E-mail:karstpro@hotmail.com。以貴州省為中心的西南喀斯特地區面積達55萬km2,是世界上喀斯特分布面積最大、發育最強的區域[1]。峰叢洼地是西南喀斯特山區主要的地貌形態,峰叢洼地區內碳酸鹽巖成片分布[2],是受石漠化威脅最嚴重的一類巖溶地貌[3]。喀斯特植被的恢復與重建是峰叢洼地生態恢復、石漠化防治的核心內容之一[4]。青岡(Cyclobalanopsis glauca)、火棘(Pyracantha fortuneana)是喀斯特原始頂極植被群落受到破壞后的主要遺留物種,同時也是石漠化地區植被恢復的主要樹種,具有很強的水土保持能力。光合作用決定了植物能量吸收、有機物積累的數量,也是其他生理過程、生命活動的基礎,并與植物生長、發育、存活密切相關,是植物生產力的最主要因素[5-6]。葉片光合特性與植物所處的環境密切相關,其對不同的生境條件會表現出不同的適應特性與適應機制[7-10]。目前對喀斯特山區的青岡光合生理研究多集中在廣西白云巖喀斯特石漠化地區[11-15],而對貴州石灰巖喀斯特石漠化的青岡光合生理研究則很少,關于火棘在喀斯特地區光合生理的研究更鮮有報道[16-17]。為了進一步了解青岡、火棘在貴州喀斯特石漠化地區的光合生理表現,本研究探討青岡、火棘的葉片凈光合速率等主要光合生理指標,以期為喀斯特石漠化環境恢復、植被演替研究提供一定的理論依據。
1材料與方法
1.1試驗材料與研究區自然概況
試驗材料為示范區朝營核心小流域內“大山”山頂退耕地邊緣自然生長的青岡和火棘。
本試驗在畢節撒拉溪國家“十二五”石漠化綜合治理示范區進行,位于貴州省畢節市七星關區撒拉溪鎮境內(105°02′01″~105°08′09″E,27°11′36″~27°16′51″N),屬于烏江上游段六沖河流域,平均海拔約1 800 m,屬亞熱帶季風濕潤氣候,年均氣溫12 ℃,無霜期245 d,年均日照時間為1 360 h,年均降水量984.4 mm,80%以上的降水集中在6—9月。研究區屬于典型的亞熱帶喀斯特高原山地(峰叢洼地)輕-中度喀斯特石漠化類型,主要土壤類型為黃壤,原始植被以青岡、火棘、杜鵑(Rhododendron simsii)等藤刺灌叢為主,局部山坡、居民地四周零星分布有云南松(Pinus yunnanensis)[18]。
1.2試驗方法
2014年4月下旬,在晴天、自然條件下進行光合試驗,在8:00—18:00時段內,取中間時段每隔2 h測定1次,共測6次,青岡、火棘各隨機選取3株,每株選取葉位相同的3張中上部成熟向陽葉片進行試驗,取平均值計算。
光合測定儀器使用英國ADC Bioscientific公司生產的Lcpro+便攜式光合測定儀,使用紅藍光源葉室測定葉片瞬時光合速率等指標。主要測定、統計指標包括:凈光合速率[Pn,μmol/(m2·s),以CO2計]、蒸騰速率[Tr,mmol/(m2·s),以H2O計]、CO2氣孔導度[Gs,mol/(m2·s),以H2O計]、細胞間CO2濃度(Ci,μmol/mol)、葉室溫度(Tch, ℃)、葉片溫度(Tleaf,℃)、光合有效輻射[PAR,μmol/(m2·s)]等。水分利用效率WUE(μmol/mmol)的計算公式:
WUE=Pn/Tr。
采用SPSS 20.0進行數據分析,用Excel 2007進行數據統計與制圖。
2結果與分析
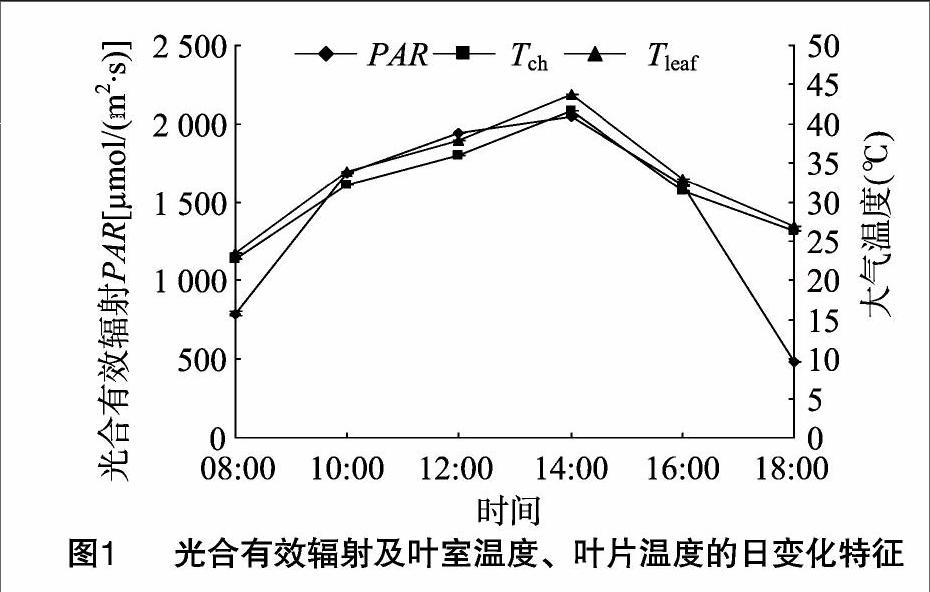
2.1光合有效輻射等環境因子的日變化特征
在08:00—18:00期間,PAR的變化范圍為481.25~2 049.08 μmol/(m2·s),最高值出現在14:00左右,最低值出現在18:00,受地形影響,08:00山頂太陽光合輻射較強。08:00—18:00期間,Tch的變化范圍為22.75~41.75 ℃,08:00 左右Tch最低,最高值出現在14:00左右,18:00下降至2630 ℃(圖1);與Tch相比,由于葉片受太陽光合輻射影響,Tleaf要高出Tch 2.05~0.50 ℃。此外,大氣壓強日變化為8.14萬~817萬Pa,受中午上升氣流影響,中午大氣壓強要高于早、晚大氣壓強(大氣壓強數據僅說明,未在圖表中體現)。
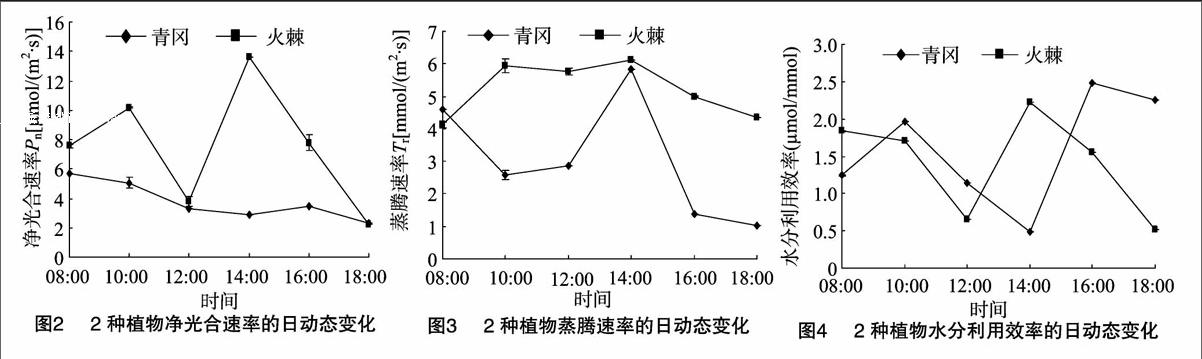
2.2凈光合速率的日變化特征
由圖2可見:青岡、火棘的凈光合速率日變化曲線存在較大差異,青岡凈光合速率的日均值僅為火棘的50%左右(表1);從峰型上看,火棘為典型的“雙峰”型日變化曲線,青岡的日變化曲線呈現逐漸下降的趨勢,下降期間(14:00—16:00)也存在1個不明顯的上升階段。青岡凈光合速率曲線日變化幅度較為平緩,變化差異遠小于火棘,在08:00—14:00凈光合速率一直呈下降趨勢,在14:00出現不明顯的光合谷值;在14:00—16:00略有上升,光合“午休”現象不明顯。火棘的凈光合速率從8:00開始快速增長,在10:00達到第1個峰值(次峰值),12:00出現光合谷值,在14:00形成第2個峰值(最大值),光合“午休”現象持續時間較短。
2.3蒸騰速率的日變化特征
由圖3可見:青岡、火棘的蒸騰速率日變化差異較大,青岡日變化幅度要高于火棘,火棘則相對平緩。青岡在08:00出現次峰值,在10:00出現蒸騰谷值,在14:00出現最大值,此后曲線陡然下降。火棘的蒸騰速率日變曲線相對青岡而言變化較為平緩,分別在10:00、14:00出現了2個峰值,在12:00 出現了不明顯的谷值。整體而言,青岡的蒸騰速率除在 08:00 外均低于火棘,且除14:00之外,均明顯低于火棘的蒸騰速率。
2.4水分利用效率的日變化特征
水分利用效率的高低是植物適應環境能力強弱的重要指標[20]。由圖4可以看出:青岡、火棘的水分利用效率日變化曲線變化趨勢較為接近,青岡水分利用效率曲線較火棘有所平移滯后;青岡水分利用效率在10:00(次峰值)、16:00(最高值)出現2個峰值,在14:00出現了1個非常明顯的谷值,在10:00—16:00之間變化差異較大,14:00、16:00之間的水分利用效率差異較大;火棘水分利用效率在14:00(最大值)出現峰值,12:00出現谷值,在14:00之后又逐漸下降。
2.5氣孔導度的日變化特征
由圖5可以看出:從日變化特征上看,青岡的氣孔導度曲線呈“L”形,火棘呈“U”形,且在14:00之前二者變化幅度較為接近,之后差異漸大;青岡、火棘的氣孔導度日均值分別為(0.13±0.20)、(0.20±0.14) mol/(m2·s),火棘氣孔導度日均值略高于青岡。
2.6胞間CO2濃度的日變化特征
由圖6可以看出:青岡、火棘的胞間CO2濃度曲線變化較為接近,在10:00前青岡的胞間CO2濃度高于火棘,之后則相反;青岡胞間CO2濃度最高值出現在8:00,最低值出現在16:00,火棘最高值出現在18:00,最低值出現在10:00。
2.7環境因子與光合特征參數間的相關性
由表2可知,青岡的Pn與PAR、Tch、Tleaf、Tr、Ci、Gs等各因子相關性均不顯著,部分環境因子之間呈顯著或極顯著相關;火棘的Pn與PAR、Tch、Tleaf、Tr、Ci、Gs等各因子的相關性均不顯著,部分環境因子之間呈顯著或極顯著相關。例如,火棘的環境參數PAR與Gs顯著相關,與Tr極顯著相關,Tch、Tleaf與Gs、Tr呈極顯著相關,Gs與Tr呈極顯著相關。
3結論與討論
研究青岡、火棘的光合生理特點及變化規律對于研究巖石樹種在石漠化地區的水土保持效益以及栽培技術措施十分必要。研究結果表明:火棘凈光合速率日變化呈“雙峰”型曲線,正午存在明顯的光合“午休”現象;青岡在本試驗中未表現明顯的光合“午休”現象,與廣西白云巖喀斯特區域日變化曲線存在一定差異[11-13,15],但廣西同一地區的不同研究也存在一定差異[14]。上述研究表明,在不同區域、不同類型喀斯特地區的青岡凈光合速率日變化特征表現存在差異。
從試驗結果可知,在一定范圍內,青岡葉片內細胞間CO2濃度越高,其供給光合所需的原料越多,凈光合速率越高,光合速率會超過呼吸速率;氣孔性限制因素是由于氣孔導度降低阻止了光合原料CO2的供應;非氣孔性限制因素是由于植物葉片細胞光合能力的下降使葉肉細胞利用CO2的能力降低,從而造成細胞間CO2含量升高。對青岡、火棘的研究表明,青岡凈光合速率主要受氣孔性限制因素的影響,火棘凈光合速率主要受非氣孔性限制因素的影響。對于非氣孔性限制光合速率的情況,在晴天中午可以采取一定的遮光降溫措施,或適時灌溉,尤其是噴灌,有利于降低火棘溫度、增加空氣相對濕度,減少水分蒸騰[19-20],是提高火棘葉片光合水平的重要措施。
相關分析表明,在相關環境因子相關性顯著的情況下,環境因子對青岡的各項光合生理特性影響的顯著性不明顯;而火棘各項光合生理參數則受環境影響較大。植物自身的生理因子與環境生態因子相互作用,共同影響青岡、火棘的光合生理功能。喀斯特的特殊地域環境、生態因子限制了石漠化地區的植被恢復。本研究僅針對貴州喀斯特石漠化治理區最主要的巖石植被群落優勢種進行觀測,關于其他石漠化區中巖石物種的光合生理特征仍然需要進一步研究。在喀斯特環境,特別是受損的喀斯特環境中,植被會產生特殊的適應機理,因此對石漠化治理中的植被群落的人工誘導演替恢復,不僅要從植被群落內部生理機制出發,還要結合外界生態環境的變化特點,因地制宜地選取適宜物種和群落組建模式。
參考文獻:
[1]熊康寧,池永寬. 中國南方喀斯特生態系統面臨的問題及對策[J]. 生態經濟,2015,31(1):23-30.
[2]曹建華. 中國地質科學院巖溶地質研究所,國土資源部巖溶動力學重點實驗室.受地質條件制約的中國西南巖溶生態系統[M]. 北京:地質出版社,2005:10-20.
[3]李先琨,何成新,蔣忠誠. 巖溶脆弱生態區生態恢復、重建的原理與方法[J]. 中國巖溶,2003,22(1):12-17.
[4]龐冬輝,沈利娜,蔣忠誠,等. 廣西弄拉峰叢洼地灌叢巖生優勢種光合生理生態特征日變化研究[J]. 中國巖溶,2010,29(3):293-300.
[5]柯世省. 珍稀植物香果樹光合作用日進程初步研究[J]. 福建林業科技,2009,36(2):226-230.
[6]李慶康,馬克平. 植物群落演替過程中植物生理生態學特性及其主要環境因子的變化[J]. 植物生態學報,2002,26(z1):9-19.
[7]Manes F,Seufert G,Vitale M. Ecophysiological studies of Mediterranean plant species at the Castelporziano estate[J]. Atmospheric Environment,1997,31(Suppl 1):51-60.
[8]Ueda Y,Nishihara S,Tomita H,et al. Photosynthetic response of Japanese rose species Rosa bracteata and Rosa rugosato to temperature and light[J]. Scientia Horticulturae,2000,84(3/4):365-371.
[9]Midgley G F,Aranibar J N,Mantlana K B,et al. Photosynthetic and gas exchange characteristics of dominant woody plants on a moisture gradient in an African savanna[J]. Global Change Biology,2004,10(3):309-317.
[10]John R,Dalling J W,Harms K E,et al. Soil nutrients influence spatial distributions of tropical tree species[J]. Proceedings of the National Academy of Sciences of the United States of America,2007,104(3):864-869.
[11]張中峰,黃玉清,莫凌,等. 桂林巖溶區青岡櫟光合速率與環境因子關系初步研究[J]. 廣西植物,2008,28(4):478-482.
[12]張中峰,尤業明,黃玉清,等. 模擬巖溶水分供應分層的干旱脅迫對青岡櫟光合特性和生長的影響[J]. 生態學雜志,2012,31(9):2197-2202.
[13]沈利娜,蔣忠誠,吳孔運,等. 峰叢洼地恢復演替系列優勢種光合生理生態特征日變化研究——以廣西馬山弄拉峰叢洼地為例[J]. 廣西植物,2010,30(1):75-81,35.
[14]莫凌,黃玉清,覃家科,等. 巖溶植物青岡和九龍藤葉片光學特性日變化研究[J]. 廣西科學,2008,15(2):189-194.
[15]張中峰,張金池,黃玉清,等. 接種叢枝菌根真菌對青岡櫟幼苗生長和光合作用的影響[J]. 廣西植物,2013,33(3):319-323,294.
[16]劉長成,劉玉國,郭柯. 四種不同生活型植物幼苗對喀斯特生境干旱的生理生態適應性[J]. 植物生態學報,2011,35(10):1070-1082.
[17]張俊佩. 貴州石漠化地區主要造林樹種耐旱特性及適應性評價[D]. 北京:中國林業科學研究院,2009.
[18]池永寬,熊康寧,張錦華,等. 喀斯特石漠化地區三種豆科牧草光合與蒸騰特性的研究[J]. 中國草地學報,2014,36(4):116-120.
[19]池永寬,熊康寧,王元素,等. 西南石漠化地區兩種經濟林木光合日動態特征[J]. 經濟林研究,2015,33(1):45-49,55.
[20]杜偉,程嘉翎,柴建萍,等. 云南鶴慶桑樹光合生理特性日變化研究[J]. 西南農業學報,2012,25(4):1227-1231
