槲皮素-AAPH氧化產物結構鑒定及其對HepG2細胞氧化還原狀態的影響
2016-08-11 11:32:57溫斐婷成向榮陳侃俊樂國偉施用暉
安徽農業科學 2016年17期
關鍵詞:氧化應激
溫斐婷,成向榮,陳侃俊,王 菲,樂國偉,施用暉
(江南大學食品學院,江蘇無錫 214122)
?
槲皮素-AAPH氧化產物結構鑒定及其對HepG2細胞氧化還原狀態的影響
溫斐婷,成向榮,陳侃俊,王 菲,樂國偉*,施用暉
(江南大學食品學院,江蘇無錫 214122)
[目的]探究AAPH誘導的槲皮素氧化產物對HepG2細胞氧化還原狀態的影響。[方法]用紫外-可見波長掃描、HPLC-MS方法分析AAPH和槲皮素在37 ℃下反應12 h生成的產物結構;以槲皮素-AAPH反應產物處理HepG2細胞24 h,測定其對細胞存活率、ROS水平及細胞總抗氧化能力(T-AOC)、谷胱甘肽過氧化物酶(GSH-Px)活性、超氧化物歧化酶(SOD)活性、GSH/GSSG比值及丙二醛(MDA)含量的影響,并測定Nrf2、Keap1、NQO1、Prdx1相關抗氧化基因的mRNA表達。[結果] AAPH與槲皮素體系在反應12 h后于294 nm波長處有特征吸收,LC-MS解析結果表明該體系反應生成了含4種主要氧化產物的混合物。與空白組相比,槲皮素與AAPH氧化產物孵育顯著降低了細胞存活率,同時細胞內ROS水平和MDA含量升高,細胞內T-AOC水平、GSH-Px及SOD活性顯著降低,并顯著下調Nrf2、NQO1、Prdx1 mRNA表達,上調Keap1 mRNA表達。[結論]槲皮素氧化產物可造成HepG2細胞出現氧化應激。
槲皮素;氧化產;HepG2細胞;氧化應激
以槲皮素為代表的黃酮類物質是重要的植物化合物,其膳食攝入對人體心腦血管保護具有特殊意義[1]。但隨著研究的深入,已有不少體外評價實驗和動物試驗發現某些黃酮類化合物在高濃度、長時間作用時表現出生物毒性,且其毒性與促氧化作用密不可分[2-3]。黃酮類化合物的還原形式主要起抗氧化作用,但其氧化形式半苯醌、苯醌和甲基醌有很強的促氧化作用,會導致谷胱甘肽(GSH)減少,與尼克酰胺腺嘌呤二核苷酸磷酸[NAD(P)H]和其他細胞還原劑的協同氧化,并伴隨活性氧(ROS)的產生[4-5]。
有研究指出,槲皮素清除自由基的中間產物具有生物毒性,可能是導致高濃度槲皮素促氧化現象出現的原因之一。在清除自由基的過程中,槲皮素吸收1或2個·H形成半苯醌和苯醌,而半苯醌和苯醌的活性形式可直接誘導DNA氧化損傷[6]。此外,高濃度槲皮素(>0.2 mmol/L)會促進O2-·和H2O2的產生,導致活性酶SH-基團的氧化[7]。但目前直接針對槲皮素清除自由基過程中生成的氧化產物進行細胞毒性研究的報道較少。2,2-偶氮二(2-甲基丙基咪)二鹽酸鹽(AAPH)是常用的ROS誘導劑,能夠在機體內持續穩定地促進ROS 生成[8]。筆者以AAPH作為氧化劑,模擬了槲皮素的自由基清除過程,并初步鑒定了兩者在化學體系中的反應產物結構,探究了該產物對HepG2細胞氧化還原狀態的影響,旨在為探討高濃度槲皮素促氧化傾向的潛在機制提供理論依據。
1 材料與方法
1.1材料
1.1.1試劑。人肝癌細胞株HepG2,中國科學院上海生物化學與細胞生物學研究所;槲皮素,美國Sigma公司;AAPH,上海阿拉丁生物科技公司;DMEM培養基,美國Gibco公司;小牛血清,浙江省杭州四季青生物材料工程研究所;青霉素、鏈霉素、胰蛋白酶、二甲基亞砜(DMSO)、噻唑藍(MTT)、2’,7’-二氯熒光素二乙酸酯(DCFH-DA)熒光探針,美國Sigma公司產品;CAT、SOD、MDA、T-AOC、GSH-PX測定試劑盒,南京建成生物工程研究所;BCA蛋白濃度測定試劑盒、Trizol試劑盒,上海生工生物工程公司;M-MLV逆轉錄酶,Promega公司;基因引物,上海捷瑞生物工程有限公司。
1.1.2儀器。UV2300雙光束紫外可見分光光度計,上海天美科學儀器有限公司;高效液相色譜(HPLC)儀、質譜(MS)儀,美國Waters公司;二氧化碳恒溫細胞培養箱,Thermo公司;7900 HT Fast Real-Time PCR儀,美國ABI公司;5804R臺式高速冷凍離心機,德國Eppendorf公司。
1.2方法
1.2.1槲皮素-AAPH氧化產物制備及紫外掃描。取槲皮素100 μmol/L與AAPH 600 μmol/L于37 ℃水浴反應,分別于0、3、6、9、12 h取樣,進行紫外-可見光全波長掃描(200~600 nm),確定槲皮素氧化產物的特征吸收波長。
按Q∶AAPH=1∶6的濃度比,取槲皮素5 mmol/L與AAPH 30 mmol/L于37 ℃水浴,分別于0、6、9、12 h取樣進行高效液相色譜測定。
1.2.2高效液相色譜條件及質譜條件。高效液相色譜儀,Waters Acquity UPLC;檢測器:Waters Acquity PDA;色譜柱:BEH-C18(250.0 mm×4.6 mm);流動相A:甲醇;流動相B:0.1%甲酸;流動相的等度洗脫條件為:45%B+55%A(20 min);檢測波長為200~600 nm;分析波長為294 nm;進樣量為10 μL;流速0.3 mL/min;柱溫保持在37 ℃。
質譜離子方式為負離子;毛細管電壓:3.0 kV;錐孔電壓:20 V;離子源溫度:100 ℃;溶劑溫度:250 ℃;脫溶劑氣體流速:500 L/h。錐孔氣體流速:50 L/h,碰撞能量:30 eV,質量范圍:50~1 500 m/z;檢測器電壓:1 700 V。
1.2.3細胞培養。HepG2細胞培養于含有10%小牛血清、100 U/L青霉素、100 g/mL鏈霉素的DMEM培養基中,于37 ℃、體積分數為5%的CO2飽和濕度培養箱中常規培養。待細胞長至80%融合時,棄去舊培養基,加入3 mL 0.25%胰蛋白酶消化液消化,按1×106個/板傳入培養皿。
1.2.4試驗分組及處理。槲皮素5 mmol/L與AAPH 30 mmol/L于37 ℃水浴反應9 h,制備槲皮素-AAPH氧化產物。細胞貼壁后分別采用不同條件處理,試驗分組如下:正常對照組(CON),槲皮素氧化產物處理組0.1 μmol/L(QO0.1)、1.0 μmol/L(QO1)、10.0 μmol/L(QO10)、50.0 μmol/L(QO50)、100.0 μmol/L(QO100)、200.0 μmol/L(QO200)、300.0 μmol/L(QO300)。
1.2.5MTT法測定細胞存活率。HepG2細胞以8 000個/孔接種于96孔板,貼壁24 h后,換無血清培養基,分別加入0.1、1.0、10.0、50.0、100.0、200.0、300.0 μmol/L槲皮素氧化產物,繼續培養6、12、24、48 h后,加入0.5 mg/mL MTT,孵育4 h。以150 μL DMSO溶解沉淀,在525 nm波長下測定吸光度。
存活率=試驗組OD值/正常對照組OD值×100%
1.2.6DCFH-DA法測定活性氧自由基(ROS)。細胞分別加入0.1、1.0、10.0、50.0、100.0 μmol/L槲皮素氧化產物,繼續培養12、24、48 h后,每孔加入100 μL終濃度為10.0 μmol/L的DCFH-DA 探針,避光孵育30 min,M5熒光酶標儀檢測細胞熒光強度(激發波長488 nm,發射波長525 nm)。
ROS水平=(試驗組熒光值/試驗組細胞存活率)/(正常對照組熒光值/正常對照組細胞存活率)
1.2.7抗氧化酶活性測定。HepG2細胞以1.7×105個/孔濃度接種于6孔板中,分組及處理同“1.2.6”,孵育結束后加RPMI細胞裂解液150 μL于冰上裂解細胞,4 ℃、10 000 r/min離心10 min,收集細胞上清液。采用BCA法測定蛋白濃度。細胞內T-AOC、GSH-Px、SOD及MDA含量均按照試劑盒說明書測定。
1.2.8qRT-PCR檢測基因mRNA表達。Trizol法提取細胞總RNA,反轉錄為cDNA,-20 ℃保存。qRT-PCR測定相關基因的mRNA水平。PCR擴增條件:95 ℃預變性 5 min;95 ℃變性20 s,62 ℃退火30 s,72 ℃延伸20 s,72 ℃終末延伸2 min,循環40次。基因引物序列見表1。

表1 基因的引物序列
2 結果與分析
2.1槲皮素-AAPH氧化產物結構鑒定槲皮素100.0 μmol/L與600.0 μmol/L AAPH于37 ℃水浴反應,溶液經過紫外-可見分光光度計全波長掃描后所得圖譜如圖1所示,隨著反應時間增加,槲皮素在254和374 nm處的特征吸收峰吸光值持續下降,而294 nm處的吸收持續上升,在12 h后可看到明顯的吸收峰,表明新生成的槲皮素氧化產物紫外特征吸收波長為294 nm。
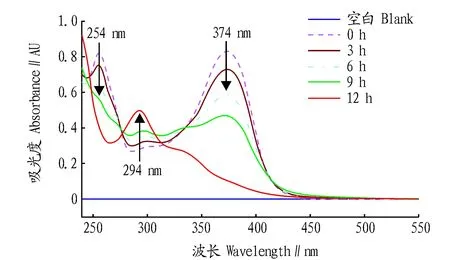
圖1 槲皮素氧化產物紫外-可見全波長掃描圖譜Fig.1 UV-vis spectra of quercetin oxidation
由圖2、3可見,隨著反應時間增加,在2.07、2.67、3.97、6.57 min 4個時間點峰面積都增加,表明這幾個時間點是槲皮素氧化產物的特征出峰時間。選取高效液相色譜圖譜中4個特征峰進行物質分析,結果如圖4所示,2.07和3.97 min的物質為槲皮素被氧化后,在A環發生裂解后B環和C環分別加氫生成的產物,分子量分別為154和170。2.67 min的物質為槲皮素A環的雙鍵被氧化,分別加合上甲氧基和氧的產物,分子量為350。6.57 min的物質為槲皮素B環上的2個羥基脫掉2個氫生成的鄰醌,分子量為300。

圖2 槲皮素-AAPH反應不同時間產物的高效液相圖譜Fig.2 HPLC chromatogram on each reaction time of quercetin oxidation products

注:*表示兩組間存在顯著差異。Note:*stands for significant difference between two groups.圖3 槲皮素-AAPH氧化產物液相圖不同保留時間的峰面積變化Fig.3 Changes of peak areas on each retention time in HPLC chromatogram of quercetin oxidation products
2.2槲皮素氧化產物對細胞存活率及ROS水平的影響正常生理狀況下,細胞內ROS生成和清除處于動態平衡,但是一旦抗氧化功能降低或者ROS過多都能引起細胞氧化應激使細胞受到損傷。用不同濃度的槲皮素氧化產物分別對HepG2細胞進行孵育,孵育6 h后,與對照組相比,低濃度的槲皮素氧化產物組細胞存活率未發生顯著變化,但氧化產物200.0和300.0 μmol/L濃度下細胞存活率顯著下降,且時間越長,下降越顯著;孵育24 h后,300.0 μmol/L濃度組細胞存活率降至70.34%。隨著孵育時間的增加,較低濃度的氧化產物也使細胞存活率顯著降低;孵育48 h后,0.1 μmol/L濃度下存活率下降顯著,半數致死劑量為300.0 μmol/L。0.1、1.0 μmol/L的低濃度槲皮素氧化產物孵育12 h,細胞ROS水平與空白組相比無顯著差異,但隨著槲皮素氧化產物濃度和作用時間的增加,細胞ROS水平極顯著增加(圖5)。說明高濃度的槲皮素氧化產物能使細胞在短時間內出現較大死亡,而低濃度在長時間孵育下也能夠造成細胞一定程度的死亡。
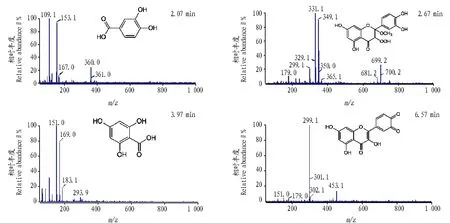
圖4 槲皮素氧化產物特征峰的 MS 分析Fig.4 MS spectrum of fractions at different retention time from HPLC

圖5 槲皮素氧化產物對HepG2細胞存活率及ROS水平的影響Fig.5 The influence of quercetin oxidation products to HepG2 cell viability and the ROS
2.3槲皮素氧化產物對細胞氧化還原狀態的影響SOD、GSH-Px是細胞受到外界刺激產生的內源性的抗氧化酶類,清除胞內活性氧物質,對維持氧化與抗氧化平衡起著重要作用,其活性可間接反映細胞內氧化應激水平[9-10]。T-AOC能全面衡量機體防御體系氧化還原狀態。MDA是細胞膜脂質被活性氧氧化后的產物,是評價細胞氧化損傷程度的重要標志物,其升高可直接反映受到的氧化應激損傷。
由圖6可知,槲皮素氧化產物孵育細胞12 h后,0.1、1.0、10.0 μmol/L低、中濃度組中細胞T-AOC水平與SOD活力相較CK組無顯著變化,100.0 μmol/L濃度組有輕微下降。但隨孵育時間延長,T-AOC水平出現顯著下降,連續孵育48 h后,低濃度組細胞內SOD活力也顯著下降。與CK組相比,50.0、100.0 μmol/L高濃度組中細胞T-AOC水平在12 h孵育后即出現顯著下降。

注:*表示P<0.05;**表示P<0.01;***表示P<0.001。Note:* stands for P<0.05;** stands for P<0.01; *** stands for P<0.001.圖6 槲皮素氧化產物對HepG2細胞氧化還原狀態的影響Fig.6 The effect of quercetin oxidation products on antioxidant index of HepG2 cell
0.1、1.0、10.0 μmol/L低、中濃度的槲皮素氧化產物孵育12 h,細胞內GSH-Px活力較CK組顯著增加;但隨孵育時間延長,酶活力下降,24 h時10.0 μmol/L組的GSH-Px活力已低于CK組水平;當孵育48 h時,細胞內GSH-Px活性出現急劇下降。和低濃度組不同,100.0 μmol/L高濃度組中細胞GSH-Px活力則是在12 h孵育后即出現顯著下降。產生這種現象的原因可能是由于低濃度的槲皮素氧化產物在短時間孵育的條件下并未對細胞造成較大影響,細胞內少量的ROS導致GSH-Px活性出現應激性的上調。但在孵育48 h后,ROS產生量超過了細胞的清除能力,GSH-Px活性受到抑制,10.0 μmol/L濃度組酶活出現急劇下降。
細胞內MDA含量則隨著加入的槲皮素氧化產物濃度的升高而升高,并與孵育時間成正比,提示槲皮素氧化產物濃度的升高和孵育時間的增長均造成了細胞內脂肪氧化加劇,進一步說明細胞內抗氧化機制已經不能清除ROS,細胞內形成了氧化應激。
2.4槲皮素及其氧化產物對細胞氧化還原相關mRNA表達的影響ROS能夠損傷細胞內大分子物質并破壞其生理功能,為了緩解細胞所遭受的損傷,細胞內存在一整套復雜的氧化應激應答系統。轉錄調節因子核細胞系因子2相關因子(Nrf2)是細胞抵抗氧化應激反應的關鍵基因,與Keap1作用,轉錄翻譯產生一些抗氧化物,如SOD、血紅素氧合酶(HO-1)、NADPH苯醌氧化還原酶1(NQO-1)等II相解毒酶,以及過氧化還原酶(Prdx)、金屬硫蛋白(MT-1)等抗氧化酶,從而對抗ROS引發的氧化應激,保護正常細胞不受破壞[11-13]。該研究對 Nrf2 以及其部分下游靶基因mRNA表達。
由圖7可知,在與槲皮素氧化產物共同孵育24 h后,1.0與10.0 μmol/L低濃度組細胞中Nrf2基因表達出現顯著上調,而50.0與100.0 μmol/L高濃度組細胞中Nrf2出現顯著下調,說明在長時間作用下,低濃度槲皮素氧化產物造成細胞出現氧化應激,Nrf2代償性表達上調,但高濃度槲皮素氧化產物則直接抑制了Nrf2的表達。而細胞中Keap1基因表達均由于槲皮素氧化產物的加入而顯著上調,并隨著其濃度的升高上調越顯著。
NADPH苯醌氧化還原酶1(NQO-1)[11]可借助NADP催化體內醌類化合物發生還原反應。由圖7可知,低濃度的槲皮素氧化產物可以顯著上調NQO-1的表達,而50.0 μmol/L的氧化產物對NQO-1的影響與CK組無顯著差異,100.0 μmol/L組則顯著下調NQO-1的表達。過氧化物還原酶(Prdx)在細胞中的氧化還原狀態調節中起著重要作用。過氧化物還原酶是通過硫氧還蛋白系統清除細胞內新陳代謝所產生的過氧化物[14]。圖7顯示,槲皮素氧化產物低濃度組均造成了細胞內Prdx表達的顯著升高,提示該試驗條件下造成了細胞出現氧化應激。而高濃度的槲皮素氧化產物則對細胞內Prdx基因表達出現下調。
3 結論
通過紫外-可見分光光譜和LC-MS法初步鑒定了槲皮素-AAPH氧化產物的結構,并測定了槲皮素氧化產物對HepG2細胞存活率和自由基水平的影響,以及氧化還原狀態、抗氧化通路相關基因的測定。結果表明,槲皮素氧化產物會加劇細胞的氧化應激,具有一定的細胞毒性,高濃度的槲皮素有可能是通過其在清除自由基過程中形成的大量氧化產物而造成促氧化現象的出現。

注:*表示P<0.05;**表示P<0.01;***表示P<0.001。Note:* stands for P<0.05;** stands for P<0.01; *** stands for P<0.001.圖7 槲皮素氧化產物對HepG2細胞Nrf2、Keap-1、NQO-1及Prdx表達的影響Fig.7 The influence of quercetin oxidation products to Nrf2,Keap-1,NQO-1 and Prdx expression of HepG2 cell
[1] 陸柏益,張英,吳曉琴.黃酮類化合物的潛在毒性作用[J].中國中藥雜志,2006,31(7):533-536.
[2] 湯立建,趙良才,李慶林,等.黃芩黃酮類成分抗腫瘤作用及機制研究進展[J].中國中藥雜志,2007,32(1):21-26.
[3] 汪何雅,趙顯峰,王茵,等.黃酮類化合物的潛在毒性[J].衛生研究,2007,36(5):640-642.
[4] NEMEIKAITE-CENIENE A,IMBRASAITE A.Quantitative struc-ture-activity relationships in prooxidant cytotoxicity of poly-phenols:Role of potential of phenoxyl radical/phenol redoxcouple[J].Arch Biochem Biophys,2005,441(2):182-190.
[5] FUKUMOTO L R,MAZZA G.Assessing antioxidant and prooxi-dant activities of phenolic compunds[J].J Agric Food Chem,2000,48(8):3597-3604.
[6] FIORUCCI S,GOLEBIOWSKI J,CABROL-BASS D,et al.DFT studyof quercetin activated forms involved in antiradical,antioxi-dant,and prooxidant biological processes[J].J Agric Food Chem,2007,55(3):903-911.
[7] SCHMALHAUSEN E V,ZHLOBEK E B,SHALOVA I N,et al.An-tioxidant and prooxidant effects of quercetin on glyceralde-hyde-3-phosphate dehydrogenase[J].Food Chem Toxicol,2007,45(10):1988-1993.
[8] BELLOMO G,VAIRETTI M,STIVALA L,et al.Demonstrationof nuclear ompartmentalization of glutathione inhepatocytes[J].Proceedings of national academy of science USA,1992,89:4412-4416.
[9] SUGINO N.The role of oxygen radical-mediated signaling pathway sinendome trial function[J].Placenta,2007,28:133-136.
[10] ANDRIANTSITOHAINA R,DULUC L,GARCIA-RODRIGUEZ J C,et al.Systems biology of antioxidants[J].Clinical science,2012,123(3):173-192.
[11] KOBAYASHI A,OHTA T,YAMAMOTO M.Unique function of the Nrf2-Keap1 pathway in the inducible expression of antioxidant and detoxifying enzymes[J].Methods in enzymology,2003,378:273-286.
[12] ALEKSUNES L M,KNIGHT T R,KLAASSEN C D,et al.Nuclear factor-e2-related factor 2 expression in liver is critical for induction of nad(p)h:Quinone oxidoreductase 1 during cholestasis[J].Cell stress & chaperones,2006,11(4):356-363.
[13] KOBAYASHI M,YAMAMOTO M.Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species[J].Adv Enzyme Regul,2006,46:113-140.
[14] NEUMANN C A,KRAUSE D S,CARMAN C V,et al.Essential role for the peroxiredoxin Prdx1 in erythrocyte antioxidant defence and tumour suppression[J].Nature,2003,424(6948):561-565.
Structure Identification of the Quercetin Oxidation Products Induced by AAPH and Effects on the Redox State of HepG2 Cells
WEN Fei-ting, CHENG Xiang-rong, CHEN Kan-jun, LE Guo-wei*et al
(School of Food Science and Technology, Jiangnan University, Wuxi, Jiangsu 214122)
[Objective] The aim was to explore how the quercetin oxidation products induced by AAPH influence the redox state of HepG2 cells. [Method] Quercetin was mixed with APPH as oxidant under 37 ℃ for 12 h. The structural characterization were analyzed by UV scan and LC-MS. HepG2 cells were treated with reaction product of quercetin-AAPH and incubated for 24 h. Viable cells were harvested, and the cell viability, ROS level, glutathione (GSH) and glutathione disulphide (GSSG) content, malandialdehyde (MDA) level, total antioxidant capacity (T-AOC), superoxide dismutase (SOD) and catalase (CAT) activity were assayed with the appropriate test kits, the oxidant stress related genes were assayed by DNA microarray. [Result] The chromatogram showed peaks eluting at 294 nm after 12 h reaction and the structural analysis result indicated that mixture of four oxidation products were obtained in the reaction system. Compared with the blank group, Quercetin-AAPH oxidation products elevated the ROS and MDA generation, and reduced cell viability, T-AOC, CAT, GSH-Px and SOD and GSH/GSSG activity significantly. Nrf2, NQO-1, Prdx expression level was down-regulated and Keap-1, Nox-2, Cox-2 expression level was up-regulated in the meanwhile. [Conclusion] These findings indicate that quercetin oxidation products could prompt HepG2 cells to generate more oxidative stress.
Quercetin; Oxidation products; HepG2 cells; Oxidative stress
溫斐婷(1990- ),女,廣西玉林人,碩士研究生,研究方向:營養與功能因子。*通訊作者,教授,博士,從事營養與功能因子研究。
2016-04-26
Q 2
A
0517-6611(2016)17-034-05
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
