‘蘭箭3號’箭筈豌豆莢果發育動態及腹縫線結構研究
2016-09-02 02:41:35董德珂王彥榮劉志鵬
西北植物學報 2016年7期
董德珂,董 瑞,王彥榮,聶 斌,劉志鵬
(蘭州大學 草地農業科技學院草地農業生態系統國家重點實驗室,蘭州 730020)
?
‘蘭箭3號’箭筈豌豆莢果發育動態及腹縫線結構研究
董德珂,董瑞,王彥榮,聶斌,劉志鵬*
(蘭州大學 草地農業科技學院草地農業生態系統國家重點實驗室,蘭州 730020)
箭筈豌豆(Viciasativa)是高海拔地區重要的一年生豆科牧草,但莢果成熟時的開裂現象會造成種子的嚴重損失。該研究以栽培品種‘蘭箭3號’為對象,對其莢果在發育過程中的形態特征、水分含量、腹縫線表面結構及腹縫線橫截面解剖結構的動態變化進行觀察分析,以探討箭筈豌豆莢果的裂莢機理,為生產中確定種子收獲的適宜時間提供理論依據。結果顯示:(1)‘蘭箭3號’約在盛花后25~30 d莢果變為淺棕色,此時莢果已完成生理成熟,且莢果的大小和干重均達到最大值,含水量降到最小值;盛花后25 d莢果腹縫線出現裂縫,盛花后35 d腹縫線完全裂開。(2)‘蘭箭3號’于盛花后20 d腹縫線處離層細胞開始解體;盛花后25 d,內、中、外果皮的薄壁細胞均開始失水皺縮,其中內果皮的薄壁細胞部分已開始破裂,離層細胞及其下面的薄壁細胞完全解體,外部果瓣緣細胞內側細胞壁破裂,但外側異常加厚的細胞壁仍然保持完整并連接兩個果瓣,使莢果不開裂;盛花后30~35 d,內、中、外果皮的薄壁細胞完全失水,細胞壁皺縮在一起,同時外部果瓣緣細胞外側細胞壁斷裂成兩部分,莢果的兩個果瓣裂開。研究表明,盛花后25~30 d莢果失去綠色變為淺棕色時是‘蘭箭3號’的適宜收獲時間,且離層和細胞失水產生的機械拉力是導致箭筈豌豆莢果開裂的主要原因,推測外部果瓣緣細胞外側增厚融合的細胞壁很可能是‘蘭箭3號’抵抗裂莢的關鍵結構。
箭筈豌豆;‘蘭箭3號’;莢果;發育動態;裂莢;適宜收獲時間;腹縫線
箭筈豌豆(Vicia.sativa)是豆科(Leguminosae)野豌豆屬(ViciaLinn.)一年生閉花授粉植物,普遍種植于中國的長江中下游、華北和西北等地區[1]。箭筈豌豆具有適應性廣、抗寒性強、營養價值高、經濟效益好等特點,是中國高海拔地區重要的牧草和綠肥作物,在中國草地農業系統中發揮著不可替代的作用[2-5]。南志標等通過在青藏高原地區引種野豌豆屬(Viciaspp.)和山黛豆屬(Lathyrusspp.),于2011年成功培育了適宜在高寒和高海拔地區種植的國家級牧草新品種‘蘭箭3號’,其具有早熟、生育期短、抗寒性強、種子產量高等特點,是中國高海拔地區的反芻動物的優質蛋白粗飼料[2,6]。
裂莢現象普遍存在于豆科植物中,對其種子生產造成巨大的損失[7,8]。確定種子適宜的收獲時間,減免裂莢的發生,可一定幅度上提高種子產量[9]。因此從19世紀60年代起一些研究者開始致力于種子適宜收獲時間的研究,避免收獲時間不當帶來的損失[10]。不同物種種子適宜收獲時間不同,研究方法也有很多。總體來說,莢果和種子的生長發育天數[11,12]、形態特征[12,13]和生理指標[12,14,15]等是種子適宜收獲確定的主要依據。
裂莢與莢果本身的組織結構有著密切聯系,在自然或物理機械壓力下,造成了莢果的某一位點開裂[9]。腹縫線是心皮邊緣閉合卷曲發育而來的,莢果首先沿著心皮愈合處的腹縫線開裂,然后由于機械扭曲力的作用使得背縫線開裂,因此腹縫線是莢果開裂的起始部位[16]。已有的對莢果解剖結構的研究發現離層、內果皮的內厚壁組織、中果皮的木質化細胞以及維管束等是莢果開裂及抗裂莢的重要結構[17-19]。在大豆裂莢的最新研究中發現,果莢開裂抗性的關鍵結構是腹縫線處位于離層細胞上方的纖維帽細胞,纖維帽細胞由細胞壁過度加厚的薄壁細胞組成,可以抵抗由離層細胞釋放的水解酶的作用,使其具有一定的裂莢抗性[19]。不同物種的莢果微觀結構不同,裂莢機理也就不同,目前裂莢區微觀結構的觀察在擬南芥[20-22]、大豆[19,23]、油菜[24,25]等模式植物和作物中相對較成熟,但在箭筈豌豆等重要豆科牧草中尚未有報道。本研究對‘蘭箭3號’箭筈豌豆各發育時期的莢果腹縫線進行解剖結構觀察,試圖找到與裂莢抗性相關的結構,為進一步研究箭筈豌豆的裂莢機理奠定基礎。
1 材料和方法
1.1實驗材料
本實驗所用材料‘蘭箭3號’均由草地農業生態系統國家重點實驗室提供。
1.2試驗設計
試驗地點設于蘭州大學草地農業科技學院榆中試驗田(35.57°N,104.09°E)。選取正常飽滿的種子種植于1 m × 20 m的小區,單行單株種植,株距20 cm,3次重復,其它條件相同。盛花期標記剛剛開放的花朵(以旗瓣還未展開為剛剛開放,如圖1盛花后第0天所示)。
1.3取樣與處理
盛花期標記的花朵,每5 d取樣1次,直至盛花后35 d。每個小區每次隨機取10個發育正常的莢果,用于莢果和種子的形態學和生理學特征觀察與分析。另外,每個小區隨機取若干個發育正常的莢果放入FAA固定液(70%乙醇∶冰乙酸∶甲醛=9∶0.5∶0.5)中,4 ℃冰箱中保存,用于電鏡掃描和半薄切片制備。
1.4莢果形態學特征
將各時期取樣的莢果,放置比例尺,用數碼相機拍照,隨后用Digimizer4.2圖像分析軟件對掃描圖像進行分析,測量莢果長、寬和厚,精確到0.01 mm。
1.5種子和莢果的生理學特征
將各時期取樣的莢果,分離莢皮和種子。用電子天平分別稱量每個莢果的莢皮和種子的鮮重,精確到0.001 g,并計算平均值。將莢皮和種子在80 ℃下烘干至恒重并稱量,精確到0.001 g,分別計算平均值和含水量。
含水量=鮮重-干重
1.6腹縫線表面結構掃描電鏡觀察
從FAA固定液中,取出各時期取樣的莢果,截取腹縫線中間的一段(約3 mm),利用JSM-5600掃描電鏡進行觀察,獲取掃描圖像。
1.7腹縫線橫截面解剖結構觀察
從FAA固定液中,取出各時期取樣的莢果,截取腹縫線中間的一段(約3 mm),70%(1 h)、80%(1 h)95%(1 h)、95%(1 h)、100%(1 h)、100%(1 h)、100%(1 h)乙醇梯度脫水,100%(1 h)、100%(1 h)二甲苯置換,預滲透液(Technovit 9100預滲透液∶二甲苯=1∶1)處理(1 h),用Technovit 9100預滲透液進行預滲透(24 h),Technovit 9100滲透液進行滲透(72 h),Technovit 9100混合液(A液∶B液=9∶1)包埋材料,放入4 ℃冰箱過夜。利用Leica RM2235手動旋轉式半薄切片機進行切片,厚度為2 μm,甲苯胺藍染色,再用電腦成像系統照相。
1.8數據分析
Excel 2016分析各時期莢果的長、寬、厚的平均值,各時期莢果和種子的鮮重、干重的平均值和其含水量,并用Origin軟件進行作圖。
2 結果與分析
2.1莢果的形態學特征
‘蘭箭3號’莢果的顏色隨生長時間的推移而發生顯著的變化(圖1)。在莢果生長發育的早期,盛花后0~20 d的莢果顏色為綠色;隨著莢果的進一步發育,盛花后25 d莢果顏色變為棕綠色;盛花后30~35 d莢果失去綠色,變為淺棕色。淺棕色是‘蘭箭3號’莢果成熟的顏色。
‘蘭箭3號’莢果的長、寬和厚在莢果發育過程中的變化如圖2所示。盛花后0~10 d是莢果長度和寬度的迅速生長期,于盛花后10 d左右達到最大值,此段時間莢果厚度生長相對較緩慢;盛花后10~20 d莢果厚度迅速增長,于盛花后20 d左右達到最大值,莢果厚度增加的原因是種子的生長,表明這段時間種子在迅速生長;盛花后20~35 d莢果寬度和厚度略有減小,是細胞失水皺縮造成的。
2.2莢果中莢皮和種子的生理學特征
‘蘭箭3號’莢果中莢皮的鮮重、干重和含水量隨其生長時間有明顯的變化(圖3)。莢皮的鮮重和含水量的變化趨勢一致,盛花后5~10 d迅速增加,隨后增速變緩,盛花后20 d左右達到最大值,盛花后20~30 d迅速減小,盛花后30~35 d兩者都基本不再發生變化;莢皮干重在盛花后0~20 d呈逐漸增加的趨勢,20 d左右達到最大值。
‘蘭箭3號’莢果中種子的鮮重、干重和含水量隨生長時間的變化如圖4所示。種子的鮮重和含水量的變化也基本保持一致,盛花后5~20 d迅速增加,盛花后20~30 d迅速減小,盛花后30~35 d基本保持不變;種子的干重在盛花后5~15 d緩慢增加,盛花后15~20 d迅速增加,盛花后20~30 d緩慢增加,盛花后30 d左右達到最大值。

d表示盛花后天數圖1 ‘蘭箭3號’莢果發育過程中莢果的顏色d represents days after peak anthesisFig. 1 Pod color during pod development of Lanjian NO.3
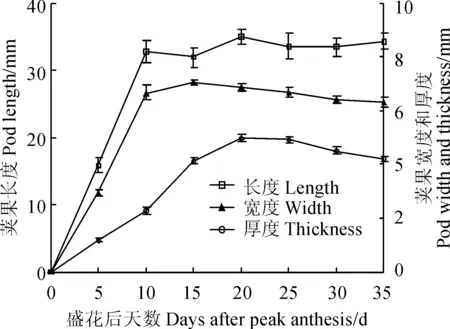
圖2 ‘蘭箭3號’莢果發育過程中的莢果的長度、寬度和厚度Fig. 2 Pod length, width and thickness during pod development of Lanjian NO.3
2.3莢果腹縫線表面結構的掃描電鏡觀察
‘蘭箭3號’莢果腹縫線的表面結構如圖5所示。隨著莢果的生長發育,表皮毛變疏,盛花后10~15 d在腹縫線中間長出一個凸起的結構(圖5,B、C),盛花后20~35 d凸起平展(圖5,D~F)。盛花后5~20 d腹縫線中間結構完整,沒有開裂跡象(圖5,A~D);盛花后25 d莢果沿著腹縫線中間開始裂開,并向兩邊延伸(圖5,E);盛花后30 d,腹縫線中間大部分已經裂開(圖5,F);盛花后35 d腹縫線中間已經完全開裂(圖5,G)。
2.4腹縫線橫截面的半薄切片觀察
‘蘭箭3號’莢果發育過程中腹縫線橫截面的微觀結構如圖6所示。盛花后5 d,莢果的兩個果瓣中間尚未分開,中果皮和內果皮有很小的厚壁細胞(圖6,A)。盛花后10 d,莢果兩個果瓣中間分開形成空腔,為種子發育提供空間,內果皮的厚壁細胞發育形成內厚壁組織;中果皮的厚壁細胞發育形成兩個分開的維管束,分別嵌入兩個果瓣中;兩個果瓣連接處的果瓣緣細胞分化形成離層細胞,與兩個維管束共同形成了一個帽子狀的結構;外部果瓣緣細胞外側的細胞壁明顯增厚,并相互融合在一起(圖6,B)。盛花后15 d,莢果微管束和內厚壁組織的細胞逐漸增多變大,細胞壁逐漸增厚;且維管束上端逐漸向外果皮延伸(圖6,C)。盛花后20 d,夾在兩個維管束之間的離層細胞開始解體(圖6,D、H)。盛花后25 d,內、中、外三個果皮的細胞開始失水皺縮,內果皮的薄壁細胞已經有一部分開始破裂;離層細胞及其下面的薄壁細胞完全解體;外部果瓣緣細胞內側細胞壁破裂,但是外側異常加厚的細胞壁仍然保持完整,連接兩個果瓣,使莢果不開裂(圖6,E)。盛花后30~35 d,莢果的兩個果瓣裂開,外部果瓣緣細胞外側細胞壁斷裂成兩部分;內果皮的薄壁細胞大部分完全破裂,靠近內厚壁組織的薄壁細胞的細胞壁皺縮在一起;并且外果皮和中果皮細胞完全失水,細胞壁皺縮在一起。
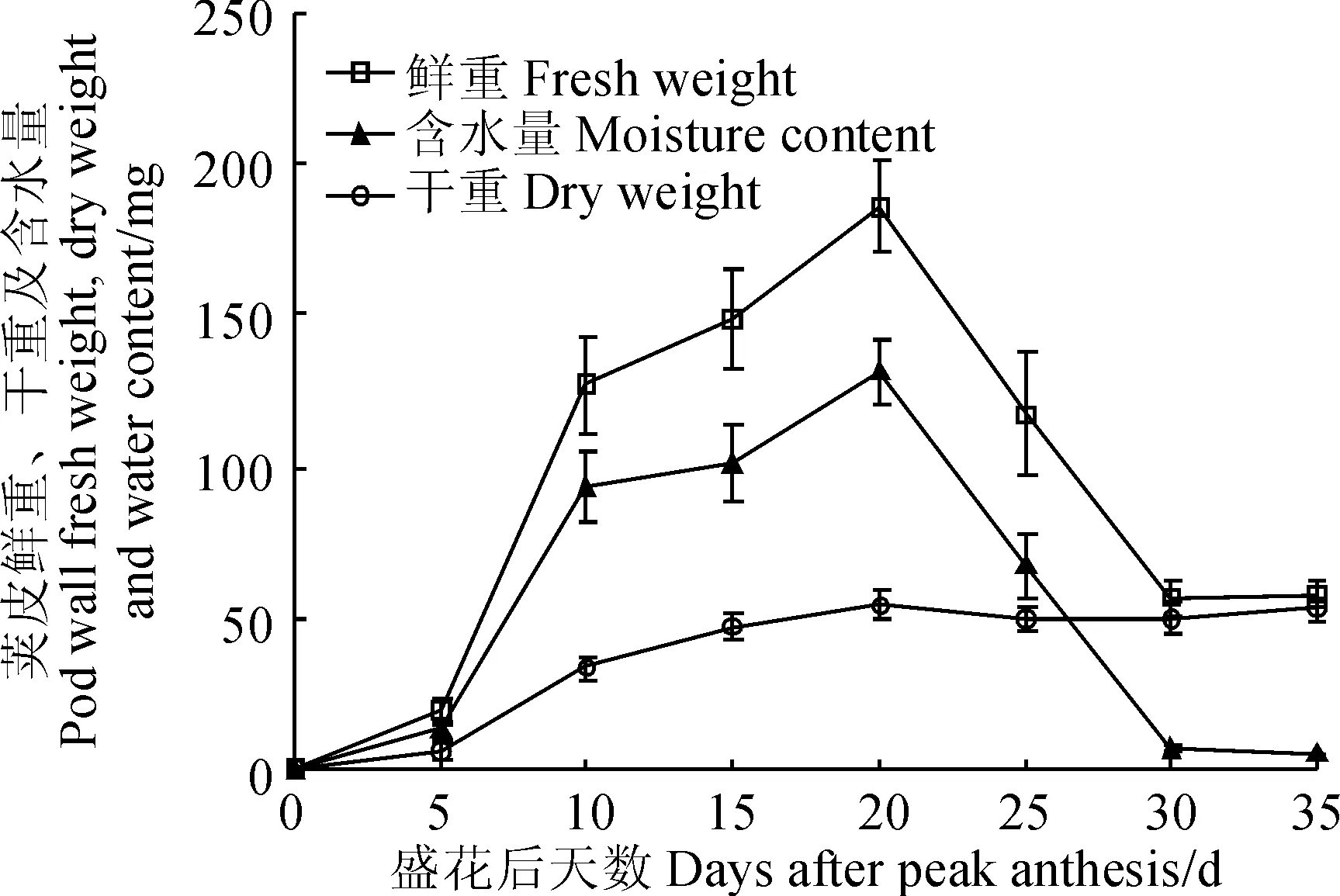
圖3 ‘蘭箭3號’莢果發育過程中莢皮的鮮重、干重和含水量Fig. 3 Pod wall fresh weight, dry weight and moisture content during pod development of Lanjian NO.3
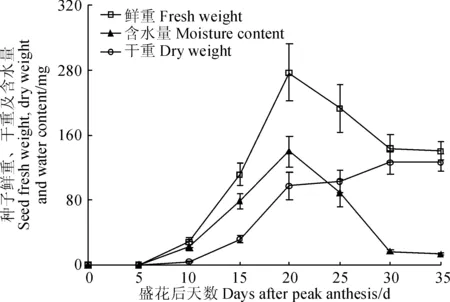
圖4 ‘蘭箭3號’莢果發育過程中種子的鮮重、干重和含水量Fig. 4 Seed fresh weight, dry weight and moisture content during pod development of Lanjian NO.3
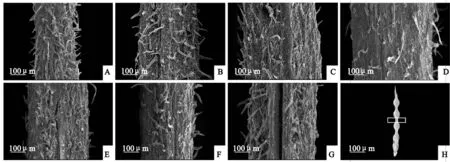
A~G. 盛花后5、10、15、20、25、30、35 d;H. 白框位置為取樣部位圖5 ‘蘭箭3號’莢果發育過程中莢果腹縫線的掃描電鏡圖片A~G. 5, 10, 15, 20, 25, 30 and 35 days after peak anthesis; H. The location of white frame is the sampling siteFig. 5 Scanning electron microscope pictures of pod ventral suture during pod development of Lanjian NO.3
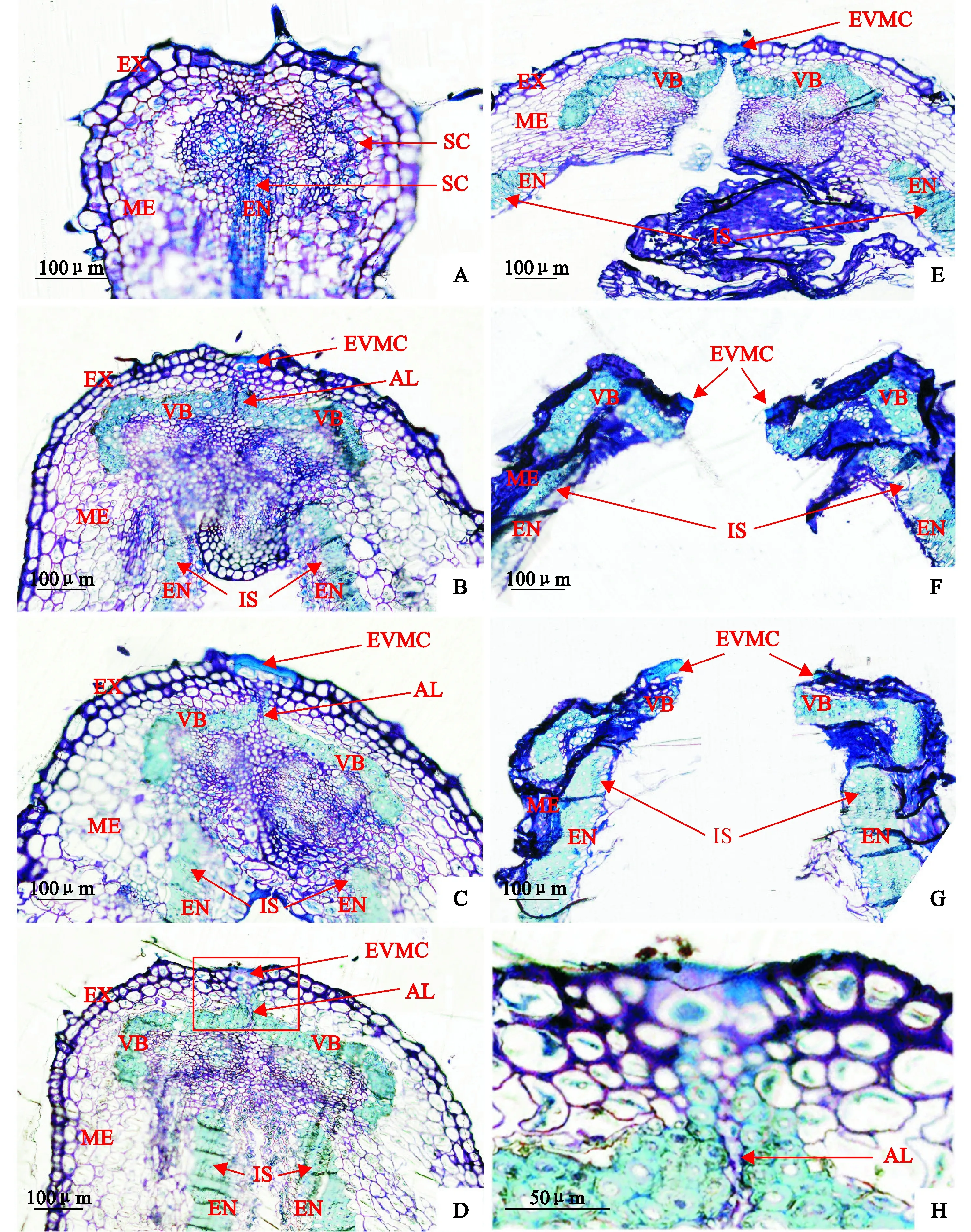
A~G. 盛花后5、10、15、20、25、30、35 d;H. D圖中紅框部分的放大圖像SC. 厚壁細胞;EVMC. 外部果瓣緣細胞;IS. 內厚壁組織;AL. 離層;VB. 維管束;EX. 外果皮;ME. 中果皮;EN. 內果皮圖6 ‘蘭箭3號’莢果發育過程中莢果腹縫線橫截面的半薄切片A~G. 5, 10, 15, 20, 25, 30 and 35 days after peak anthesis; H. Enlarged image of the red frame section in picture D.SC. Sclerenchymatous cells; EVMC. External valve margin cells; IS. Inner sclerenchyma; AL. Abscission layer; VB. Vascular bundle;EX. Exocarp; ME. Mesocarp; EN. EndocarpFig. 6 Semi-thin slices of pod ventral suture cross-section during pod development of Lanjian NO.3
3 討 論
裂莢對植物體本身來說是生命過程中重要的生理現象,對物種的繁殖和傳播有重要的生物學意義,但是在豆科牧草種子生產中,裂莢后的種子脫落會給農民帶來嚴重的損失[17]。過早收獲會導致種子成熟度差、活力低、質量差,過晚收獲又會造成種子的嚴重損失,因此適宜收獲間的確定可以有效減免因提前收獲和裂莢帶來的損失[10]。種子含水量是種子生理成熟和收獲時間的精確判斷指標[12]。本實驗通過對‘蘭箭3號’箭筈豌豆莢果發育動態的研究,發現盛花后25~30 d莢果失去綠色變為淺棕色時,莢果大小和干重不再發生明顯變化,莢皮和種子的含水量達到最低,表明此時‘蘭箭3號’的種子已經完成了生理成熟,且對裂莢起始部位腹縫線微觀結構的觀察也發現此時莢果已經出現開裂跡象,因此推斷盛花后25~30 d莢果失去綠色變為淺棕色時是‘蘭箭3號’種子的適宜收獲時間。本研究將發育動態與微觀結構聯系起來,使收獲時間的確定更加可靠,對‘蘭箭3號’種子生產具有重要的參考價值和實際指導意義。
離層和細胞失水產生的機械拉力是導致箭筈豌豆莢果開裂的主要原因。離層是莢果開裂不可或缺的結構[17]。本研究發現,‘蘭箭3號’莢果的離層是發育完全且有功能的,離層位于果瓣連接處,由非木質化的薄壁細胞組成的一條很窄的細胞帶。盛花后20 d莢果大小和莢皮含水量、干重已經達到最大(圖2、3),此時離層細胞開始發揮作用,釋放水解酶,溶解細胞壁使細胞解體,是莢果開裂的起始開關。莢果發育后期,細胞失水產生的機械力是導致莢果開裂的另一個重要原因[17]。本研究中,盛花后25 d莢皮細胞失水(圖3),把果瓣緣細胞向兩個方向拉扯,果瓣緣細胞屬于薄壁細胞,當其承受不了拉力時就會破裂,導致莢果開裂。值得注意的是起初內果皮失水最為嚴重,因而對內側的薄壁細胞拉力較大,推測可能是導致莢果先從內部開裂的原因。
內果皮的內厚壁組織和中果皮的維管束是分擔由細胞失水產生的機械力的重要結構。內果皮的內厚壁組織以及維管束由厚壁細胞組成,厚壁細胞的細胞壁由胞間層、初生壁和次生壁構成,起支撐作用。在細胞失水的過程中,內、中、外三個果皮的薄壁細胞紛紛失水皺縮,而內厚壁組織和維管束細胞仍然保持穩定的結構,分擔了大部分的機械拉力,是抵抗裂莢的重要結構。但是這些結構在大豆[19]、擬南芥[26]等其他豆科植物中也同樣存在,并不是箭筈豌豆所獨有的結構。百脈根屬的莢果解剖結構發現,抗裂莢品種朝著中果皮木質化程度減小的方向發展[27]。本研究發現,箭筈豌豆的莢果的中果皮并沒有木質化的趨勢,也是其抗裂莢的一個原因。
在腹縫線的電鏡掃描圖像中,盛花后10~15 d出現了一個表面較為光滑的凸起結構(圖5,B、C),且半薄切片的結果顯示盛花后10~15 d外部果瓣緣細胞外側細胞壁融合并劇烈加厚,融合后的細胞表面較為光滑(圖6,B、C),因此推斷該凸起結構即為外部果瓣緣細胞外側細胞壁異常加厚產生的。隨后莢皮失水,在內部拉力的作用下,果瓣緣細胞被向內拉伸,凸起逐漸展平,但細胞壁的厚度并沒有受到影響。從凸起的幅度可以看出該結構對腹縫線具有很大的保護作用,可在一定程度上抵抗裂莢的發生,同時盛花后25 d兩個果瓣僅靠外部果瓣緣細胞外側增厚融合的細胞壁連接,推測該結構很可能是箭筈豌豆抵抗裂莢的關鍵結構。
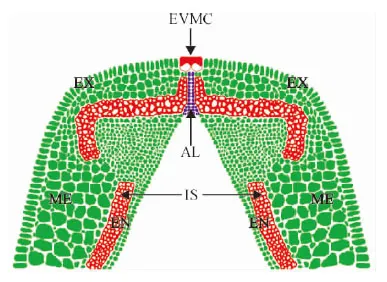
EVMC. 外部果瓣緣細胞;AL. 離層;VB. 維管束;IS. 內厚壁組織;EX. 外果皮;ME. 中果皮;EN. 內果皮圖7 箭筈豌豆腹縫線橫截面解剖結構示意圖EVMC. Extemal value margin cells; AL. Abscission layer; VB. Vascular bundle; IS. Inner sclerenchyma; EX. Exocarp mesocarp; ME. Mesocarp; EN. EndocarpFig. 7 Anatomical structure diagram of V. sativa pod ventral suture cross-section
根據‘蘭箭3號’各時期腹縫線橫切面的半薄切片觀察,繪制箭筈豌豆腹縫線橫切面的解剖結構示意圖(圖7),以便于對箭筈豌豆的解剖結構進行觀察和分析。在大豆裂莢的最新研究進展中指出,離層上方細胞壁過度增厚的纖維帽細胞是栽培大豆具有裂莢抗性的主要原因[19],但是箭筈豌豆的離層上方中并沒有發現細胞壁過度增厚的纖維帽細胞,取而代之的是外側細胞壁增厚融合的外部果瓣緣細胞。對比栽培大豆的解剖結構示意圖[26],可以發現箭筈豌豆的外側果瓣緣細胞與大豆的纖維帽細胞在位置上都處于離層上方且都與外部空氣相接觸,結構上都有過度增厚的細胞壁,功能上都是抵抗離層細胞釋放的水解酶的作用,牢固連接兩個果瓣使莢果不開裂,因此推測它們具有一定的同源性,但需要借助分子手段等進一步驗證。該研究結果為進一步探討箭筈豌豆的裂莢機理提供了解剖結構基礎,有望在裂莢抗性的研究方面取得新的研究進展。
[1]陳默君, 賈慎修. 中國飼用植物[M]. 北京:中國農業出版社, 2002: 673-675.
[2]南志標,張吉宇,等.5個箭筈豌豆品系基因型與環境互作效應及農藝性狀穩定性[J].生態學報,2004,24(3):395-401.
NAN Z B, ZHANG J Y,etal.Genotype×environment interactions and consistency analysis for agronomic characteristics of fiveViciasativalines[J].ActaEcologicaSinica, 2004,24(3):395-401.
[3]董德珂,董瑞,劉志鵬,等.532份箭筈豌豆種質資源復葉表型多樣性[J].草業科學,2015,36(2):536-541.
DONG D K ,DONG R ,LIU Z P ,etal.Diversity analysis of the compound leaf phenotypic characters of 532Viciasativagermplasm resources[J].PrataculturalScience, 2015,36(2):536-541.
[4]LIU Z P, LIU P, LUO D,etal. Exploiting illumina sequencing for the development of 95 novel polymorphic EST-SSR markers in common vetch (Viciasativasubsp.sativa)[J].Molecules, 2014, 19(5): 5 777-5 789.
[5]LIU Z P, MA L C, NAN Z B,etal. Comparative transcriptional profiling provides insights into the evolution and development of the zygomorphic flower ofViciasativa(Papilionoideae)[J].PloSONE, 2013, 8(2): e57338.
[6]劉鵬,馬利超,王宇,等.野豌豆屬牧草種質花粉形態的掃描電鏡觀察[J].草業科學,2015,32(6):978-987.
LIU P ,MA L C ,WANG Y ,etal.Observation on pollen morphology ofViciagermplasms using scanning electron microscope[J].PrataculturalScience, 2015,32(6):978-987.
[7]PHILBROOK B D, OPLINGER E S. Soybean field losses as influenced by harvest delays[J].AgronomyJournal, 1989, 81(2): 251-258.
[8]張躍進,馬賽斐,高啟云,等.黃淮流域主栽大豆品種炸莢性研究[J].河南農業科學,2006(6):56-59.
ZHANG Y J ,MA S F ,GAO Q Y ,etal.Study on the pod-shattering of main soybean varieties of Huanghuai area[J].JournalofHenanAgriculturalScience, 2006,(6):56-59.
[9]孫東鳳,康玉凡.大豆炸莢性研究進展[J].大豆科學,2011,(6):1 030-1 034.
SUN D F ,KANG Y F .Advances in studies of pod-shattering of soybean[J].SoybeanScience,2011,(6):1 030-1 034.
[10]王穎. 廣布野豌豆莢果和種子發育動態及種子適宜收獲時間的確定[D].長春:東北師范大學, 2008.
[11]毛培勝,韓建國,吳喜才.收獲時間對老芒麥種子產量的影響[J].草地學報,2003,1(11):33-37.
MAO P S ,HAN J G ,WU X C .Effects of harvest time on seed yield of siberian wildrye[J].ActaAgrestiaSinica, 2003,1(11):33-37.
[12]ELIAS S G, COPELAND L O. Physiological and harvest maturity of canola in relation to seed quality[J].AgronomyJournal, 2001, 93: 1 054-1 058.
[13]EASTIN J D, HULTQUIST J H,etal. Physiologic maturity in grain sorghum[J].CropScience, 1973, 13: 175-178.
[14]FRASER J, EGLI D B, LEGGETT J E. Pod and seed development in soybean cultivars with differences in seed size[J].AgronomyJournal, 1982, 74: 81-85.
[15]WANG P, ZHOU D W,etal. Seed maturity and harvest time effects seed quantity and quality of HORDEUM BREVISUBULATUM[J].SeedScienceTechnology, 2006, 34: 125-132.
[16]SUZUKI M, FUJINO K,etal. A major soybean QTL,qPDH1, controls pod dehiscence without marked morphological change[J].PlantProductionScience, 2009, 12(2): 217-223.
[17]羅汝葉,鞏鵬濤.植物落粒性狀研究進展[J].豆科基因組學與遺傳學,2011,2(1):1-13.
LUO R Y ,GONG P T .Research progress of seed-shattering habit in plants[J].LegumeGenomicsandGenetics,2011,2(1):1-13.
[18]羅棟,王彥榮,劉志鵬.豆科植物裂莢生物學基礎的研究進展[J].草地學報,2015,23(5):927-935.
LUO D ,WANG Y R ,LIU Z P .Research progress in biological basis of legume pod dehiscence[J].ActaAgrestiaSinica,2015,23(5):927-935.
[19]DONG Y, YANG X, LIU J,etal. Pod shattering resistance associated with domestication is mediated by a NAC gene in soybean[J].NatureCommunications, 2015, 5(2): 3 352-3 352.
[20]DINNENY J R, DETLEF W, YANOFSKY M F. A genetic framework for fruit patterning inArabidopsisthaliana[J].Development, 2005, 132(21): 4 687-4 696.
[22]CAROL W, YANG T J W,etal. A molecular and structural characterization of senescingArabidopsissiliques and comparison of transcriptional profiles with senescing petals and leaves[J].PlantJournal, 2009, 57(4): 690-705.
[24]KADKOL G P, BEILHARZ V C,etal. Anatomical basis of shatter-resistance in the oilseed brassicas[J].AustralianJournalofBotany, 1986, 34(34): 595-601.
[25]CHILD R D, BRUCE D M. Increased resistance to pod shatter is associated with changes in the vascular structure in pods of a resynthesizedBrassicanapusline[J].JournalofExperimentalBotany, 2003, 54(389): 1 919-1 930.
[26]DONG Y, WANG Y Z. Seed shattering: from models to crops[J].FrontiersinPlantScience, 2015, 6: 476.
[27]YANG J B, SOMERS D A, WRIGHT R L,etal. Seed pod dehiscence in birdsfoot trefoil,Lotusconimbricensis, and their interspecific somatic hybrid[J].CanadianJournalofPlantScience, 1990, 70(1): 279-284.
(編輯:潘新社)
Study on Pod Development and Ventral Suture Structure ofViciasativaCultivar Lanjian NO.3
DONG Deke, DONG Rui, WANG Yanrong, NIE Bin, LIU Zhipeng*
(College of Pastoral Agriculture Science and Technology, Key Laboratory of Grassland Farming Systems, Lanzhou University, Lanzhou 730020, China)
The common vetch (Viciasativa) is an important annual legume forage at high altitudes. However, the phenomenon of pod shattering when it matured would casue serious loss of seeds. WithV.sativacultivar Lanjian NO.3 as the research object, this study analyzed dynamic changes in pod morphological, moisture content, surface structure of ventral suture, anatomical structure of ventral suture cross section during pod development in order to explore the pod shattering mechanism ofV.sativaand provide theoretical basis for determining its optimal seed harvesting time in production. The results showed that: (1) When the pod of Lanjian NO.3 turned to light brown at about 25-30 days after peak anthesis, it had completed physiological maturity and its size and dry weight had reached the highest with the moisture content had reduced to the lowest. A crack appeared in the pod ventral suture at 25 days after peak anthesis and it separated completely at 35 days after peak anthesis. (2) The abscission layer cells began to disintegrate at 20 days after peak anthesis. At 25 days after peak anthesis, all the parenchyma cells of endocarp, mesocarp and exocarp began to lose water and shrink wherein the portion of endocarp parenchyma cells began to break. The abscission layer cells and the following parenchyma cells disintegrated completely. The inside cell walls of external valve margin cells broke but the outside cell walls which abnormal thickened still remained intact and connected two fruit valves which made the pod do not crack. During the period from 25 to 30 days after peak anthesis, the parenchyma cells of endocarp, mesocarp and exocarp lost water completely and their cell walls shrank together. At the same time the outside cell walls of external valve margin cells broke into two parts and the two fruit valves of the pod cracked. The research shows that the optimal harvest time of Lanjian NO.3 seeds was the period from 25 to 30 days after peak anthesis when the pod turned to light brown. The abscission layer cells and machine tensile which generated by cell water loss were the main causes ofV.sativapod crack. Conjecture that the thickening and fused cell walls of external valve margin cells were the key structure resistant to pod shattering probably.
V.sativa; Lanjian NO.3; pod; development dynamic; pod shattering; optimum harvest time; ventral suture
1000-4025(2016)07-1376-07
10.7606/j.issn.1000-4025.2016.07.1376
2016-01-30;修改稿收到日期:2016-07-12
國家重點基礎研究發展計劃(973計劃)(2014CB138704)
董德珂(1991-),在讀碩士研究生,主要從事草類作物遺傳育種研究。E-mail:dongdk10@lzu.edu.cn
劉志鵬,教授,博士生導師,主要從事牧草種質資源與育種研究。E-mail:lzp@lzu.edu.cn
Q944.59
A
