新城疫病毒F蛋白納米抗體的篩選及活性鑒定
2016-09-07 10:06:04高小龍胡湘云付向晶王燕平仝麗娜王海新王興龍張淑霞杜恩岐楊增岐
畜牧獸醫學報 2016年8期
高小龍,胡湘云,付向晶,王燕平,仝麗娜,王海新,王興龍,張淑霞,蕭 颯,杜恩岐,楊增岐
(西北農林科技大學 動物醫學院,楊凌 712100)
?
新城疫病毒F蛋白納米抗體的篩選及活性鑒定
高小龍△,胡湘云△,付向晶,王燕平,仝麗娜,王海新,王興龍,張淑霞,蕭颯,杜恩岐*,楊增岐*
(西北農林科技大學 動物醫學院,楊凌 712100)
旨在篩選新城疫病毒(NDV)F蛋白的納米抗體,并對篩選的抗體活性進行初步鑒定。以NDV F蛋白中和表位構建誘餌,利用酵母雙雜交技術對雙峰駝天然重鏈抗體可變區(VHH)酵母雙雜交文庫進行篩選,獲得4株VHH抗體序列。挑選其中2株最符合VHH特征的進行原核表達與純化,并對2株VHH活性進行鑒定。ELISA結果顯示,VHH1與VHH2與NDV病毒的反應性顯著高于陰性對照(P<0.05),表明VHH與NDV病毒反應活性良好;Western blot結果顯示,2株VHH可特異性識別F蛋白;細胞中和試驗結果表明,2株VHH抗體均具有一定的中和活性。以上結果提示本研究成功篩選出2株活性較好的抗NDV F蛋白的VHH抗體,為VHH在NDV防控中的應用奠定了基礎。
新城疫病毒;F蛋白;VHH抗體;酵母雙雜交
新城疫(Newcastle disease,ND)是由新城疫病毒(Newcastle disease virus,NDV)引起的一種嚴重危害養禽業的重要疫病,被OIE列為必須報告的疫病。NDV屬于副黏病毒科禽腮腺炎病毒屬,其基因組為不分節段的單鏈負股RNA,大小約15 kb,依次編碼6種結構蛋白。融合蛋白(fusion protein,F)是病毒囊膜上一個非常重要的糖蛋白,可介導病毒與宿主細胞的膜融合,同時還是誘導機體保護性免疫應答的主要抗原。研究表明,針對F蛋白的抗體可有效阻止病毒對宿主細胞和機體的侵染[1]。因此,F蛋白是NDV防控中的理想靶標。
NDV疫苗的廣泛使用使ND大規模流行得到了有效控制。然而,近年來ND的流行出現了新的特點,如免疫雞群仍可發生ND,且可從中分離到致病性NDV[2-4]。研究表明,現有商品化疫苗和基于常規抗體的治療類制劑不能完全阻止ND發生[5-6]。同時,珍稀野生鳥類發生ND后往往缺乏特異性治療藥物[7],且在應用傳統的多抗和單抗做治療時存在免疫原性強、潛在的生物安全不明確等缺點。因此,開發新的治療性藥物尤為必要。
VHH(variable domains of camelid heavy-chain antibodies,VHH)抗體衍生于駱駝科動物體內存在的重鏈抗體[8-9]。由于其相對分子質量小(~15 ku)、結構穩定、可溶性好、組織穿透力強、免疫原性低且可識別隱蔽表位等特點,使其在病毒性疾病診斷和治療上具有廣闊的前景[10],但目前仍未見抗NDV的VHH報道。本研究為了獲得F蛋白的VHH抗體,以F蛋白表位為誘餌對VHH抗體文庫進行了篩選,成功獲得2株VHH抗體,并對其活性進行了初步鑒定,為應用VHH防控ND奠定基礎。
1 材料與方法
1.1病毒、菌株、質粒、試劑
NDV La Sota毒株和F48E9毒株均由本實驗室保存。Y2H Gold菌株、pGBKT7、pGADT7、pGADT7-T、pGADT7-p53、pGADT7-Lam質粒及篩庫所用試劑購自Clontech公司。雙峰駝天然重鏈抗體可變區酵母雙雜交文庫由本實驗構建與保存[11]。pHSIE原核表達質粒(帶6×His標簽和SUMO標簽)由本實驗室保存。2×Taq Master mix、HRP標記抗His單克隆抗體和cECL發光試劑盒購自康為世紀公司。IPTG購自Sigma公司,TMB購自天根生化科技(北京)有限公司,2K plus DNA marker購自全式金生物有限公司,限制性內切酶均購自Fermentas公司。其他無機試劑均為國產分析純。
1.2引物及基因合成
根據GenBank上公布的VHH序列,設計1對克隆VHH的引物。參考文獻中NDV F蛋白線性中和表位序列(LLPNLPKDKEACAKAPL)[12],合成一段由Linker(GGGS)串聯重復3次的中和表位DNA片段,命名為F-neu,并在前段和末端分別引入EcoRⅠ、SalⅠ酶切位點(圖1)。引物和基因片段均由蘇州金唯智生物科技有限公司合成。

圖1 F-neu編碼序列結構Fig.1 The structure of synthesised F-neu
1.3誘餌質粒構建
將合成的F-neu和pGBKT7質粒用EcoRⅠ和SalⅠ雙酶切,回收切出的F-neu片段和pGBKT7質粒,在T4 DNA連接酶的作用下16 ℃連接過夜;次日將連接產物轉化DH5α感受態細胞,挑取單克隆搖菌后抽提質粒并做雙酶切鑒定,酶切和測序正確后命名為pGBKT7-F-neu。
1.4文庫篩選
將pGBKT7-F neu質粒按照Clontech公司酵母細胞轉化手冊轉化Y2HGold酵母菌,進行誘餌的自激活和毒性驗證。文庫篩選時,取Y2H Gold(pGBKT7-F neu)與文庫菌Y187(pGADT7-VHH)做雜交,方法參照Clontech公司MatchmakerTMGold Yeast Two-Hybrid protocol。雜交產物涂布70個150 mm的DDO/X/A平板,30 ℃倒置培養5 d后,將平板上發藍的菌落挑到QDO/X/A平板上再次做篩選。QDO/X/A平板上仍生長且發藍的菌落進行酵母質粒拯救,拯救的文庫質粒與誘餌質粒共轉Y2H Gold酵母菌做進一步回轉驗證。回轉驗證仍然發藍的質粒進行測序比對分析,以確定篩選出的VHH抗體序列信息。
1.5VHH的表達與純化
以拯救的pGADT7-VHH質粒為模板,用引物VHH-PF (5′-CGGGATCCCAGGTGCAGCTGGT-GGAGTCTGGRGGAGG-3′)和VHH-PR (5′-GC-GTCGACTTAGCTGGAGACGGTGACCWGGG-T-3′)在TaqDNA聚合酶的作用下進行PCR擴增,回收擴增出的VHH片段,通過BamHⅠ和SalⅠ酶切位點將VHH克隆至pHSIE載體6×His-SUMO標簽下游以進行融合表達,酶切和測序鑒定正確后命名為pHSIE-VHH。
將pHSIE-VHH質粒轉化Rosetta感受態細胞進行原核表達。表達時,從平板上挑取單菌落接種于氨芐LB培養基中,37 ℃振蕩培養至OD600nm值為0.4~0.6時,加入終濃度為0.4 mmol·L-1的IPTG,于20 ℃誘導6 h后收集菌體,超聲裂解破碎后取上清按Talon Metal Affinity Resin(Clontech)說明書進行蛋白質純化,純化產物進行12% SDS-PAGE電泳分析。
1.6間接ELISA
將NDV病毒用包被液稀釋后包被酶標板,4 ℃過夜;次日棄包被液,PBST洗4次,用5%的脫脂奶粉進行封閉,37 ℃孵育1 h;棄封閉液,PBST洗板4次,每孔加100 μL稀釋好的VHH,37 ℃孵育2 h;棄掉孔中的抗體,PBST洗4次,加入100 μL HRP標記的抗His單克隆抗體(1∶5 000),37 ℃孵育1 h;棄去孔中的抗體,PBST洗板4次,加入100 μL TMB顯色液,室溫避光顯色20 min,加入50 μL 2 mol·L-1H2SO4終止反應,于OD450 nm波長處測定吸光度值。
1.7Western blot
將純化的LaSota病毒200倍稀釋進行SDS-PAGE電泳,電泳結束后轉膜。轉膜完成后,5%脫脂牛奶4 ℃封閉過夜;次日,TBST洗膜,加入稀釋的VHH抗體室溫孵育3 h;TBST洗膜,加入HRP標記抗His單克隆抗體(1∶6 000)室溫孵育1.5 h;TBST洗膜后,用ECL發光試劑于暗室中壓片。
1.8中和試驗
將DF-1細胞鋪96孔板,次日,待細胞長滿80%~90%時,進行以下操作:用PBS將VHH做梯度稀釋,取50 μL VHH與50 μL F48E9(100 TCID50)混勻后,置37 ℃孵育1 h;用PBS洗細胞一次,將孵育的VHH-病毒混合液加到細胞中,于CO2培養箱中37 ℃孵育1 h,每隔15 min振蕩混勻一次;棄掉各孔中的VHH-病毒混合液,PBS洗細胞一次,加入100 μL維持液于CO2培養箱中37 ℃培養36~72 h,每個稀釋度重復8個孔,24 h后觀察并記錄每孔細胞病變,按下列公式計算中和活性:中和活性(%)=(無病變孔數/總接種孔數)×100。
2 結 果
2.1誘餌質粒構建
將構建的pGBKT7-F-neu質粒用EcoRⅠ和SalⅠ做雙酶切,切出了204 bp的目的條帶,結果如圖2所示。并且測序結果正確,表明誘餌質粒構建成功。
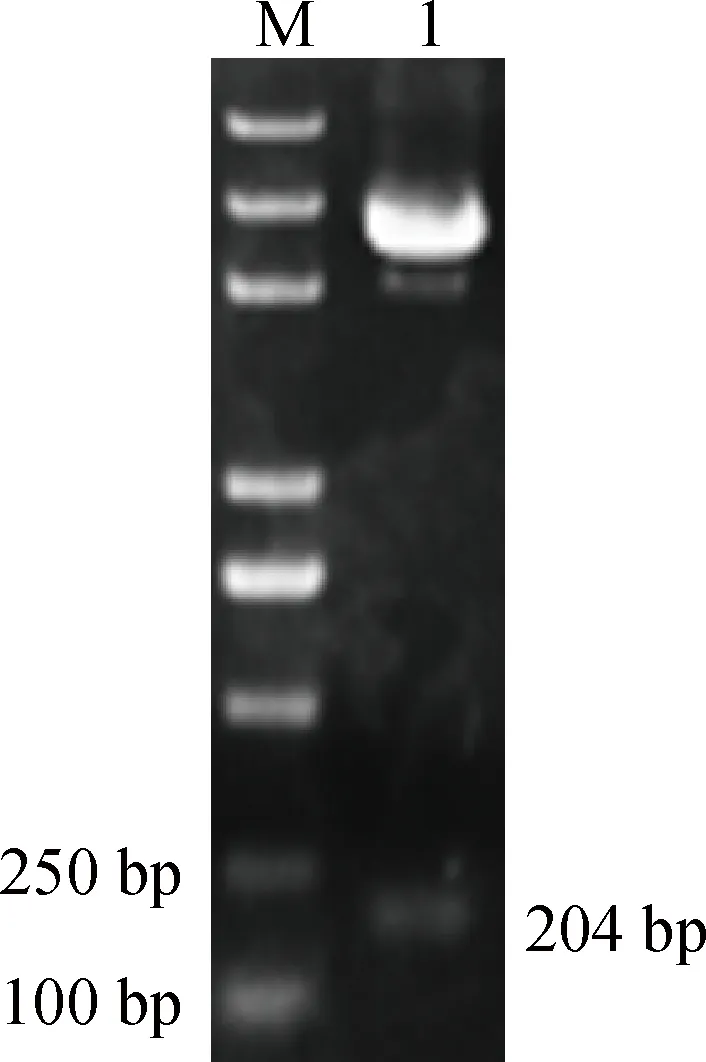
M.5 000的DNA相對分子質量標準標準;1.pGBKT7-F-neu雙酶切M.5000 DNA marker;1.Digestion of pGBKT7-F-neu圖2 誘餌質粒pGBKT7-F-neu雙酶切鑒定Fig.2 Restriction enzyme digestion of bait plasmid pGBKT7-F-neu
2.2文庫篩選
將篩選的陽性克隆進行質粒拯救,拯救的pGADT7-VHH質粒與pGBKT7-F-neu質粒共轉Y2H Gold做回轉驗證,同時設立pGBKT7-Lam+pGADT7-T和pGBKT7-p53+pGADT7-T為陰陽性對照。結果如圖3所示,篩選出的4個VHH與pGBKT7-F-neu共轉后在DDO/X/A和QDO/X/A平板上菌落均為藍色,VHH與pGBKT7空載體共轉后在DDO/X/A和QDO/X/A上均不變藍或不生長,且陰陽性對照均成立,表明篩選到的4株VHH為真陽性。
2.3序列分析
將4個陽性克隆進行測序和比對分析,結果如圖4所示,可知篩選出的4株VHH與駱駝源重鏈抗體重鏈可變區高度同源,且第1框架區(framework region 1,FR1)區和FR3區有2個保守的半胱氨酸,同時FR2區37位、44位、45位和47位存在特征性氨基酸替換。
2.4VHH表達與純化
從4株VHH中,挑選VHH1和VHH2進行原核表達。以拯救的pGADT7-VHH1和pGADT7-VHH2質粒分別為模板進行PCR,擴增出大小約400 bp的VHH片段,結果如圖5A。將VHH片段膠回收后,通過BamHⅠ和SalⅠ酶切位點克隆至pHISE表達載體上,雙酶切鑒定結果如圖5B所示,成功切出400 bp的目的條帶,且測序鑒定均正確。

A列.白色克隆表示誘餌蛋白無自激活和毒性;B列~E列.藍色克隆表示F-neu與VHH在酵母細胞內可互作;F列.白色和藍色克隆分別表示陰性和陽性對照Lane A.The white clones showed that the bait has no autoactivator activity in yeast;Lane B to E.When pBD-F and 4 positive pAD-VHH were cotransformed into Y2H Gold yeast strain,all cotransformants turned blue on DDO/X/A and QDO/X/A;Lane F.The white and blue clone indicated the negative and positive control,respectively圖3 誘餌蛋白F-neu與VHH在酵母細胞內互作結果Fig.3 Interaction between F-neu and VHH in yeast
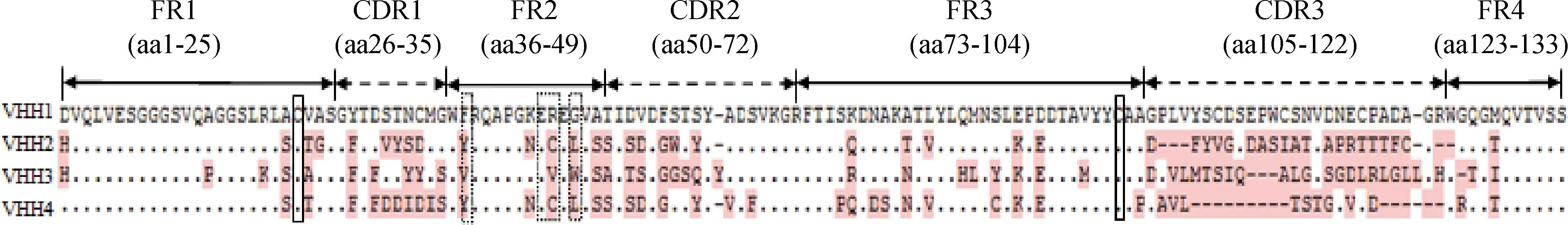
“.”表示與VHH1序列相同的氨基酸,不同的氨基酸用粉色字母表示;“-”表示此處氨基酸缺失;實線框內的為保守型半胱氨酸殘基,虛線框內的為4個特征性氨基酸殘基The dot indicate the same sequences compared with VHH1.Differences in the sequences are pinked,and the dash represent the missing sequences.Two hallmark Cys residues are labeled by the thick-line boxes.The four conservative hallmark residues of VHH in FR2 are labeled by the dotted line boxes圖4 4株篩選出來的VHH氨基酸序列比對結果Fig.4 Deduced amino acid alignment of four selected VHH
將pHSIE-VHH1和pHSIE-VHH2質粒轉化Rosetta感受態細胞進行誘導表達,表達產物用Talon樹脂進行純化,最后用含200 mmol·L-1咪唑的洗脫液洗脫目的蛋白質,結果如圖6所示,成功純化出約30 ku的VHH抗體。
2.5間接ELISA
用NDV LaSota病毒包被酶標板,封閉結束后,先加入純化的VHH抗體進行孵育,洗板后,再加入HRP標記的抗His單克隆抗體進行孵育,最后用TMB顯色液顯色,結果如圖7所示,VHH1和VHH2與NDV病毒的反應性顯著高于陰性對照(P< 0.05),與其他病毒不反應(OD450 nm<0.1),且陰性對照的OD450nm<0.1。表明VHH1和VHH2與NDV反應性良好。
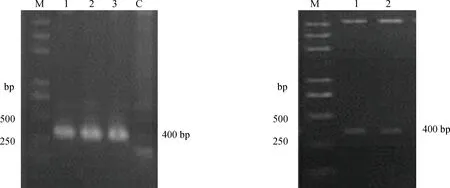
(A) VHH片段的PCR擴增:M.5000 DNA相對分子質量標準;1.VHH1的PCR擴增結果;2.VHH2的PCR擴增結果;3.陽性對照;C.陰性對照。(B) pHSIE-VHH雙酶切鑒定:M.5000 DNA相對分子質量標準;1.pHSIE-VHH1酶切結果;2.pHSIE-VHH2酶切結果(A) The amplification of VHH,M.5000 DNA marker;1.PCR products of VHH1;2.PCR products of VHH2;3.Positive control;C.Negative control.(B)Double enzyme digestion of pHSIE-VHH,M.5000 DNA marker;1.Digestion of pHSIE-VHH1;2.Digestion of pHSIE-VHH2圖5 VHH原核表達質粒構建Fig.5 Construction of VHH prokaryotic expression vector
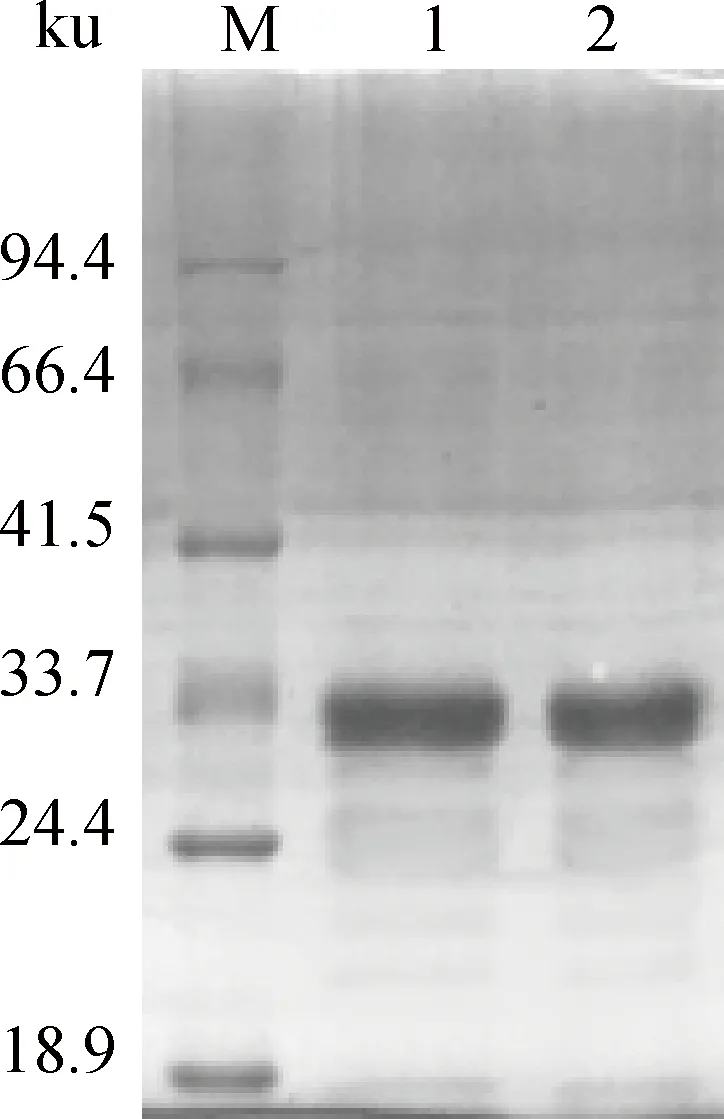
M.蛋白質相對分子質量標準;1.VHH1;2.VHH2M.Protein marker;1.VHH1;2.VHH2圖6 VHH抗體的純化結果Fig.6 Purification of VHH
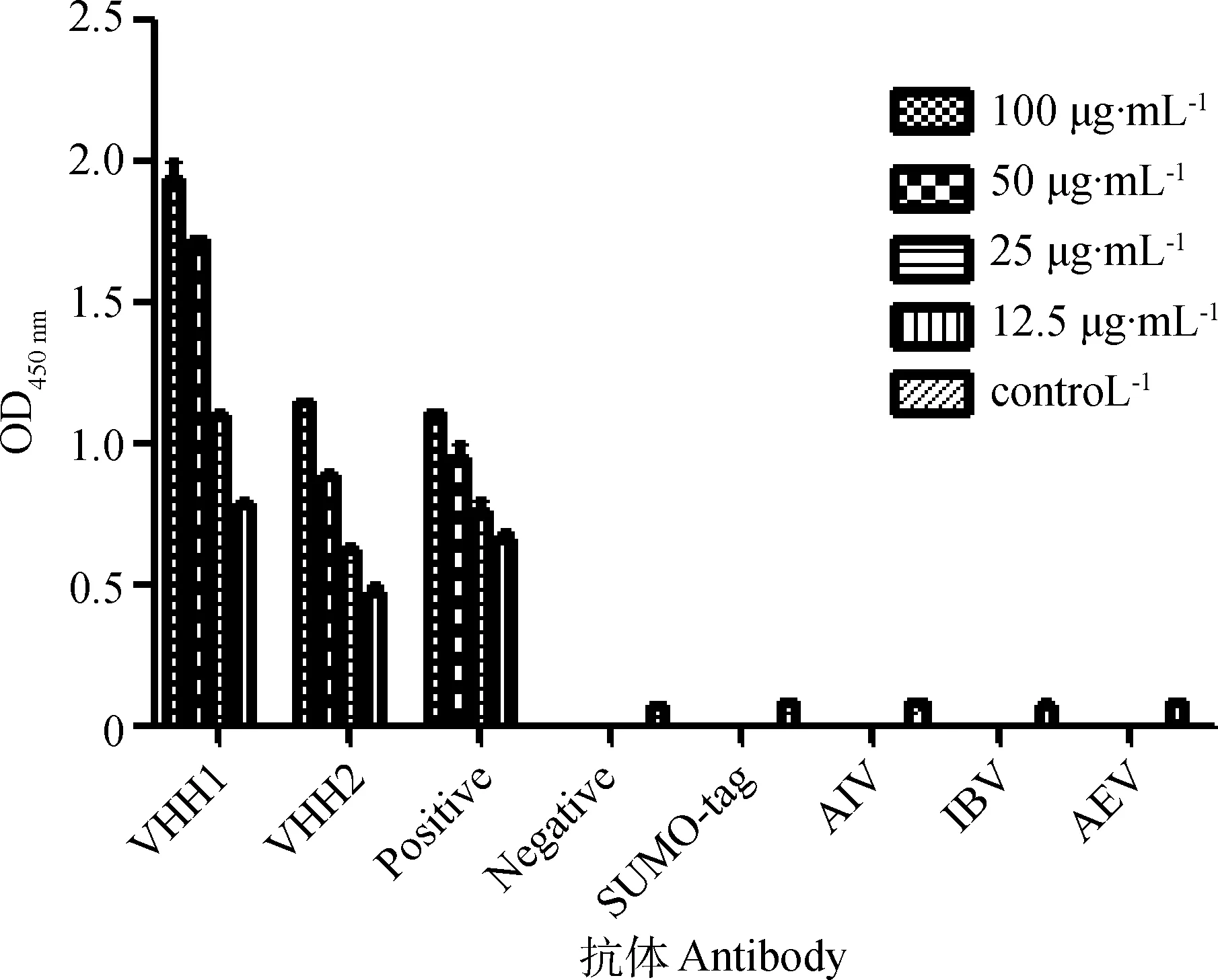
圖7 間接ELISA檢測VHH與NDV病毒的反應性Fig.7 Detection the interaction between the selected VHHs and NDV virion by indirect ELISA
2.6Western blot
用純化的VHH抗體進行Western blot后,結果如圖8所示,VHH1和VHH2均能識別大小約55 ku的F蛋白特異性條帶,而空載體不反應,表明VHH1和VHH2可識別F蛋白。

M.蛋白質相對分子質量標準;1.VHH1;2.VHH2;3.空載體的表達菌裂解液M.Protein marker;1.VHH1;2.VHH2;3.The bacteria lysate of empty vector圖8 Western blot 結果Fig.8 The result of Western blot
2.7中和試驗
將純化的2株VHH抗體稀釋后進行中和試驗。結果如圖9所示,24 h以內VHH1和VHH2在低稀釋度時(≤ 23)均能有效中和F48E9病毒在DF-1細胞上的增殖,細胞無合胞體出現;當抗體稀釋度高于24時,VHH抗體的中和活性逐漸下降,出現合胞體的孔數逐漸增多。隨著培養時間的延長,即36 h之后,各稀釋度孔中合胞體也會逐漸出現并增多。表明VHH1和VHH2在病毒感染早期具有一定的中和活性。

圖9 中和活性的測定Fig.9 Detection of neutralizing titer
3 討 論
VHH抗體來源于駱駝胚系基因組上一套獨特V基因片段的V-D-JH重排[13],并且發現目前VHH至少存在4個獨特的亞群。盡管VHH抗體存在不同的亞群,但相對VH抗體而言各亞群均存在一些特征性氨基酸替換,即Val37-Phe/Tyr、Gly44-Glu/Gln、Leu45-Arg、Trp47-Gly/Leu,上述疏水性氨基酸向親水性氨基酸的替換增加了VHH抗體的親水性,同時彌補了缺失輕鏈而造成的不穩定和易聚集的缺點。本研究中,通過用F蛋白中和表位構建誘餌質粒對天然VHH抗體文庫進行篩選,初步得到4株VHH抗體;對篩選到的4株VHH進行序列比對分析后發現,4株抗體37、44、45和47位存在VHH抗體的特征性氨基酸,與先前報道的一致[14]。同時發現所篩選的VHH抗體氨基酸差異主要集中在抗原互補決定區(complement determine region,CDR),FR區則相對保守,與之前報道一致[13,15]。
VHH抗體由于相對分子質量小,為單域結構,并且FR2區4個疏水性氨基酸位點發生了替換,增強了親水性,使其可以在大腸桿菌和酵母菌中高效表達[16]。本研究中,前期在表達VHH抗體時選用了pColdTMI DNA載體,在進行誘導表達時發現VHH可溶性差,多為包涵體形式存在,與X.J.Fu等報道的不一致[17],可能與抗體自身序列有關。小泛素樣相關修飾蛋白(small ubiquitin-related modifier,SUMO)是一類廣泛存在于真核細胞的高度保守的蛋白質[18],研究表明,當給異源蛋白N段融合SUMO后,不僅可以促進異源蛋白質的正確折疊和增加表達水平,而且還能保護靶蛋白免受降解,起到分子伴侶的作用[19]。為了增加VHH抗體的可溶性可表達量,本研究中后來選取了帶有SUMO助溶標簽的pHSIE原核表達載體,誘導表達后發現重組VHH抗體表達量得到提高,并且多為可溶性表達。表明SUMO標簽具有很好的助溶作用,與先前的報道一致[19-21]。
為了探究純化的VHH抗體與NDV病毒和F蛋白的反應性,本研究進行了ELISA和Western blot。ELISA結果顯示,篩選的VHH1抗體與LaSota病毒的反應性顯著高于陰性和陽性對照(P<0.05),VHH2抗體與病毒反應性顯著高于陰性對照(P<0.05),但與陽性對照差異不顯著(P>0.05),表明表達的2株VHH與NDV病毒反應性良好。Western blot結果顯示,表達的VHH1和VHH2可識別55 ku左右的特異性條帶,而陰性對照不反應,表明篩選的VHH抗體能與F蛋白反應。中和試驗結果顯示,VHH在稀釋度低時(≤ 23)可有效中和病毒,但36 h后會細胞會逐漸出現合胞體病變,且中和能力顯著低于陽性對照血清(≥ 28,P<0.05);可能與以下因素有關:1)完全有效阻止病毒侵染機體和細胞依靠一種多抗體機制,即需要靠針對不同靶抗原不同表位的抗體共同協作[22];因此靶向單個中和表位的抗體中和能力有限;2)由于VHH抗體體積和相對分子質量小,只有常規抗體的1/10左右,因此組織穿透能力強于常規抗體,在某些方面具有無可比擬的優勢;但是另一方面,小的體積和相對分子質量可能使VHH抗體結合到靶標后不足以形成足夠的空間位阻效應來封閉靶位點活性,從而導致中和效價低。為了增加VHH的中和活性,后續可嘗試將上述2株抗體多聚化,構建二價、三價等多價抗體以增強其活性。
[1]MERZ D C,SCHEID A,CHOPPIN P W.Importance of antibodies to fusion glycoprotein of paramyxoviruses in the prevention of spread of infection[J].JExpMed,1980,151(2):275-288.
[2]ZHANG R,PU J,SU JL,et al.Phylogenetic characterization of Newcastle disease virus isolated in the mainland of China during 2001-2009[J].VetMicrobiol,2010,141(3-4):246-257.
[3]LIEN Y Y,LEE J W,SU H Y,et al.Phylogenetic characterization of Newcastle disease viruses isolated in Taiwan during 2003-2006[J].VetMicrobiol,2007,123(1-3):194-202.
[4]DIEL D G,SUSTA L,CARDENAS GARCIA S,et al.Complete genome and clinicopathological characterization of a virulent Newcastle disease virus isolate from South America[J].JClinMicrobiol,2012,50(2):378-387.
[5]HU S L,MA H L,WU Y T,et al.A vaccine candidate of attenuated genotype VII Newcastle disease virus generated by reverse genetics[J].Vaccine,2009,27(6):904-910.
[6]XIAO S,NAYAK B,SAMUEL A,et al.Generation by reverse genetics of an effective,stable,live-attenuated Newcastle disease virus vaccine based on a currently circulation,highly virulent indonesian strain[J].PLoSOne,2012,7(12):e52751.
[7]CHEN S,HAO H,LIU Q,et al.Phylogenetic and pathogenic analyses of two virulent Newcastle disease viruses isolated from Crested Ibis (Nipponia nippon) in China[J].VirusGenes,2013,46(3):447-453.
[8]HAMERS-CASTERMAN C,ATARHOUCH T,MUYLDERMANS S,et al.Naturally occurring antibodies devoid of light chains[J].Nature,1993,363(6428):446-448.
[9]HOLLIGER P,HUDSON P J.Engineered antibody fragments and the rise of single domains[J].NatBiotechnol,2005,23(9):1126-1136.
[10]VANLANDSCHOOT P,STORTELERS C,BEIRNAERT E,et al.Nanobodies?:new ammunition to battle viruses[J].AntiviralRes,2011,92(3):389-407
[11]胡湘云,高小龍,付向晶,等.雙峰駝天然重鏈抗體可變區酵母雙雜交文庫的構建與鑒定[J].畜牧獸醫學報,2015,46(6):1071-1076.
HU X Y,GAO X L,FU X J,et al.Construction and characterization of a naiveCamelusBactrianusvariable domain of heavy chain of heavy-chain antibody(VHH)Yeast two-hybrid library[J].ActaVeterinariaetZootechnicaSinica,2015,46(6):1071-1076.(in Chinese)
[12]ZHAO Y,HAMMOND R W.Development of a candidate vaccine for Newcastle disease virus by epitope display in the Cucumber mosaic virus capsid protein[J].BiotechnolLett,2005,27(6):375-382.
[13]HARMSEN M M,RUULS R C,NIJMAN I J,et al.Llama heavy-chain V regions consist of at least four distinct subfamilies revealing novel sequence features[J].MolImmunol,2000,37(10):579-590.
[14]VAN DER LINDEN R,DE GEUS B,STOK W,et al.Induction of immune responses and molecular cloning of the heavy chain antibody repertoire of Lama glama[J].JImmunolMethods, 2000,240(1-2):185-195.
[15]VU K B,GHAHROUDI M A,WYNS L,et al.Comparison of llama VH sequences from conventional and heavy chain antibodies[J].MolImmunol,1997,34(16-17):1121-1131.
[16]RAHBARIZADEH F,RASAEE M J,FOROUZANDEH M,et al.Over expression of anti-MUC1 single-domain antibody fragments in the yeast Pichia pastoris[J].MolImmunol,2006,43(6):426-435.
[17]FU X J,GAO X L,HE S F,et al.Design and selection of a camelid single-chain antibody yeast two-hybrid library produced de novo for the cap protein of porcine circovirus type 2 (PCV2)[J].PLoSOne, 2013,8(3):556222.
[18]JONES D,CROWE E,STEVENS T A,et al.Functional and phylogenetic analysis of the ubiquitylation system in Caenorhabditis elegans:ubiquitin-conjugating enzymes,ubiquitin-activating enzymes,and ubiquitin-like proteins[J].GenomeBio,2002,3(1):RESEARCH0002.
[19]YANG S,SHANG Y,YIN S,et al.Selection and identification of single-domain antibody fragment against capsid protein of porcine circovirus type 2 (PCV2) from C.bactrianus[J].VetImmunolImmunopathol,2014,160(1-2):12-19.
[20]MALAKHOV M P,MATTERN M R,MALAKHOVA O A,et al.SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins[J].JStructFunctGenomics,2004,5(1-2):75-86.
[22]DELLA-PORTA A J,WESTAWAY E G.A multi-hit model for the neutralization of animal viruses[J].JGenVirol,1978,38(1):1-19.
(編輯白永平)
Screening and Characterization of VHH against Newcastle Disease Virus Fusion Protein
GAO Xiao-long△,HU Xiang-yun△,FU Xiang-jing,WANG Yan-ping,TONG Li-na,WANG Hai-xin,WANG Xing-long,ZHANG Shu-xia,XIAO Sa,DU En-qi*,YANG Zeng-qi*
(CollegeofVeterinaryMedicine,NorthwestA&FUniversity,Yangling712100,China)
This study aimed to obtain VHH (variable domains of camelid heavy-chain antibodies,VHH) against Newcastle disease virus (NDV) fusion protein (F protein).The synthesised DNA sequence encoding neutralizing epitope of F protein was used to construct bait plasmid.Then,the constructed bait was used to screen a naiveCamelusBactrianusVHH yeast two-hybrid library through yeast two-hybrid technology.After screening,four positive clones were primarily obtained and two of four VHHs with hallmark of VHH sequence were picked out for expression and purification.Subsequently,the biological activities of purified VHH were evaluated.Indirect ELISA results showed that the reactivity between VHH1 or VHH2 and NDV virion was significant higher than negative control.Western blot results suggested that VHH could specifically react with F protein.Neutralization assay further demonstrated that these two VHH could inhibit the replication of virulent NDV at some extent.These results indicate that we successfully obtained two VHH against NDV F protein,which laid the foundation for its application in ND control.
Newcastle disease virus;fusion protein;VHH;yeast two-hybrid
10.11843/j.issn.0366-6964.2016.08.015
2016-02-29
國家自然科學基金(31272578);西北農林科技大學引進人才科研啟動基金(Z111021006)
高小龍(1988-),男,陜西寶雞人,博士,主要從事動物傳染病和基因工程抗體研究,E-mail:gaoxiaolong1017@163.com;△共同第一作者
楊增岐,教授,E-mail:yzq8162@163.com
S852.659.5;S852.43
A
0366-6964(2016)08-1645-07
