長鏈非編碼RNA MALAT1的研究進展
2016-10-11 01:27:27宋鐵峰袁穎王會琴莊春雨王楠張同存
生物技術通報 2016年1期
關鍵詞:肺癌
宋鐵峰 袁穎 王會琴 莊春雨 王楠 張同存
(天津科技大學生物工程學院 分子藥理學和分子生物學研究室,天津 300457)
長鏈非編碼RNA MALAT1的研究進展
宋鐵峰 袁穎 王會琴 莊春雨 王楠 張同存
(天津科技大學生物工程學院 分子藥理學和分子生物學研究室,天津 300457)
長鏈非編碼RNA(Long noncoding RNAs,lncRNAs)是一類轉錄本長度超過 200 nt 的RNA分子,它們并不編碼蛋白質,而是以RNA的形式在轉錄水平、轉錄后水平、表觀遺傳學修飾等多種層面上調控基因的表達。肺癌轉移相關轉錄本1(Metastasis-associated lung adenocarcinoma transcript 1,MALAT1)首先在非小細胞肺癌被發現,并引起學者關注。MALAT1定位于染色體11q13.1,具有高度保守性。近年來,研究發現,MALAT1能特異性招募SR蛋白家族成員,并參與表觀遺傳調控以及細胞周期調控等;MALAT1高表達于多種腫瘤中,促進腫瘤細胞的增殖、轉移和侵襲。此外,最近發現MALAT1在血管生成過程中發揮重要作用。主要對MALAT1的特性、作用機制,以及在腫瘤和血管系統的作用作一系統綜述。
長鏈非編碼RNA;MALAT1;作用機制;腫瘤;血管生成
近年來,長鏈非編碼RNA(Long noncoding RNAs,lncRNAs)的研究進展迅猛,已成為分子生物學領域的最新研究熱點。 lncRNA是一類轉錄本長度超過 200 nt 的RNA分子,它們并不編碼蛋白質,而是以RNA的形式在多種層面上調控基因的表達水平[1]。LncRNA起初被認為是基因組轉錄的“噪音”,是RNA聚合酶Ⅱ轉錄的副產物,不具有生物學功能。但近年來的研究表明,lncRNA參與了表觀遺傳調控、轉錄調控以及轉錄后調控等多種重要的調控過程,與干細胞維持和分化、胚胎發育、細胞增殖與凋亡、細胞代謝、免疫調節等幾乎所有的生理或病理過程的調控密切相關,因此lncRNA的功能及其調控機制的闡明對現代生命科學具有重大的意義[2]。
肺癌轉移相關轉錄本1(Metastasis-associatedlung adenocarcinoma transcript 1,MALAT-1),又名核富集常染色體轉錄產物2(Nuclera-enriched autosomal transcript2,NEAT2)屬lncRNA家族重要成員,2003年在非小細胞肺癌(Non-small cell lung cancer,NSCLC)研究中被發現,近年來已逐步引起學者關注。MALAT1在多種腫瘤、組織中表達,已有研究表明,MALAT1能特異性招募SR蛋白家族成員、并參與表觀遺傳調控以及細胞周期調控等;MALAT1高表達于多種腫瘤中,能促進腫瘤細胞的增殖、轉移和侵襲等;MALAT1在血管生成過程中也發揮重要作用。本研究主要介紹MALAT1的發現、特性、并描述其已知的功能、作用機制和相互作用的分子,以及對近年來的研究作一系統綜述。
1 MALAT1的發現
2003年,Ji等[3]從初期非小細胞肺癌患者的腫瘤細胞中篩選出數個差異表達基因,其中包括含一個與肺癌轉移和預后相關的未知轉錄物,采用RACE法擴增到一個長約 940 nt 的片段,經檢索比對,該片段定位于人染色體 11q13,屬于長約 8.7 kb 的a基因。Ji等[3]根據其與NSCLC發生發展的關系,將a基因轉錄物重命名為肺癌轉移相關轉錄本1(MALAT1),后來也被稱作NEAT2。MALAT1缺乏有意義的開放性編碼框,在體外無法翻譯蛋白質,屬長鏈非編碼RNA。MALAT1是第一個與肺癌疾病相關聯的長鏈非編碼RNA。10多年前發現關于MALAT1的大量數據積累,包括MALAT1與其他癌癥細胞或疾病的關聯、對其生物合成的見解、與其他細胞的相互作用及分子機制等。
2 MALAT1的特性
2.1 MALAT1的結構特點
MALAT1編碼基因定位于染色體11q13.1,轉錄本序列長約 8 kb,屬于基因間轉錄本。在轉錄后水平,兩個內源RNA酶-RNase P 和 RNase Z可以修飾MALAT1[4]。初生的轉錄產物MALAT1在其多聚腺苷酸位點上游幾百堿基處可形成tRNA樣三葉草二級結構,此結構能被RNase P特異識別,RNase P結合三葉草結構后迅速在其最近上游端進行剪切,形成大小為 7 072 nt 的MALAT1成熟轉錄本和近3'端小轉錄本兩個片段。小轉錄本被轉運至胞漿,經RNase Z剪切加工,并在CCA添加酶的作用下,其3'端加上CCA臂后形成大小約為 61 nt 的胞漿成熟tRNA樣轉錄本[4],稱為MALAT1-associated small cytoplasmic RNA(mascRNA),隨后被運送到細胞質(圖1)[5,6]。研究發現,RNA轉錄后的修飾,如剪切、加帽等可大大增加RNA功能的多樣性,但mascRNA的功能迄今未知[5]。因此,目前在功能上無法將mascRNA與MALAT1聯系起來。
MALAT1的5'端沒有帽子結構,不參與蛋白質的翻譯,3'端沒有ploy(A)尾,但其是一個非常穩定的lncRNA,最新研究發現,可能是由于在3'末端形成了三螺旋結構[8,9]。Wilusz等[8]研究發現MALAT1 3'末端形成的三螺旋結構可保護MALAT1 3' 端免受 3'-5' 核酸外切酶的剪切,從而維持MALAT1的完整性。
2.2 MALAT1的核定位
LncRNA的功能在很大程度上決定于亞細胞結構定位的特性。與MALAT1同一家族的成員,核富集常染色體轉錄物1(Nuclera-enriched autosomal transcript,NEAT1)參與核內的超核小斑結構的組裝和結構維持[10]。與NEAT1相似,MALAT1基因特異性定位于細胞核核小體核斑(Nuclearspeckles)區,在該區進行轉錄,但其在該核斑區富集是以RNA聚合酶Ⅱ依賴的轉錄被激活為前提[11]。進一步研究發現,干擾MALAT1不影響某些核斑的表達、定位,因此MALAT1可能不是組成核斑的結構成分[12]。同時,MALAT1序列本身還存在兩個必需的獨立結構域指導其定位于核斑,分別位于MALAT1序列的 1 961-3 040 nt和 6 387-7 011 nt,這兩個區域中的任何一個基因突變都導致MALAT1定位失常,游離到細胞質中,失去功能[13]。例如,用siRNA干擾MALAT1后,MALAT1分散到核質中,失去調節基因表達的功能。
2.3 MALAT1的高度保守性
MALAT1在哺乳動物進化中高度保守,核苷酸序列3' 末端 5 kb 左右人鼠同源性高達90%,這都提示其在進化過程中MALAT1扮演了重要角色。MALAT1的全序列存在 4 個人鼠高度同源區域,提示這些區域極可能為MALAT1的功能性區域。楊敏慧等[14]將人鼠同源序列集中區域(3 207-8 441 bp,覆蓋4個人鼠高同源區),即MALAT1的功能性區域作為研究對象,構建真核重組表達載體,并轉染人大腸癌細胞株SW620,為進一步研究MALAT1的結構、功能及其與人大腸癌的關聯奠定基礎。
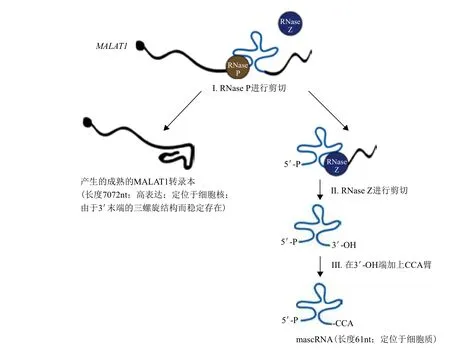
圖1 MALAT1 的結構剪切[7]
3 MALAT1相關作用機制
3.1 MALAT1調節剪接
已知mRNA前體加工過程中,核內核小斑的絲氨酸/精氨酸富集蛋白(Serine/arginie riched protein,SR protein)家族成員具有重要作用。SR蛋白的分子結構包含N末端的RNA結構域和C末端RS結構域,N末端的RNA結構域可以與mRNA前體相互作用。SR蛋白從核小斑轉移到轉錄活化部位,結合mRNA前體,并招募多種剪接因子,介導mRNA前體加工[15,16]。Tripathi等[17]通過RNA-FISH實驗,使用MALAT1和U2-snRNA特異性探針和SR剪接因子抗體,證實核富集的MALAT1與絲氨酸精氨酸SR蛋白相互作用,影響剪接因子的配送等。沉默MALAT1或過表達SR蛋白可以不同程度的改變內源性前mRNA的選擇性剪接。磷酸化是決定SR蛋白功能活化的重要步驟,研究還發現,MALAT1在細胞水平調節SR蛋白質的磷酸化和去磷酸化水平,從而控制其活性。Wang等[18]發現,在GC細胞系,過表達SF2/ASF的核仁中,MALAT1異常高表達,當下調MALAT1表達時,SF2/ASF的核分布和表達顯著受損,MALAT1可調控SF2/ASF的核內定位。
3.2 MALAT1與表觀遺傳
多梳蛋白2(Polycomb 2,PC2)是PcG蛋白家族的一員,是一種重要的基因轉錄抑制因子。Pc2蛋白甲基化后聚集在轉錄因子E2F1的靶基因啟動子區,與轉錄本TUG1結合定位于亞核結構多梳體(Polycomb body,PcG body),此結構含有多種轉錄抑制因子,抑制E2F1靶基因的轉錄[19]。絲裂因子刺激后,甲基化的Pc2去甲基,轉而與MALAT-1結合,共同作為一種分子支架,特異性定位于亞核結構染色質間顆粒(Interchromatin granule,ICG)中[20]。MALAT-1作為停泊位點與去甲基的Pc2結合后,不僅促進生長基因從PcG重新定位到ICG,而且還通過組裝多種共調節復合體,與多種基因激活相關蛋白相互作用,促進基因表達。與此同時,Pc2作為組蛋白的閱讀器[21],MALAT-1與之結合后可改變Pc2對組蛋白的識別能力,從識別組蛋白H3K9me3轉換為閱讀組蛋白H2AK5ac和組蛋白H2AK13ac,而這兩種蛋白是基因激活的標志物。
3.3 MALAT1與細胞周期
最近的研究表明在哺乳動物細胞中幾個lncRNAs可以調節細胞周期的進程。研究表明,在正常細胞周期進程中,MALAT1參與調節。Tripathi等[22]對人二倍體成纖維細胞進行了全基因組和轉錄組學分析,證實MALAT1調控細胞周期相關基因的表達,并且它是有絲分裂進程所必須的基因。MALAT1的沉默導致p53及其靶基因的活化,同時細胞周期進程取決于p53的表達變化,這暗示著p53是MALAT1的主要下游介質。此外,在MALAT1沉默的細胞中,B-MYB(MybrelatedproteinB)表達降低,B-MYB是一個參與G2/M期的進程的致癌轉錄因子,MALAT1改變了B-MYB 前體mRNA上的剪接結合因子結合位點和它的異常剪接。在人類成纖維細胞中,MALAT1通過調節細胞周期相關轉錄因子的表達促進細胞增殖。Yang等[23]發現,在G2 / M期,MALAT1的轉錄物由細胞核移位進入細胞質。調查還發現,在此過程中,MALAT1與核糖核蛋白C(Heterogeneous nuclear ribonucleoprotein C,hnRNP C)相互作用。利用功能缺失實驗,MALAT1的表達下調抑制了hnRNP C在G2/M期的轉錄,并導致G2/ M期阻滯。這些發現提供了有關MALAT1在調節細胞增殖作用機理的見解。
4 MALAT1與腫瘤
MALAT1廣泛表達于哺乳動物正常組織,并在諸多人類惡性腫瘤中異常表達,其異常表達改變了腫瘤細胞的生物學表型。雖然MALAT1不直接編碼蛋白,但其對腫瘤的增殖、凋亡、侵襲轉移、耐藥都有不同程度的影響,發揮重要作用。有關MALAT1在多種腫瘤中的調節和功能匯總于表1。
4.1 MALAT1與肺癌
Tano等[24]為了研究MALAT1是否與肺癌的轉移有關,用siRNA干擾了MALAT1的表達發現,細胞的運動性受損,同時也影響眾多基因的表達,其中包括CTHRC1、CCT4、HMMR或ROD1基因,降低受MALAT1調控的這些基因的表達都明顯抑制細胞遷移。因此,MALAT1是一種通過從轉錄和轉錄后水平調控運動相關基因表達促進細胞運動的非編碼RNA。Schmidt等[25]研究發現,MALAT1在人類的非小細胞肺癌細胞系中表達明顯高于癌旁正常組織。并在非小細胞肺癌A549中,用siRNA介導的方法干擾MALAT1,抑制了細胞的遷移以及裸鼠中腫瘤的形成,而在NIH 3T3細胞中過表達MALAT1,顯著增加細胞遷移,說明MALAT1具有促進肺癌形成的能力。Schmidt等[25]還對非小細胞肺癌組織進行原位雜交檢測,揭露鱗狀細胞肺癌中,MALAT1的高表達與預后不良有關。這表明MALAT1有望成為肺癌預后的新指標。Gutschner等[26]發現,在肺癌細胞中,MALAT1不會改變剪接,而是積極調節基因表達,包括一組轉移相關基因。Weber等[27]研究證明,可以根據人外周血細胞中MALAT1表達的差異區分癌癥患者和正常志愿者。因此,MALAT1可作為診斷非小細胞肺癌的血液生物標志物的候選者。Shen等[28]還研究了在 78 例非小細胞肺癌中,與非腦轉移組織相比,肺癌腦轉移組織MALAT1的水平顯著升高。由此可見,MALAT1的表達水平與患者生存相關聯。同時,沉默MALAT1抑制腦轉移肺癌細胞侵襲和轉移。因此MALAT1 非常有希望成為治療肺癌的腦轉移的靶標。
4.2 MALAT1與宮頸癌
Guo等[29]發現在人屬宮頸癌細胞系CaSki中沉默MALAT1的表達,可以抑制細胞的增殖和遷移。對細胞周期分析發現,干擾MALAT1后,G1期細胞明顯增多,S期細胞減少,說明MALAT1可以促進細胞從G1期到S期的轉換,從而加速細胞增殖。進一步研究發現,在干擾MALAT1時,誘導了凋亡基因caspase-3、caspase-8和Bax的表達,而降低了Bcl-2和Bcl-xL的表達。說明MALAT1參與調控宮頸癌細胞生長、增殖和凋亡。Jiang等[30]研究證明,宮頸癌細胞系與正常宮頸鱗狀細胞樣品進行比較,MALAT1表達上調。亦證明MALAT1能夠促進細胞的遷移和增殖,值得注意的是,MALAT1的表達下降與HPV16 E6/ E7的敲低有關,這表明,HPV是導致MALAT1在宮頸癌鱗狀細胞癌中活化的重要因素之一。

表1 MALAT1在多種腫瘤中的作用
4.3 MALAT1與肝癌
Luo等[31]根據轉錄分析,發現MALAT1過度表達在肝母細胞瘤HPBL,肝細胞癌HCC,以及相鄰的肝硬化組織。Lin等[32]發現在肝癌組織中,MALAT1的表達較癌旁組織高出 6 倍,并且MALAT1可能成為肝癌診斷的潛在分子標志。Lai等[33]收集包括 9 種不同細胞系,其中1種正常的肝細胞系L02和8種不同的肝癌細胞系(HepG2、Hep3B、SMMC7721、HUH7、MHCC97L、MHCC97H、HCCLM3和SKhep1),同時收集了112例HCC病例樣品(其中有 60 例接受了肝移植的病人),并用qPCR檢測了MALAT1的表達量發現,高表達MALAT1顯著增加了肝移植病人肝癌復發的風險。多因素分析,MALAT1是預測肝癌復發的獨立預后因素,可作為一種新的標志物用于預測肝移植后肝癌的復發。此外,在HepG2細胞中抑制MALAT1的表達,能有效降低細胞活力、運動、侵襲,并增加細胞凋亡的敏感性。Wang等[34]發現癌蛋白相關蛋白(YAP)可以通過對絲氨酸/富含精氨酸的剪接因子1(Recombinant serine/arginine-rich splicing factor 1,SRSF1)發揮抑制作用,進而從轉錄和轉錄后水平上調MALAT1的表達。過表達的YAP通過與血管動蛋白(Human angiomotin,AMOT)的相互作用,使核內的SRSF1受損。而SRSF1本身對MALAT1有拮抗作用,通過YAP對SRSF1的抑制,使這種拮抗作用解除,從而上調MALAT1的表達。Wang等還發現,過表達YAP與同時降低SRSF1的表達將導致細胞移動性顯著增強,腫瘤快速增殖。總之,在SRSF1、YAP和MALAT1之間,這些數據提供了一個新穎的平衡機制,但關于YAP調節長鏈非編碼RNA的具體機制尚不清楚。
4.4 MALAT1與膀胱癌
Ying等[35]通過qPCR檢測了膀胱癌中MALAT1的含量,證實了MALAT1在膀胱癌組織的表達高于癌旁組織。用siRNA干擾MALAT1,膀胱癌細胞的遷移能力下降。MALAT1表達下調后,與EMT相關的鋅指E盒結合同源盒蛋白1(Zinc-finger E-boxbinding homeobox 1,ZEB1)、ZEB2和鋅指轉錄因子Slug等蛋白表達降低,與細胞黏附相關的鈣黏連蛋白E-cadherin表達增加,同時Wnt信號通路中的關鍵分子β-連環蛋白也減少。Ying等推測,膀胱癌中高表達的MALAT1可能通過激活Wnt信號通路促進癌細胞的上皮-間質轉化(Epithelial-to-mesenchymal transition,EMT)來促進癌細胞轉移。2014年,Fan等[36]又發現MALAT1可與抑制劑zeste 12結合,而這種結合會導致E-cadherin和N-caherind表達增加。Han等[37]檢測了 36 例膀胱尿路上皮癌組織,證實膀胱尿路上皮癌與正常上皮相比較,MALAT1的表達上調。同時在膀胱尿路上皮癌T24和5637細胞中,干擾MALAT1的表達,抑制細胞增殖,誘導細胞凋亡,降低細胞運動性。到目前為止,盡管人們對lncRNA的認識還不夠全面,但是人們發現lncRNA可與microRNA相互調控,從而對腫瘤發揮作用。Han等[38]發現hsa-miR-125b可以通過下調MALAT1的表達而抑制膀胱癌的發展。
4.5 MALAT1與食管鱗狀細胞癌
MALAT1已被證明是各種人類惡性腫瘤的重要參與者,但其在食管鱗狀細胞癌的作用仍知之甚少。Hu等[39]通過qPCR分析了臨床組織中MALAT1的含量發現,MALAT1過度表達于食管鱗狀細胞癌。在體內和體外,沉默MALAT1抑制了腫瘤的增殖,以及細胞侵襲和遷移能力。MALAT1的沉默也使細胞周期阻滯在G2/M期,誘導細胞凋亡。同時,Western Blot結果顯示,MALAT1的沉默使與阻滯G2 / M期相關聯的ATM-CHK2通路磷酸化。數據表明,MALAT1作為食管癌的癌基因,它通過作用于ATMCHK2通路調節食管癌的增長,具體調節機制尚不清楚。Wang等[40]發現在食管癌細胞中,miR-101和miR-217轉錄后調控MALAT1沉默,MALAT1這種轉錄后沉默可顯著抑制食管癌細胞的增殖,通過阻滯阻滯G2 / M期,這可能是由于MALAT1介導的上調p21和p27表達和下調B-MYB表達的作用。
4.6 MALAT1與其他腫瘤
在其他腫瘤中,如乳腺癌中,用siRNA干擾MALAT1后,可以觀察到細胞遷移受到抑制,這說明MALAT1在乳腺癌中起到促進遷移的作用[41]。在胃癌SGC-7901細胞中,干擾MALAT1,可以觀察到細胞阻滯在G0/G1期,同時也伴隨著細胞增殖的顯著抑制[42]。在結腸直腸癌細胞中,干擾MALAT1,可促進腫瘤的生長和轉移,此外PRKA激酶錨蛋白9(A kinase anchor protein 9,AKAP-9)在mRNA和蛋白質水平顯著上調,而AKAP-9的敲除阻斷MALAT1介導的癌細胞的增殖、遷移和侵襲,證明MALAT1可調節靶蛋白AKAP-9促進結直腸癌增殖、遷移和侵襲[43]。在人骨肉瘤細胞中,Dong等[44]發現在體內和體外,MALAT1的沉默顯著抑制細胞的增殖和侵襲和轉移,推測MALAT1可能通過抑制PI3K / AKT信號通路而促進腫瘤的生長和轉移,總之,MALAT1可以視為人骨肉瘤的治療靶點。MALAT1在許多腫瘤中都高表達,其功能失調普遍存在于人類多種惡性腫瘤中,說明了MALAT1與腫瘤具有重要關系。
5 MALAT1對血管內皮細胞的作用
最近研究發現MALAT1也參與調控內皮細胞的血管生成。已有關于microRNA調控血管內皮細胞功能、血管生長和重塑的報道,但人們對lncRNAs在內皮細胞中的功能知之甚少[45]。
Michalik等[46]發現了長鏈非編碼RNA在人臍靜脈內皮細胞HUVEC中的高表達特征,界定了在體內和體外,MALAT1 對內皮細胞和血管生成的功能性作用。在體外,通過siRNA干擾MALAT1的表達,可增加內皮細胞基礎出芽和促進細胞遷移,反之內皮細胞的出芽也會受到抑制。在siRNA干擾MALAT1表達的基礎上,進一步用VEGF刺激血管內皮細胞,可誘導內皮細胞不連續的出芽生長,并影響細胞周期進程,使細胞分裂期S期減少。同時,利用廣泛性抑制長鏈非編碼RNA的抑制劑——GapmeR抑制MALAT1的表達,也出現了促進內皮細胞出芽和遷移的現象,加入VEGF后,也誘導內皮細胞的不連續出芽、影響細胞周期進程。由此證實了在體外,MALAT1對內皮細胞出芽、遷移和增殖都有作用。在體內,Michalik等利用植物凝集素-B4對出生5 d小鼠的視網膜進行染色,可以使視網膜可視化。在敲除MALAT1的小鼠的視網膜切片中可觀察到視網膜的血管叢密度降低,并觀察到視網膜血管內皮細胞的增殖受到抑制。在小鼠的單側下肢缺血模型中,利用GapmeRs抑制MALAT1可減少血流的恢復和后肢局部缺血后毛細血管的密度。由此證實了,在體內,沉默MALAT1可減少血管內皮細胞增殖、抑制新生血管化、減少后肢局部缺血后的血流恢復和毛細血管密度。為了研究MALAT1對內皮細胞增殖的作用,利用qRT-PCR檢測干擾MALAT1表達后的細胞周期相關基因表達,結果顯示MALAT1的沉默調控各種細胞周期調控因子的表達,分裂期S期的調控蛋白CCNA2、CCNB1、CCNB2等表達下調,細胞周期抑制劑因子p21、p27Kip1等表達上調,進一步證實MALAT1促進內皮細胞增殖。
治療性血管新生(Therapeutic angiogenesis)是一種有應用前景的治療策略,以促進缺血組織的再生[46]。lncRNAs已經描述了在內皮細胞出芽過程中起關鍵作用,但對于在血管生成lncRNAs的作用還沒有被研究[45]。lncRNA MALAT1高度表達于內皮細胞,它是由缺氧誘導,并是內皮細胞增殖的需要。GapmeRs 的沉默或MALAT1基因缺失減少小鼠中新生血管形成。因此,增強MALAT1是可以促進血管新生治療一個潛在的戰略。
Puthanveetil等[47]用含有不同葡萄糖水平的培養基培養人的臍靜脈內皮細胞,12 h后發現高葡萄糖水平會引起MALAT1的表達增加。隨后又證明MALAT1的增加與血清中淀粉樣蛋白抗原3(SAA3)的增加相關,而SAA3的增加進一步伴隨著腫瘤壞死因子α(Tumor necrosis factor-α,TNF-α)和白細胞介素6(Interleukin-6,IL-6)的mRNA和蛋白水平增加。這項研究的結果表明,MALAT1通過活化淀粉樣蛋白抗原3(Serum amyloid A3,SAA3)而上調葡萄糖誘導的炎性介質TNF-α和IL-6。這些關于MALAT1的新的調節機制的發現對于糖尿病微觀和宏觀血管并發癥的研究具有重要意義。
Liu等[48]發現沉默MALAT1能明顯改善糖尿病患者的視網膜病變(Diabetic retinopathy,DR),如周細胞損失、毛細管變性、微血管滲漏和視網膜炎癥。此外,沉默MALAT1可以調節視網膜血管內皮細胞增殖、遷移和體外血管的生成。研究表示MALAT1的低表達可作為糖尿病相關的微血管并發癥的標志。
6 展望
近年來,隨著研究的深入,更多的lncRNA被發現,更多的疾病與lncRNA的異常表達有關。MALAT1作為其中的一員,具有高度保守性,定位于細胞核,參與基因的剪接,并且通過表觀遺傳控制基因的表達。MALAT1高度表達在腫瘤細胞中,其功能失調普遍存在于多種惡性腫瘤中,對腫瘤的增殖、凋亡、侵襲轉移都有不同程度的影響,發揮重要作用,但是其在腫瘤中的生物特性、作用途徑、作用靶點、調節機制仍未完全研究清楚,還需進行深入研究和探討。例如,目前普遍采用干擾MALAT1的方式,也可過表達MALAT1,MALAT1功能的過表達研究可以有助于確定相關的潛在分子機制。除了腫瘤細胞,還可探索MALAT1在其他細胞中的作用,如心肌細胞、平滑肌細胞、炎癥細胞或心臟成纖維細胞等。相信通過體內外的深入研究將會在更廣的層面上揭示MALAT1的功能及調控機制,為人類創造更多新的突破口。
[1] Ponting CP, Belgard TG. Transcribed dark matter:meaning or myth?[J]. Hum Mol Genet, 2010, 19(R2):R162-168.
[2] Gutschner T, Diederichs S. The hallmarks of cancer:a long noncoding RNA point of view[J]. RNA Biol, 2012, 9:703-719.
[3] Ji P, Diederichs S, Wang W, et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in earlystage non-small cell lung cancer[J]. Oncogene, 2003, 22(39):8031-8041.
[4] Kuhn CD, Wilusz JE, Zheng Y, et al. On-enzyme refolding permits small RNA and tRNA surveillance by the CCA-adding enzyme[J]. Cell, 2015, 160(4):644-658.
[5] Wilusz JE, Freier SM, Spector DL, et al. 3' end processing of a long nuclear-retained noncoding RNA yields a tRNA-like cytoplasmic RNA[J]. Cell, 2008, 135(5):919-932.
[6] Fejes-Toth K, Sotirova V, Sachidanandam R, et al. Posttranscriptional processing generates a diversity of 5'- modified long and short RNAs[J]. Nature, 2009, 457(7232):1028-1032.
[7] Gutschner T, Hammerle M, Diederichs S, et al. MALAT1—a paradigm for long noncoding RNA fuction in cancer[J]. J Mol Med(Berl), 2013, 91(7):791-801.
[8] Wilusz JE, Jnbaptiste CK, Lu LY, et al. A triple helix stabilizes the 3' ends of long noncoding RNAs that lack poly(A)tails[J]. Genes Dev, 2012, 26(21):2392-2407.
[9] Brown JA, Valenstein ML, Yario TA, et al. Formation of triplehelical structures by the 3'-end sequences of MALAT1 and MENbeta noncoding RNAs[J]. Proc Natl Acad Sci USA, 2012, 109(47):19202-19207.
[10] Wilusz JE, Sunwoo H, Spector DL, et al. Long noncoding RNAs:functional surprises from the RNA world[J]. Genes Dev, 2009,23(13):1494-1504.
[11] Clemson CM, Hutchinson JN, Sara SA, et al. An architectural role for a nuclear noncoding RNA:NEAT1 RNA is essential for the structure of paraspeckles[J]. Mol Cell, 2009, 33(6):717-726.
[12] Bernard D, Prasanth KV, Tripathi V, et al. A long nuclear-retained non-coding RNA regulates synaptogenesis by modulating gene expression[J]. EMBO J, 2010, 29(18):3082-3093.
[13] Miyagawa R, Tano K, Mizuno R, et al. Identification of cis- and trans-acting factors involved in the localization of MALAT-1 noncoding RNA to nuclear speckles[J]. RNA, 2012, 18(4):738-751.
[14] 楊敏慧, 王雙雙, 王曉燕, 等. 非編碼基因MALAT1功能性序列真核表達載體的構建及其在SW620細胞中的表達[J]. 熱帶醫學雜志, 2010, 10(3):248-257.
[15] Mukherjee B, Tomimatsu N, Amancherla K, et al. The dual PI3K/ mTOR inhibitor NVP-BEZ235 is a potent inhibitor of ATM- and DNA-PKCs-mediated DNA damage responses[J]. Neoplasia,2012, 14(1):34-43.
[16] Graveley BR. Sorting out the complexity of SR protein functions[J]. RNA, 2000, 6(9):1197-1211.
[17] Tripathi V, Ellis JD, Shen Z, et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation[J]. Mol Cell, 2010, 39(6):925-938.
[18] Wang JQ, Su LP, Chen XH, et al. MALAT1 promotes cell proliferation in gastric cancer by recruiting SF2/ASF[J]. Biomed Pharmacother, 2014, 68(5):557-564.
[19] Bantignies F, Roure V, Comet I, et al. Polycomb- dependent regulatory contacts between distant Hox loci in Drosophila[J]. Cell, 2011, 144(2):214-222.
[20] Spector DL, Lamond AI. Nuclear speckles[J]. Cold Spring Harb Perspect Biol, 2010,(2):a005546.
[21] Bernstein E, Duncan EM, Masui O, et al. Mouse polycomb proteins bind differentially to methylated histone H3 and RNA and are enriched in facultative heterochromatin[J]. Mol Cell Biol, 2006,26(7):2560-2569.
[22] Tripathi V, Shen Z, Chakraborty A, et al. Long noncoding RNA MALAT1 controls cell cycle progression by regulating the expression of oncogenic transcription factor B-MYB[J]. PLoS Genet, 2013, 9(3):e1003368.
[23] Yang F, Yi F, Han X, et al. MALAT-1interacts with hnRNP C in cell cycle regulation[J]. FEBS Lett, 2013, 587(19):3175-3181.
[24] Tano K, Mizuno R, Okada T, et al. MALAT-1 enhances cell motility of lung adenocarinoma cells by influencing the expression of motility-related genens[J]. FEBS Lett, 2010, 584(22):4575-4580.
[25] Schmidt LH, Spieker T, Koschmieder S, et al. The long noncoding MALAT-1 RNA indicates a poor prognosis in non-small cell lung cancer and induces migration and tumor growth[J]. J Thorac Oncol, 2011, 6(12):1984-1992.
[26] Gutscher T, H?mmerle M, Eissmann M, et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells[J]. Cancer Res, 2013, 73(3):1180-1189.
[27] Weber DG, Johnen G, Casjens S, et al. Evaluation of long noncoding RNA MALAT1 as a candidate blood-based biomarker for the diagnosis of non-small cell lung cancer[J]. BMC Res Notes, 2013, 6:518.
[28] Shen L, Chen L, Wang Y, et al. Long noncoding RNA MALAT1 promotes brain metastasis by inducing epithelial-mesenchymal transition in lung cancer[J]. J Neurooncol, 2015, 121(1):101-108.
[29] Guo F, Li Y, Liu Y, et al. Inhibition of metastasis-associated lung adenocarinoma transcript 1 in CaSki human cervical cancer cells suppresses cell proliferation and invasion[J]. Acta Biochim Biophys Sin(Shanghai), 2010, 42(3):224-229.
[30] Jiang Y, Li Y, Fang S, et al. The role of MALAT1 correlates with HPVin cervical cancer[J]. Oncol Lett, 2014, 7:2135-2141.
[31] Luo JH, Ren B, Keryanov S, et al. Transcriptomic and genomic analysis of human hepatocellular carcinomas and hepatoblastomas[J]. Hepatology, 2006, 44(4):1012-1024.
[32] Lin R, Maeda S, Liu C, et al. A large noncoding RNA is a marker for murine hepatocellular carcinomas and a spectrum of human carcinomas[J]. Oncogene, 2007, 26(6):851-858.
[33] Lai M C, Yang Z, Zhou L, et al. Long non-coding RNA MALAT1 overexpression predicts tumor recurrence of hepatocellular carcinoma after liver transplantation[J]. Med Oncol, 2012, 29(3):1810-1816.
[34] Wang JY, Wang HM, Zhang Y, et al. Mutual inhibition between YAP and SRSF1 maintains long non-coding RNA, MALAT1-induced tumourigenesis in liver cancer[J]. Cellular Signalling,2014, 26(5):1048-1059.
[35] Ying L, Chen Q, Wang Y, et al. Upregulated MALAT-1 contributes to bladder cancer cell migration by inducing epithelial-tomesenchymal transition[J]. Mol Biosyst, 2012, 8:2289-2294.
[36] Fan Y, Shen B, Tan M, et al. TGF-β-induced upregulation of malat1 promotes bladder cancer metastasis by associating with suz12[J]. Clin Cancer Res, 2014, 20(6):1531-1541.
[37] Han Y, Liu Y, Nie L, et al. Inducing cell proliferation inhibition,apoptosis, and motility reduction by silencing long noncoding ribonucleic acid metastasis-associated lung adenocarcinoma transcript 1 in urothelial carcinoma of the bladder[J]. Urology,2013, 81(1):209. e1-7.
[38] Han Y, Liu Y, Zhang H, et al. Hsa-miR-125b suppresses bladder cancer development by down-regulating oncogene SIRT7 and oncogenic long nocoding RNA MALAT1[J]. FEBS Lett, 2013,587(23):3875-3882.
[39] Hu L, Wu Y, Tan D, et al. Up-regulation of long noncoding RNA MALAT1 contributes to proliferation and metastasis in esophageal squamous cell carcinoma[J]. J Exp Clin Cancer Res, 2015, 34:7.
[40] Wang X, Li M, Wang Z, et al. Silencing of long noncoding RNA MALAT1 by miR-101 and miR-217 inhibits proliferation,migration, and invasion of esophageal squamous cell carcinoma cells[J]. J Biol Chem, 2015, 290(7):3925-3935.
[41] Zhao Z, Chen C, Liu Y, et al. 17β-Estradiol treatment inhibits breast cell proliferation, migration and invasion by decreasing MALAT-1RNA level[J]. Biochem Biophys Res Commun, 2014, 445(2):388-393.
[42] Wang J, Su L, Chen X, et al. MALAT1 promotes cell proliferation in gastric cancer by recruiting SF2/ASF[J]. Biomed Pharmacother,2014, 68(5):575-564.
[43] Yang MH, Hu ZY, Xu C, et al. MALAT1promotes colorectal cancer cell proliferation/migration/invasion via PRKA kinase anchor protein 9[J]. Biochim Biophys Acta, 2015, 1852:166-174.
[44] Dong Y, Liang G, Yuan B, et al. MALAT1 promotes the proliferation and metastasis of osteosarcoma cells by activating the P13K/Akt pathway[J]. Tumour Biol, 2015, 36(3):1477-1486.
[45] Thum T, Fiedler J. LINCing MALAT1 and angiogenesis[J]. Circulation Research, 2014, 114(9):1366-1368.
[46] Michalik KM, You X, Manavski Y, et al. Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth[J]. Circulation Research, 2014, 114(9):1389-1397.
[47] Puthanveetil P,Chen S,Feng B,et al. Long non-coding RNA MALAT1 regulates hyperglycaemia induced inflammatory process in the endothelial cells[J]. J Cell Mol Med,2015,DOI:10. 1111/jcmm. 12576.
[48] Liu JY,Yao J,Li XM,et al. Pathogenic role of lncRNAMALAT1 in endothelial cell dysfunction in diabetes mellitus[J]. Cell Death Dis,2014,e1506. DOI:10.1038/cddis. 2014. 466.
[49] Wu XS, Wang XA, Wu WG, et al. MALAT1 promotes the proliferation and metastasis of gallbladder cancer cells by activating the ERK/MAPK pathway[J]. Cancer Biol Ther, 2014, 15(6):806-814.
[50] Ma KX, Wang HJ, Li XR, et al. Long noncoding RNA MALAT1 associates with the malignant atatus and poor prognosis in glioma[J]. Tumour Biol, 2015, DOI:10.1007/s13277-014-2969-7.
[51] Pang EJ, Yang R, Fu XB, et al. Overexpression of long noncoding RNA MALAT1 is correlated with clinical progression and unfavorable prognosis in pancreatic cancer[J]. Tumour Biol,2015, 36(4):2403-2407.
(責任編輯狄艷紅)
Research Progress on a Long Non-coding RNA MALAT1
SONG Tie-feng YUAN Ying WANG Hui-qin ZHUANG Chun-yu WANG Nan ZHANG Tong-cun
(Laboratory of Molecular Biology and Molecular Pharmacology,Tianjin University of Science and Technology,Tianjin 300457)
Long non-coding RNAs(lncRNAs)are the ones with transcripts that are longer than 200 nt. They do not encode proteins,but regulate the expression levels of the gene as a RNA molecular at transcriptional, post-transcriptional and epigenetic modification. Metastasisassociated lung adenocarcinoma transcript 1(MALAT1)was firstly discovered in non-small cell lung cancer, and has caused the concerns from scholars. MALAT1 located on chromosome 11q13.1 and was highly conserved. In recent years, studies have shown that MALAT1 specifically recruited SR protein family members, was involved in epigenetic regulation and cell cycle regulation. MALAT1, which is highly expressed in many human tumors, promotes tumor proliferation, invasion and metastasis. In addition, recently it is found that MALAT1 plays an important role in angiogenesis. This article systematically reviewed the characteristics and mechanism of MALAT1, as well as its biological function in tumor and vascular systems.
lncRNA;MALAT1;mechanism;tumor;angiogenesis
10.13560/j.cnki.biotech.bull.1985.2016.01.005
2015-04-30
國家自然科學基金項目(31470816,31401170,31171297,31270837,31171303)
宋鐵峰,女,碩士,研究方向:長鏈非編碼RNA在平滑肌中作用的初步探究;E-mail:songtiefeng1025@126.com
王楠,女,教授,研究方向:轉錄因子和表觀遺傳修飾在干細胞分化中的調控作用;E-mail:wn929@tust.edu.cn張同存,男,教授,研究方向:心血管疾病和腫瘤等重大疾病的轉錄調控機制;E-mail:tony@tust.edu.cn
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
中國藥學藥品知識倉庫(2022年1期)2022-03-23 04:16:57
昆明醫科大學學報(2021年4期)2021-07-23 01:21:44
天津醫科大學學報(2021年2期)2021-03-29 05:30:38
天津醫科大學學報(2019年6期)2019-08-13 07:04:26
癌變·畸變·突變(2016年3期)2016-02-27 06:15:34
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國當代醫藥(2015年7期)2015-03-01 02:01:19
鄭州大學學報(醫學版)(2015年1期)2015-02-27 14:50:26
