TALENs新型模塊組裝法及活性鑒定方法
2016-10-11 01:27:53高敬魏迪池振奮張癸榮張萬明聶凌云
生物技術(shù)通報(bào) 2016年1期
高敬魏迪池振奮張癸榮張萬明聶凌云
(1.河北北方學(xué)院,張家口 075000;2.中國人民解放軍總后勤部衛(wèi)生部藥品儀器檢驗(yàn)所,北京 100166;3.中國科學(xué)院北京基因組研究所,北京 100101)
TALENs新型模塊組裝法及活性鑒定方法
高敬1,2魏迪1,2池振奮3張癸榮1,2張萬明1聶凌云1,2
(1.河北北方學(xué)院,張家口 075000;2.中國人民解放軍總后勤部衛(wèi)生部藥品儀器檢驗(yàn)所,北京 100166;3.中國科學(xué)院北京基因組研究所,北京 100101)
旨在提供一種新型的轉(zhuǎn)錄激活樣效應(yīng)子核酸酶(Transcription activator-like effector nucleases,TALENs)模塊組裝法(Unit assembly,UA)及活性鑒定方法。利用TALE-NT 2.0在線工具在線粒體DNA(Mitochondrial DNA,mtDNA)設(shè)計(jì)TALENs識(shí)別位點(diǎn)和剪切位點(diǎn),借助pEGFP-N1質(zhì)粒將該段靶序列隨機(jī)整合于HEK293F核基因組中,構(gòu)建HEK293F-T1細(xì)胞系,用于TALENs活性鑒定。依據(jù)轉(zhuǎn)錄激活樣效應(yīng)子(Transcription activator-like effectors,TALEs)自然單元模塊序列,設(shè)計(jì)出一種新型的人工TALEs偏單元;根據(jù)TALENs識(shí)別位點(diǎn),選取相應(yīng)一單元模塊,利用UA法組裝;并設(shè)計(jì)出含有相應(yīng)雙酶切位點(diǎn)的TALEs串聯(lián)模塊序列和TALENs載體序列,定向連接后瞬時(shí)轉(zhuǎn)染HEK293F-T1細(xì)胞系。結(jié)果顯示,新型模塊組裝法定向組裝TALEs模塊,克隆效率高且瞬時(shí)轉(zhuǎn)染后可看到清晰的套峰。結(jié)果表明,新型模塊組裝方法提高了TALENs構(gòu)建過程中的克隆效率,并且不受末位0.5模塊的RVD(repeat-variable diresidue)限制,增加了靶序列設(shè)計(jì)的靈活性。
轉(zhuǎn)錄激活樣效應(yīng)子核酸酶;模塊組裝法;線粒體DNA;HEK293F-T1
現(xiàn)階段最常用的三大基因編輯技術(shù)主要有鋅指核酸酶技術(shù)(Zinc finger nucleases,ZFNs)、轉(zhuǎn)錄激活樣效應(yīng)子核酸酶技術(shù)(Transcription activatorlike effector nucleases,TALENs)、CRISPR/Cas9技術(shù)(Clustered regularly interspaced short palindromic Repeats/Cas9 nuclease)[1]。其中TALENs技術(shù)具有無上下文效應(yīng)[2]、脫靶效率低于ZFNs技術(shù)和CRISPR/Cas9技術(shù)[3]、特異性高等優(yōu)點(diǎn),現(xiàn)已成為一種廣泛應(yīng)用的基因定點(diǎn)修飾技術(shù)。
TALENs由轉(zhuǎn)錄激活樣效應(yīng)子蛋白(Transcription activator-like effector,TALE)、識(shí)別域和FokIⅡ核酸酶及切割域組成[4]。其中,TALE蛋白源自黃單胞桿菌,一般由N端、C端和中央串聯(lián)重復(fù)模塊組成,N端包含Ⅲ型轉(zhuǎn)運(yùn)信號(hào)(Translocation signal);C端包含核定位信號(hào)(Nuclear localization signal,NLS)和酸性轉(zhuǎn)錄激活域(Activation domain,AD);中央重復(fù)模塊具有“一模塊識(shí)別一堿基”的特點(diǎn),且沒有上下文效應(yīng),其結(jié)構(gòu)高度保守,一般由34個(gè)氨基酸組成,僅在12和13位高度可變,又稱這兩位的氨基酸為RVD(Repeat-variable diresidue)[2,5,6]。本研究采用并改造TALENs的精簡處理的骨架,即N端包含136個(gè)氨基酸,C端包含63個(gè)氨基酸,且C端融合有FokIⅡ型核酸內(nèi)切酶[7-10]。利用TALENs這些結(jié)構(gòu)特點(diǎn)可以構(gòu)建識(shí)別任何DNA序列的TALENs,包括核基因組和線粒體基因組[11,12]。
在TALENs應(yīng)用中,TALENs的中央重復(fù)模塊的組裝是限速步驟,目前為止,最常用的TALENs構(gòu)建方法大體可分為Golden Gate法[13-16]、FLASH(Fast Ligation-based automatable solid-phase highthroughput)法[17,18]、模塊組裝法(unit assembly,UA)法[7]及其他方法,如LIC法(Ligationindependent cloning)[19]、ICA法(Iterative capped assembly)[20]、idTALE法[21]、REAL法(Restriction enzyme and ligation)[22]等。其中Golden Gate法、FLASH法最為省時(shí),然而這兩種方法存在的酶切連接效率問題或因PCR技術(shù)的限制性而出現(xiàn)非特異的問題難以避免,相比之下,UA法雖構(gòu)建時(shí)間較長,但TALENs編碼序列可控且沒有上述問題,同時(shí)可以根據(jù)密碼子簡并原則改變TALEs編碼序列,降低TALENs重復(fù)模塊編碼序列的重復(fù)性,降低mRNA的重復(fù)性,有助于提高TALENs在真核細(xì)胞內(nèi)表達(dá)效率。本研究提供了TALENs的一單元模塊編碼序列庫,并改造表達(dá)載體pCS2-peas/perr,利用UA法定向組裝TALENs。鑒于目前沒有報(bào)告清晰的介紹線粒體DNA(Mitochondrial DNA,mtDNA)雙鏈斷裂(Double-strand breaks,DSB)的修復(fù)機(jī)制,本研究將mtDNA靶序列隨機(jī)整合于核染色體中,利用核基因組的同源重組(Homologous recombination,HR)和非同源末端連接(Non-homologous DNA end joining,NHEJ)修復(fù)方式[23],以檢測新型模塊組裝法構(gòu)建的TALENs活性,旨在為編輯mtDNA提供新方法。
1 材料與方法
1.1 材料
1.1.1 細(xì)胞 HEK293F細(xì)胞為本實(shí)驗(yàn)室保存。
1.1.2 試劑和引物 Xba I、Nhe I、AsiS I、Sac I、BamH I等限制性核酸內(nèi)切酶購自NEB公司;質(zhì)粒小提試劑盒購自北京莊盟國際生物公司;高純度小提中量試劑盒購自天根生物科技公司;HA、Flag、β-actin抗體購自Sigma公司;Anti-Rabbit、Antimouse抗體購自CST公司;轉(zhuǎn)染試劑X-tremeGENE HP DNA Transfection Reagent購自Roche公司;DMEM高糖培養(yǎng)基購自Invitrogen公司;胎牛血清購自Biowest公司;pUC19、pEGFP-N1質(zhì)粒由本實(shí)驗(yàn)室保存;pCS2-peas/perr質(zhì)粒購自康為世紀(jì)公司;所有引物序列(表1)由生工生物工程(上海)股份有限公司合成;測序由華大基因公司完成。
1.2 方法
1.2.1 細(xì)胞的培養(yǎng) HEK293F細(xì)胞以及HEK293FT1細(xì)胞用含10%胎牛血清的DMEM培養(yǎng)基培養(yǎng)于37℃、5%CO2溫箱中。
1.2.2 pCS2-peas/perr載體骨架的改造 本研究于載體0.5單元的RVD編碼序列兩側(cè)分別構(gòu)建AsiS I和Xba I酶切位點(diǎn),如圖1所示。實(shí)驗(yàn)方法如下:以pCS2-peas/perr載體質(zhì)粒為模板,TALEN0.5_N-F/R為引物對(duì),擴(kuò)增得到全長質(zhì)粒對(duì),PCR產(chǎn)物經(jīng)DpnI酶切,轉(zhuǎn)化后挑單克隆菌落,pCS2-NF/CR引物對(duì)測序鑒定出陽性質(zhì)粒,最終得到載體質(zhì)粒pCS2-0.5U-peas/perr。
1.2.3 TALEs一單元模塊的構(gòu)建 基于TALEs自然單元模塊序列[6]和UA法的人工單元模塊序列[7],本研究設(shè)計(jì)的人工單元模塊序列命名為偏單元(圖2),起始于TALEs自然模塊的+19位氨基酸殘基Leu,終止于下一個(gè)自然模塊的+18位氨基酸殘基Ala。于偏單元序列的兩端分別構(gòu)建Xba I、Nhe I同尾酶識(shí)別位點(diǎn),于RVD區(qū)附近構(gòu)建AsiS I酶切位點(diǎn)。根據(jù)AsiS I酶切位點(diǎn)的有無分為活動(dòng)一單元模塊(含AsiS I酶切位點(diǎn))和固定一單元模塊(不含AsiS I酶切位點(diǎn))。活動(dòng)一單元模塊位于TALEs重復(fù)模塊的首位,并命名為0序列,依次標(biāo)記為A0、C0、G0、T0。固定一單元模塊位于第2位及后續(xù)位置。由于氨基酸密碼簡并性和第4位、32位氨基酸殘基的高度可變,TALEN模板序列數(shù)目模大,經(jīng)排列組合計(jì)算后結(jié)果如表2所示。選取了氨基酸序列一致的4種基本骨架a1、c1、g1、t1,堿基序列不同但差異率并非最大(折中差異性),N-LETVQRLLPVLCQ AHGLTPEQVVAIASNIGGKQA-C(灰色背景部分為RVD區(qū))。依次構(gòu)建含有4種RVD(NI、HD、NN、NG)的一單元模塊,依次標(biāo)記為A1、A2、A3、A4,C1、C2、C3、C4,G1、G2、G3、G4,T1、 T2、T3、T4,如表3所示。

表1 pCS2-0.5U-peas/perr構(gòu)建所需引物
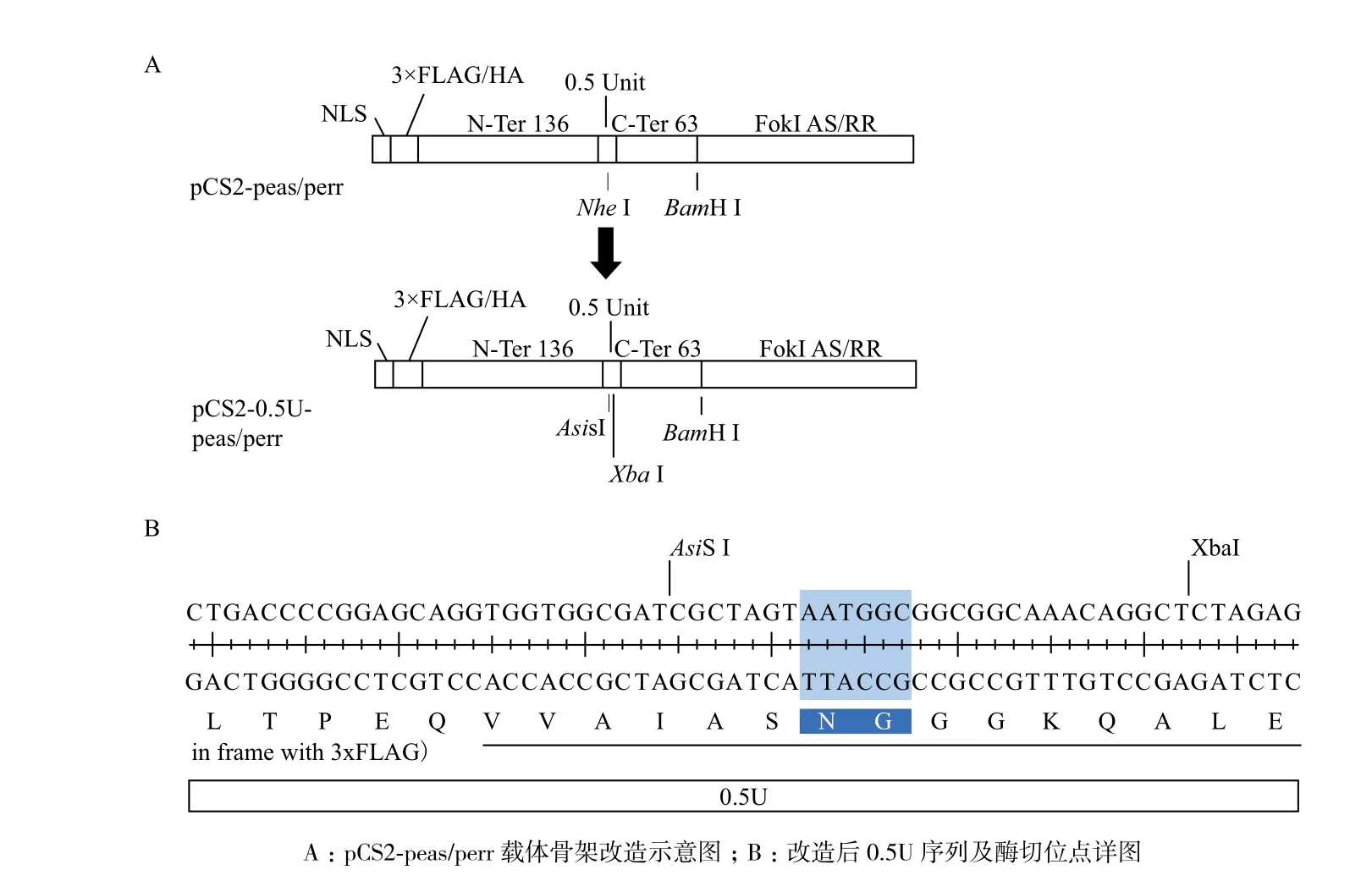
圖1 pCS2-0.5U-peas/perr構(gòu)建示意圖
活動(dòng)單元的構(gòu)建以C0序列為例,構(gòu)建方法如下:以C0的DNA編碼序列(表3)為模板分別合成Oligo片段C0-L1、C0-L2和C0-S1、C0-S2(表4),C0-L1/C0-L2、C0-S1/C0-S2分別退火,得到片段C0-L、C0-S,對(duì)pUC19質(zhì)粒進(jìn)行BamH I/Sac I雙酶切,將片段C0-S連入,再次進(jìn)行Xba I/AsiS I雙酶切,將片段C0-L連入,得到包含C0序列的pUC19-C0質(zhì)粒(圖3)。合成A0-S1/A0-S2、G0-S1/G0-S2(NN)、T0-S1/T0-S2(序列見表4),兩兩退火后,分別連入AsiS I/Nhe I雙酶切后的pUC19-C0偏單元載體,得到分別包含A0、G0和T0序列的質(zhì)粒pUC19-A0、pUC19-G0、pUC19-T0。從合成方法來看,A0、C0、T0和G0序列除了RVD區(qū)編碼序列不同外,其他位置編碼序列均相同。
固定單元的構(gòu)建以A1為例,構(gòu)建方法如下:根據(jù)A1的DNA編碼序列(表3),合成Oligo片段A1-L1、A1-L2和A1-S1、A1-S2(表5),分別退火,得到片段A1-L、A1-S。以A1-L1和A1-S2為引物,A1-L、A1-S為模板,擴(kuò)增得到A1。以A1為骨架,RVD區(qū)分別換為其他3種RVD(HD、NN和NG)的編碼序列,可得到C2、G2和T2。其他模塊構(gòu)建方法為,將A1-S2的RVD去分別改為其他3種RVD(HD、NN和NG)的編碼序列,命名為C2-ar、G2-ar和T2-ar(表5),分別和A1-L1組成引物對(duì),以A1為模板,擴(kuò)增得到C2、G2和T2。最終得到A1、C1、G1、T1,A2、C2、G2、T2,A3、C3、G3、T3,A4、C4、G4、T4。
設(shè)置射頻通道的本振頻率前,先設(shè)置多功能管腳為鎖相環(huán)鎖定檢測管腳,將鑒相器增益還原為默認(rèn)值;由于未使用到芯片內(nèi) IF鎖相環(huán),則關(guān)閉IF鎖相環(huán)電源;開啟RF鎖相環(huán)電源。

圖2 TALEs偏單元的氨基酸序列示意圖
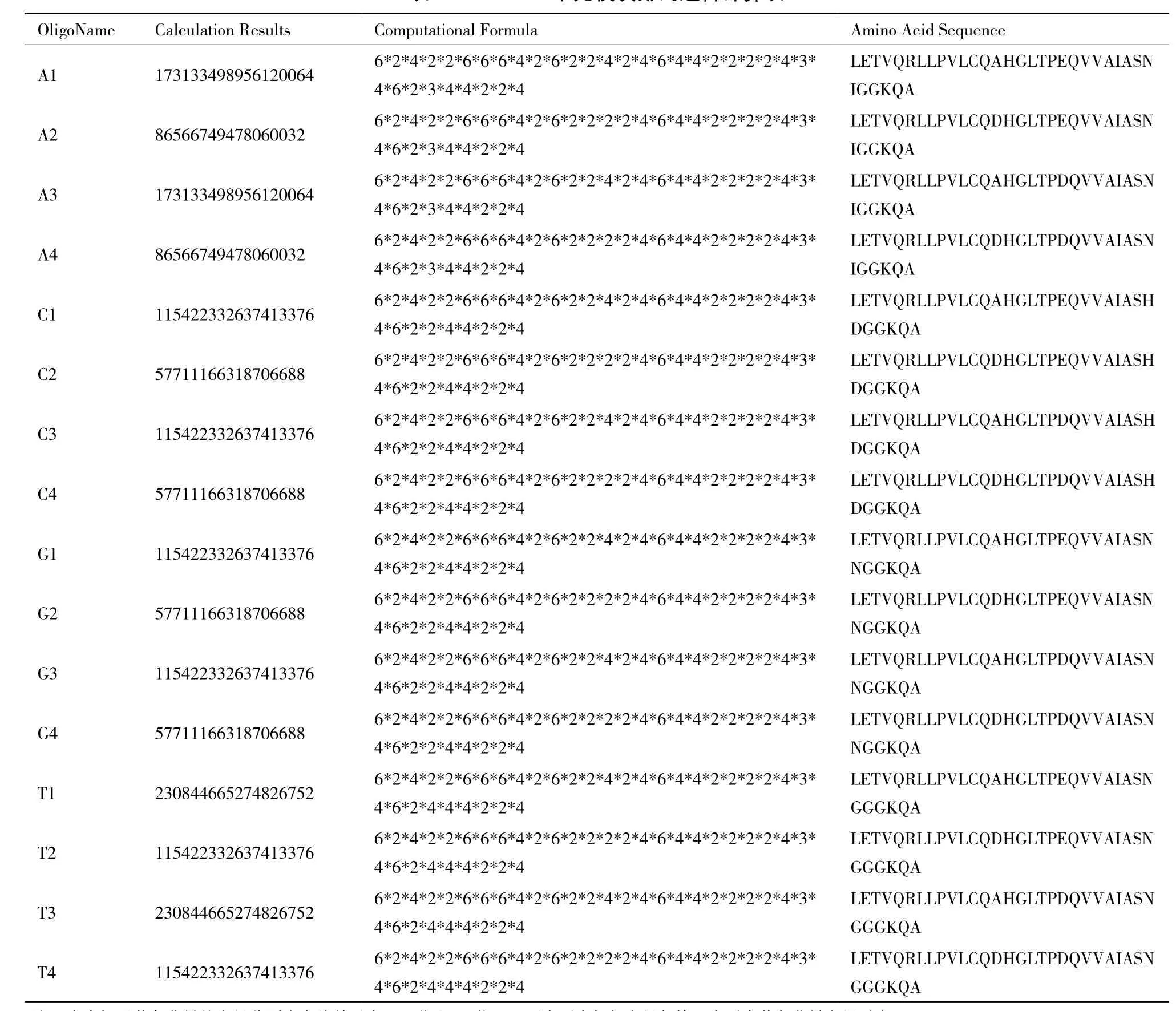
表2 TALE一單元模塊排列組合計(jì)算表

表3 0序列與4×4一單元模塊序列表
1.2.4 靶序列的選擇與TALEs中央重復(fù)模塊的組裝 在NCBI查詢得到人線粒體DNA序列(NC_012920.1),利用TALE-NT 2.0(TAL Effector Nucleotide Targeter 2.0)設(shè)計(jì)一系列TALENs靶序列。經(jīng)過靶位點(diǎn)比對(duì)和篩選,本研究選取選擇線粒體DNA(NC_012920.1)的16347-16387位為靶序列,5'-TCAAATCCCTTCTCGTCCCCATGGATGACCC CCCTCAGA TA-3',并依據(jù)該靶序列設(shè)計(jì)TALEs的串聯(lián)重復(fù)模塊。本研究設(shè)計(jì)選取12.5個(gè)TALEs模塊,Left-arm:5' -CAAATCCCTTCTC-3';Right-Arm:5'-ATCTGAGGGGGGT-3'。
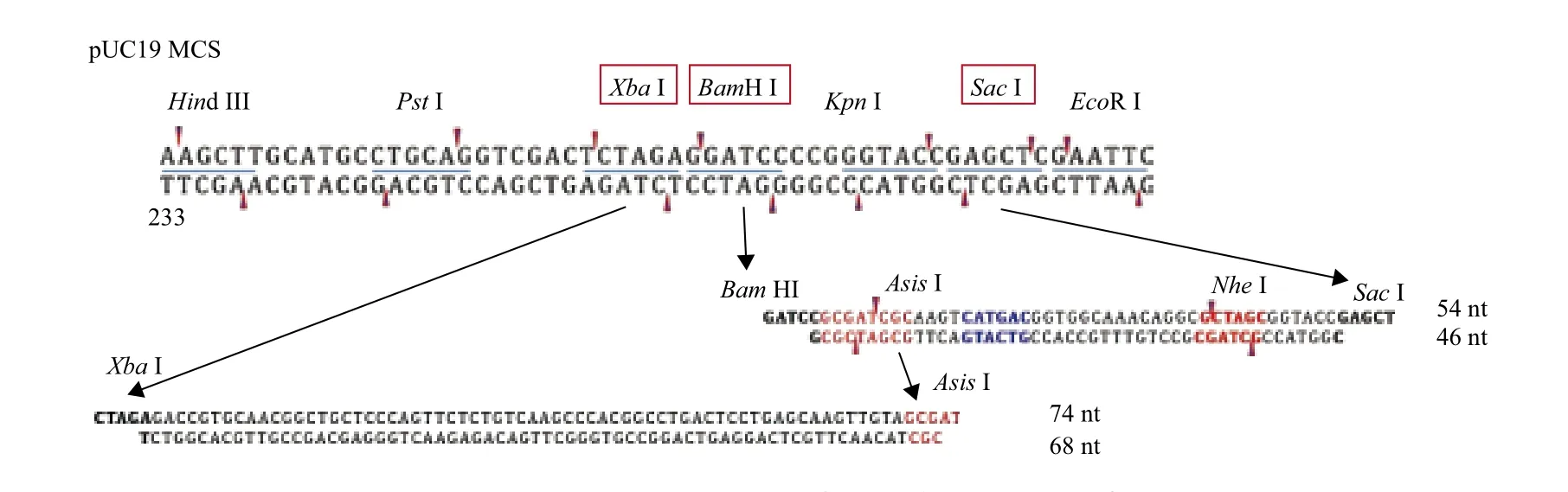
圖3 TALEs偏單元重復(fù)模塊的堿基序列設(shè)計(jì)示意圖

表5 4×4序列構(gòu)建中所需oligo序列表
用Nhe I/Sac I和Xba I/sac I分別對(duì)模塊載體和目的模塊序列進(jìn)行酶切、連接,依次由一單元得到二單元,得到四單元,再根據(jù)設(shè)計(jì)序列進(jìn)行靈活連接[7]。本研究組構(gòu)建的三單元模塊庫,分別由4個(gè)三單元模塊兩兩相連,得到2個(gè)六單元模塊,再次相連后與活動(dòng)一單元連接,最后得到13單元TALE-13U,Left-arm的構(gòu)建如圖4所示,然后將其進(jìn)行AsiS I和Nhe I(Xba I的同尾酶)雙酶切,連接入pCS2-0.5U-peas/perr載體,得到pCS2-12.5U-L/R質(zhì)粒對(duì)。
圖4 TALE-13U構(gòu)建示意圖
1.2.5 HEK293F-T1細(xì)胞系的構(gòu)建 合成5'端和3'端分別含有Nhe I、Age I酶切位點(diǎn)的靶序列片段,分別對(duì)靶序列片段和pEGFP-N1質(zhì)粒進(jìn)行Nhe I/Age I雙酶切后,將二者進(jìn)行連接、轉(zhuǎn)化,挑取單克隆菌落,經(jīng)PCR檢測,測序驗(yàn)證含有靶序列的目標(biāo)質(zhì)粒,轉(zhuǎn)染培養(yǎng)于35 mm dish的HEK293F細(xì)胞,24 h后消化轉(zhuǎn)移至10 cm dish培養(yǎng),同時(shí)加G418篩選,在推薦藥物濃度附近設(shè)置濃度梯度,經(jīng)摸索得到G418的最適濃度為1 mg/mL。挑取單克隆細(xì)胞系,繼續(xù)培養(yǎng),經(jīng)PCR擴(kuò)增并測序,得到整合有靶序列的目標(biāo)細(xì)胞系,標(biāo)記為HEK293F-T1。
1.2.6 TALENs在細(xì)胞內(nèi)表達(dá)活性的檢測 將HEK293F-T1細(xì)胞接種于兩個(gè)10 cm dish中,待兩盤細(xì)胞長至生長對(duì)數(shù)期且細(xì)胞密度一致(約24 h),選取其中一盤細(xì)胞瞬時(shí)轉(zhuǎn)染1.2.4中構(gòu)建的質(zhì)粒對(duì)pCS2-12.5U-L/R,另一盤作為control,再繼續(xù)培養(yǎng)48 h,收細(xì)胞,提取基因組,PCR擴(kuò)增后測序鑒定打靶效果。PCR反應(yīng)為活性檢測的關(guān)鍵步驟,反應(yīng)體系為:基因組(模板)200 ng,上/下游引物各0.5μL,LA Taq 0.5 μL,10×LA PCR BufferⅡ 2.5 μL,dNTP Mixture(各2.5 mmol/L)4 μL,ddH2O補(bǔ)足至25 μL。反應(yīng)程序:95℃ 2 min;95℃ 15 s、60℃ 30 s,72℃ 1 min(35個(gè)循環(huán));72℃ 5 min。
2 結(jié)果
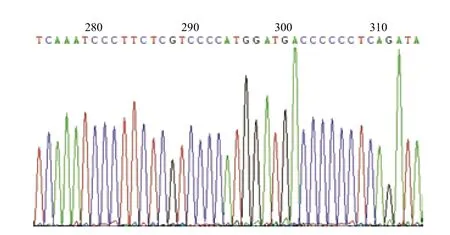
圖5 mtDNA中靶序列測序峰圖
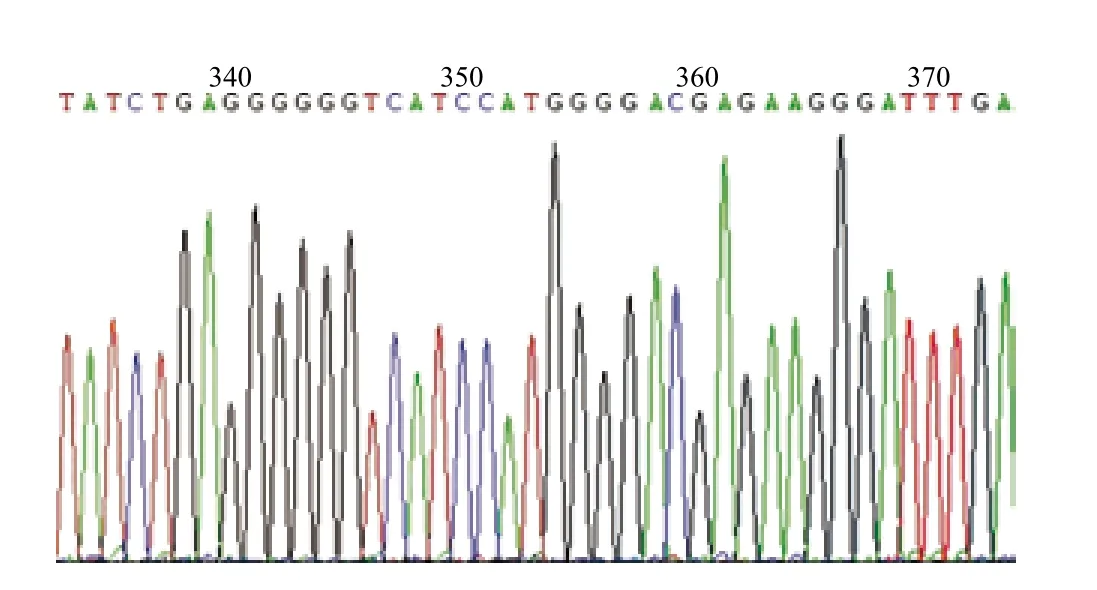
圖6 HEK293F-T1基因組中靶序列測序峰圖
2.1 靶序列的確定
2.1.1 HEK293F細(xì)胞線粒體中靶序列的確定 依據(jù)線粒體DNA(NC_012920.1),在靶序列附近設(shè)計(jì)引物,以提取的HEK293F基因組為模板,擴(kuò)增含有靶序列的片段并測序,測序結(jié)果峰圖如圖5所示。
2.1.2 細(xì)胞系HEK293F-T1中靶序列的確定 提取HEK293F-T1細(xì)胞基因組并作為模板,在pEGFPN1-T1質(zhì)粒靶序列兩側(cè)設(shè)計(jì)引物,擴(kuò)增靶序列,測序結(jié)果峰圖6如圖所示。
2.2 TALENs的構(gòu)建及酶切鑒定
模塊組裝法組裝TALENs,構(gòu)建得到pCS2-12.5U-L/R質(zhì)粒,經(jīng)AsiS I/BamH I酶切鑒定,片段為1 448 bp,大小如圖7-A所示,測序結(jié)果經(jīng)Blast比對(duì),左右臂的氨基酸比對(duì)結(jié)果如圖7-B所示。
2.3 TALENs活性檢測
通過提取瞬時(shí)轉(zhuǎn)染pCS2-12.5U-L/R質(zhì)粒的HEK293F-T1基因組,擴(kuò)增、測序分析后,觀察到靶序列強(qiáng)峰下出現(xiàn)套峰,結(jié)果(圖8)表明,TALENs成功的作用于HEK293F-T1基因組上的靶序列,造成雙鏈斷裂后,在細(xì)胞核內(nèi)進(jìn)行了修復(fù)。
3 討論
3.1 TALENs編碼序列重復(fù)性與活性
TALENs識(shí)別域是由n個(gè)串聯(lián)的高度重復(fù)的模塊組成,在細(xì)胞轉(zhuǎn)錄與翻譯過程中,能否對(duì)高度重復(fù)的序列十分準(zhǔn)確的轉(zhuǎn)錄翻譯成重復(fù)的蛋白模塊,至今還未有研究組論證,本研究組嘗試著盡量降低TALENs編碼序列的重復(fù)性,而模塊組裝法恰好能夠保證構(gòu)建的TALENs編碼序列的高保真性[24],使得TALENs編碼序列重復(fù)性可控,提高了TALENs的表達(dá)效率。
3.2 針對(duì)mtDNA的相關(guān)編輯研究
因mtDNA的堿基序列長度遠(yuǎn)遠(yuǎn)小于nDNA(Nuclear DNA),且線粒體中針對(duì)雙鏈斷裂的修復(fù)機(jī)制尚未準(zhǔn)確解析。本研究組中將mtDNA序列整合于核基因組中,利用核基因組中的HR和NHEJ機(jī)制對(duì)其進(jìn)行修復(fù),同時(shí)考慮到TALENs中央重復(fù)模塊的數(shù)目是否會(huì)影響打靶效率,本研究中設(shè)計(jì)的TALENs均選取12.5個(gè)重復(fù)模塊,對(duì)nDNA打靶效果比較理想,進(jìn)而為mtDNA的編輯研究奠定了基礎(chǔ)。Bacman[12]實(shí)驗(yàn)組首次利用TALENs編輯mtDNA的相關(guān)研究,為本實(shí)驗(yàn)室提供了可參考的思路與方法,本實(shí)驗(yàn)組下一步將進(jìn)行mtTALENs(Mitochondriatargeted TALENs)的研究。

圖7 pCS2-12.5U-L/R質(zhì)粒酶切(A)及pCS2-12.5U-L質(zhì)粒測序比對(duì)(B)結(jié)果
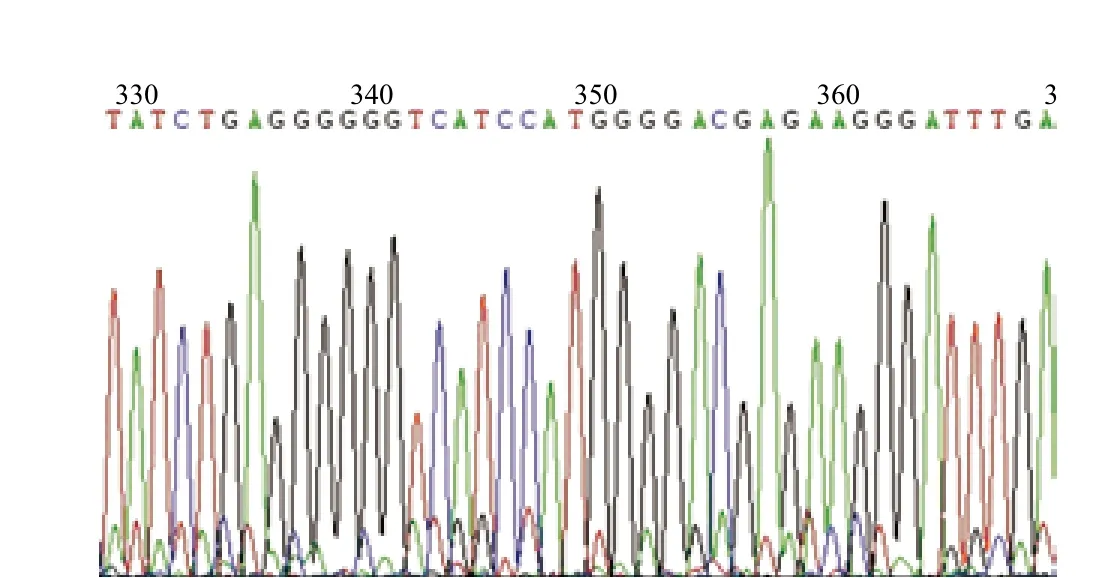
圖8 pCS2-12.5U-L/R打靶結(jié)果圖
4 結(jié)論
本研究中提供的新型模塊組裝法可定向的組裝TALENs,提高克隆效率,并增加了靶位點(diǎn)設(shè)計(jì)的靈活性,在表達(dá)強(qiáng)度和打靶效果上均可達(dá)到理想效果。
[1] Gaj T, Gersbach CA, Barbas CF. ZFN, TALEN, and CRISPR/ Cas-based methods for genome engineering[J]. Trends in Biotechnology, 2013, 31(7):397-405.
[2] Moscou MJ, Bogdanove AJ. A Simple Cipher Governs DNA Recognition by TAL Effectors[J]. Science, 2009, 326(5959):1501.
[3] Chen L, Tang L, Xiang H, et al. Advances in genome editing technology and its promising application in evolutionary and ecological studies[J]. GigaScience, 2014, 3:24.
[4] Christian M, Cermak T, Doyle EL, et al. Targeting DNA doublestrand breaks with TAL effector nucleases[J]. Genetics, 2010,186(2):757-761.
[5] Boch J, Scholze H, Schornack S, et al. Breaking the code of DNA binding specificity of TAL-type III effectors[J]. Science, 2009,326(5959):1509-1512.
[6] Boch J, Bonas U. Xanthomonas AvrBs3 family-type III effectors:discovery and function[J]. Annual Review of Phytopathology,2010, 48:419-436.
[7] Huang P, Xiao A, Zhou MG, et al. Heritable gene targeting in zebrafish using customized TALENs[J]. Nature Biotechnology,2011, 29(8):699-700.
[8] Miller JC, Tan S, Qiao G, et al. A TALE nuclease architecture forefficient genome editing[J]. Nature Biotechnology, 2011, 29(2):143-148.
[9] Szurek B, Rossier O, Hause G, et al. Type III-dependent translocation of the Xanthomonas AvrBs3 protein into the plant cell[J]. Mol Microbiol, 2002, 46(1):13-23.
[10] Mussolino C, Morbitzer R, Lutge F, et al. A novel TALE nuclease scaffold enables high genome editing activity in combination with low toxicity[J]. Nucleic Acids Research, 2011, 39(21):9283-9293.
[11] Li T, Liu B, Spalding MH, et al. High-efficiency TALEN-based gene editing produces disease-resistant rice[J]. Nature Biotechnology, 2012, 30(5):390-392.
[12] Bacman SR, Williams SL, Pinto M, et al. Specific elimination of mutant mitochondrial genomes in patient-derived cells by mitoTALENs[J]. Nature Medicine, 2013, 19(9):1111-1113.
[13] Cermak T, Doyle EL, Christian M, et al. Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting[J]. Nucleic Acids Research, 2011,39(12):e82.
[14] Weber E, Gruetzner R, Werner S, et al. Assembly of designer TAL effectors by Golden Gate cloning[J]. PloS One, 2011, 6(5):e19722.
[15] Zhang F, Cong L, Lodato S, et al. Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription[J]. Nature Biotechnology, 2011, 29(2):149-153.
[16] Sanjana NE, Cong L, Zhou Y, et al. A transcription activator-like effector toolbox for genome engineering[J]. Nature Protocols,2012, 7(1):171-192.
[17] Reyon D, Tsai SQ, Khayter C, et al. FLASH assembly of TALENs for high-throughput genome editing[J]. Nature Biotechnology,2012, 30(5):460-465.
[18] Reyon D, Maeder ML, Khayter C, et al. Engineering customized TALE nucleases (TALENs) and TALE transcription factors by fast ligation-based automatable solid-phase high-throughput (FLASH)assembly[J]. Current Protocols in Molecular Biology, 2013,Chapter 12:Unit 12.6.
[19] Schmid-Burgk JL, Schmidt T, Kaiser V, et al. A ligation-independent cloning technique for high-throughput assembly of transcription activator-like effector genes[J]. Nature Biotechnology, 2013, 31(1):76-81.
[20] Briggs AW, Rios X, Chari R, et al. Iterative capped assembly:rapid and scalable synthesis of repeat-module DNA such as TAL effectors from individual monomers[J]. Nucleic Acids Research,2012, 40(15):e117.
[21] Li L, Piatek MJ, Atef A, et al. Rapid and highly efficient construction of TALE-based transcriptional regulators and nucleases for genome modification[J]. Plant Mol Biol, 2012, 78(4-5):407-416.
[22] Sander JD, Cade L, Khayter C, et al. Targeted gene disruption in somatic zebrafish cells using engineered TALENs[J]. Nature Biotechnology, 2011, 29(8):697-698.
[23] Larsen NB, Rasmussen M, Rasmussen L J. Nuclear and mitochondrial DNA repair:similar pathways?[J]. Mitochondrion, 2005, 5(2):89-108.
[24] 沈延,肖安,黃鵬,等. 類轉(zhuǎn)錄激活因子效應(yīng)物核酸酶(TALEN)介導(dǎo)的基因組定點(diǎn)修飾技術(shù)[J]. 遺傳, 2013, 35(4):395-409.
(責(zé)任編輯李楠)
A Novel Method of Unit Assembly and Activity Assay for Transcription Activator-like Effector Nucleases
GAO Jing1,2WEI Di1,2CHI Zhen-fen3ZHANG Gui-rong1,2ZHANG Wan-ming1NIE Ling-yun1,2
(1. Hebei North University,Zhangjiakou 075000;2. Institute for Drug and Instrument Control,Health Department,the General Logistics Department of People’s Liberation Army,Beijing 100166;3. Beijing Institute of Genomics,Chinese Academy of Sciences,Beijing 100101)
A novel method of unit assembly(UA)and activity assay based on transcription activator-like effector nucleases(TALENs)are described. An online tool TALE-NT 2.0 was used to design recognition and splice sites on mitochondrial DNA(mtDNA)of TALENs. By means of pEGFP-N1 plasmid, the segments of target sequence were randomly integrated in the nuclear genome of HEK293F. The constructed cell line HEK293F-T1 was for activity assay of TALENs. Based on transcription activator-like effector(TALE)natural repeats, a new type of artificial TALEs single-unit repeats were designed. According to the recognition sites of TALENs, appropriate single-unit repeats were selected and assembled by UA. TALEs series unit sequence containing the corresponding double enzyme cutting sites and TALENs vector sequence were designed, which was directionally ligated and transiently transfected into HEK293F-T1 cell line. The results showed that the TALES units were directionally assembled by new method, and it had a high cloning efficiency;moreover, the nested peaks clearly appeared after transient transfection. In conclusion, the novel method of unit assembly improves cloning efficiency of constructing TALLENs, even is not limited by repeat-variable residue(RVD)in the last 0.5 unit, and increases the flexibility of designing the target sequences.
transcription activator-like effector nucleases;unit assembly;mitochondrial DNA;HEK293F-T1
10.13560/j.cnki.biotech.bull.1985.2016.01.035
2015-03-04
國家自然科學(xué)基金面上項(xiàng)目(31371346)
高敬,女,碩士研究生,研究方向:遺傳學(xué)研究新技術(shù)與新方法;E-mail:gaojing201201@126.com
聶凌云,女,博士,研究方向:遺傳學(xué)研究新技術(shù)與新方法;E-mail:nielingyun@126.com
