基于高通量測序的微生物強化污泥減量工藝中微生物群落解析
2016-10-13 21:37:26宋云龍張金松邵明非鄧仁健
中國環境科學 2016年7期
宋云龍,張金松*,朱 佳,邵明非,鄧仁健
?
基于高通量測序的微生物強化污泥減量工藝中微生物群落解析
宋云龍1,張金松1*,朱 佳2,邵明非1,鄧仁健1
(1.哈爾濱工業大學深圳研究生院,土木與環境工程學院,廣東 深圳 518055;2.深圳職業技術學院建筑與環境工程學院,廣東 深圳 518055)
利用高通量測序技術對微生物強化原位污泥減量工藝中微生物群落進行解析.結果表明,外源菌劑的投加改變了活性污泥中微生物群落結構,菌劑的有效成分乳桿菌屬和醋酸桿屬在強化組中含量顯著增加.科水平上的群落多樣性分析顯示:強化組厭氧區Siompson指數、Shannon指數和Pielou指數均不同程度升高,群落多樣性增加.聚類分析顯示,微生物群落按時間順序明顯聚為3簇,強化組厭氧區微生物群落隨時間發生較大演變.主坐標分析顯示,強化組和對照組微生物群落明顯聚為2類,其厭氧區群落分別按照不同的方向演變,微生物強化和DO降低的協同作用是強化組群落演變驅動力,DO降低是對照組群落演變驅動力.典范對應分析顯示顯著影響微生物群落結構的環境因子依次為pH值、水溫和DO.
高通量測序;微生物強化;水解酸化;群落解析
微生物強化技術在污水處理系統中主要用于去除難降解有機物[1-4],近年來由于我國污泥問題日益嚴峻[5-7],研究者嘗試將微生物強化技術用于污泥的處理處置[8-9].微生物強化原位污泥減量作為一種新興、高效的污泥減量技術受到廣泛關注,并取得了顯著的污泥減量效果[10-12],但是由于其減量機理尚不明確,因而尚未得到大范圍內推廣應用.現有的微生物強化污泥減量機理分析多停留在理化性質層面,而微生物強化對微生物群落結構的影響方面的研究較少.現有微生物群落分析多是依靠PCR-DGGE技術,僅通過電泳圖譜中的幾十個條帶信息對微生物群落多樣性進行定性或半定量的分析, 而DGGE技術對于群落中相對豐度低于1%的非優勢物種檢測效果甚微[13],難以從多個分類層對物種豐度和群落特征進行更全面深入的信息挖掘[14-16].以Illumina測序為代表的第2代高通量測序具有測序成本低、信息采集量大、適用性廣等優點,成為近年來應用最廣泛的測序技術[17-20],為深度解析活性污泥微生物群落提供了有力的支持.
課題組前期通過投加菌劑的方式對A/O工藝活性污泥進行微生物強化作用,取得了30%左右的污泥減量效果[21].在此基礎上,進一步依托高通量測序平臺Illumina-HiSeq2000,研究了低污泥濃度工況中外源菌劑的投加對活性污泥微生物群落結構的影響,從群落演變的層面探討了A/O工藝中微生物強化原位污泥減量的機理,為微生物強化原位污泥減量技術的規模化推廣應用提供理論依據.
1 材料與方法
1.1 A/O中試試驗
1.1.1 中試裝置 裝置位于深圳市羅芳污水處理廠,整個系統為兩套完全平行且獨立運行的A/O工藝中試裝置,模擬羅芳污水處理廠一期處理工藝,單臺設備處理規模240m3/d,構造如圖1所示,其厭氧1區、厭氧2區、好氧1區、好氧2區和沉淀池有效容積分別為12.5,25,22,22,30m3.
1.1.2 試驗污水污泥 中試進水取自深圳羅芳污水處理廠一期工程的中間沉淀區,具體水質如下:pH值為6.25~7.68,均值7.03;COD為102~ 334mg/L,均值155mg/L;BOD5為47~135mg/L,均值67mg/L;總氮為21~52mg/L,均值32mg/L;氨氮為13~42mg/L,均值21mg/L;總磷為1.59~ 6.97mg/L,均值3.25mg/L.試驗污泥取自深圳羅芳廠一期AA/O工藝好氧區.
1.1.3 菌劑投加及運行方式 菌劑投加量為日處理水量(240m3/d)的萬分之四(96kg),投加點為厭氧2區,一次性投加.投加微生物菌劑的稱強化組,未投加微生物菌劑稱對照組.中試運行條件模擬羅芳污水處理廠的生產工況:反應體系MLSS維持在3500~4000mg/L,好氧區DO維持在3~5mg/L,厭氧區DO維持在0.05~0.15mg/L,污泥回流比為1~1.2,強化組和對照組污泥齡分別為28.04和17.99d,兩組HRT均為8.15h,在保證出水達到GB18918-2002[22]規定的一級A標準的前提下盡量減少排泥.
1.2 試驗方法
1.2.1 取樣 取強化組和對照組厭氧2區的第3,10,21d的活性污泥,相應標記為d3-Y1、d3-Y2、d10-Y1、d10-Y2、d21-Y1、d21-Y2,取強化組和對照組好氧2區的第5、21d的活性污泥,相應標記為d5-H1、d5-H2、d21-H1、d21-H2.活性污泥樣品經0.2mm玻璃纖維濾膜過濾后-80℃保存.
1.2.2 檢測方法 用FastDNA?土壤試劑盒(MP Biomedicals,Illkirch,France)進行細菌組DNA提取.16S rRNA基因V6區序列采用引物對967F (CAACGCGAAGAACCTTACC)與1046R (CGACAGCCATGCANCACCT)進行擴增[23]. PCR條件:98℃變性2min,然后28個循環(98℃20s,55℃20s, 68℃1min),最后在68℃延伸5min.為減少PCR擴增前期的潛在錯誤,每DNA樣品同步進行2個平行擴增,最后再進行混合與后續分析.擴增后采用2.0%的瓊脂糖凝膠電泳對擴增產物進行驗證.最后,PCR擴增產物采用純化試劑盒(天根,北京)進行純化,并采用光度法(NanoDrop-1000)測定產物濃度后混合樣品,保證各樣品核酸濃度基本相同.最后采用Illumina- HiSeq2000(深圳華大基因)高通量測序平臺測序后進行群落解析.
1.2.3 多樣性指數的計算 辛普森多樣性指數(Simpson's diversity index,)[24]:
式中:p表示種的個體在群落中的比例;p2表示隨機取2個個體為同種的概率.
香濃多樣性指數(Shannon-Wiener,¢)[25]:
式中:表示總的物種數;p表示第個種占總數的比例.
Pielou均勻度指數(Pielou's evenness index,)[26]:
式中:¢為香農指數;是¢的最大值.
2 結果與討論
2.1 微生物強化污泥減量效果及污水處理效能
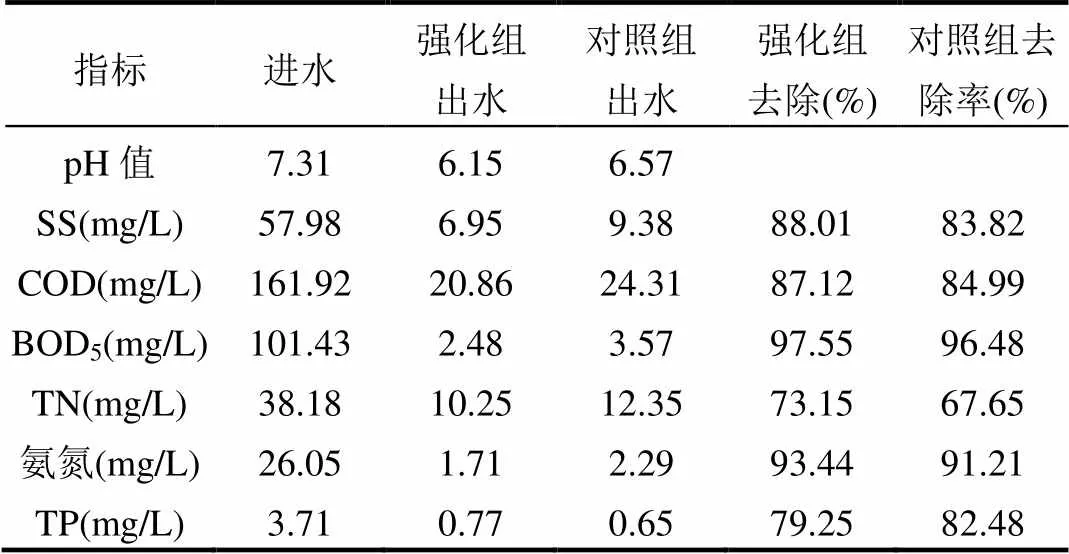
表1 微生物強化污水處理效果 Table 1 Pollutants removal efficiency by bioaugmentation
中試裝置有效運行了30d,取得了良好的污泥減量效果.強化組和對照組的MLSS產率分別為0.36,0.49kgMLSS/kgCOD,MLVSS產率分別為0.19,0.26kgMLSS/kgCOD,MLSS和MLVSS減量率分別為26.24%和28.09%.微生物強化污水處理效果見表1,可以看出,投加菌劑后,強化組SS、COD、BOD5、TN、氨氮的去除效果得到不同程度的加強,但TP的去除能力略微下降.強化組和對照組均出水達到GB18918-2002一級A標準.
2.2 高通量測序對微生物群落的鑒定效率
本次測序各樣品原始序列數分布比較均勻,除D21-H2較高之外,基本在10000條左右(表2).對原始DNA序列去除了古菌序列,并刪除了錯誤序列,最終將各樣品的DNA序列數統一為7341條.之后采用Uclust方法以及0.97的相似度對序列進行聚類,針對聚類獲得的OTU采用First方法抽取代表性序列,將代表序列與Greengene數據庫進行比對,完成對各OTU的注釋.從而將各樣本中每條序列鑒定到門、綱、目、科、屬、種等不同的分類水平.
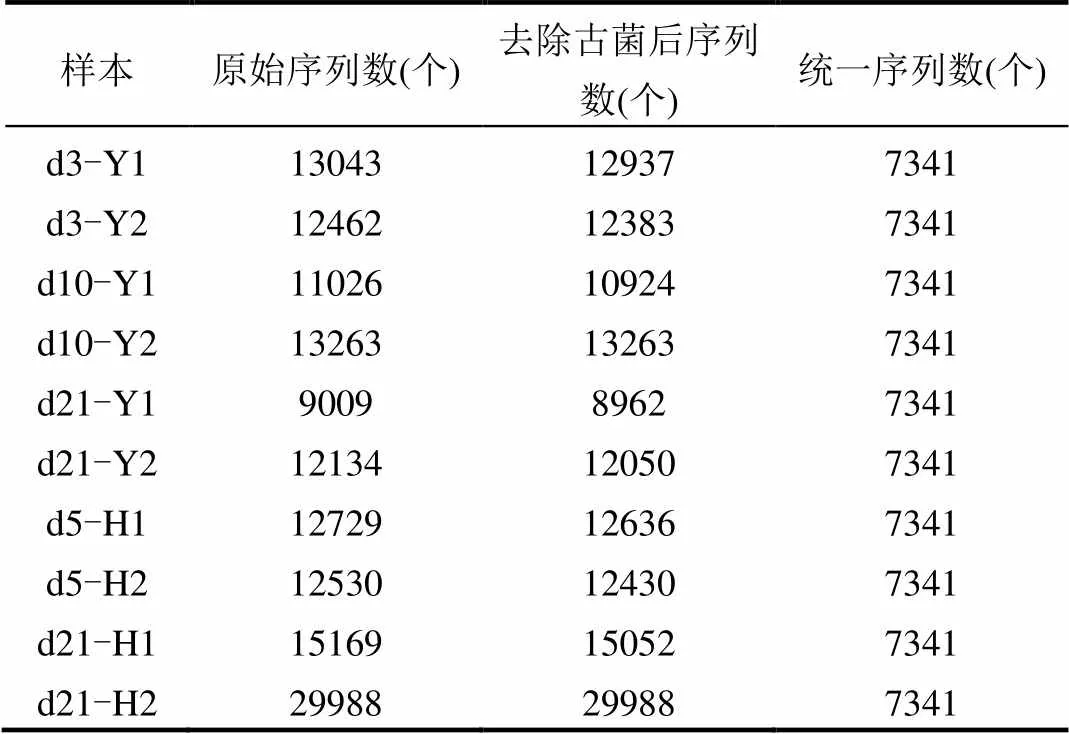
表2 各樣品測序深度 Table 2 Sequence depth of each sample
注:可鑒定序列是指本次測序測得序列中與Greengene數據庫已知物種序列相似度不低于97%的序列,低含量物種是指比例低于1%的可鑒定序列所對應的物種.
本次測序在門、綱、目、科、屬、種水平上的鑒定率分別是98.76%、94.58%、87.56%、82.65%、51.14%和0.69%,在門、綱、目、科水平鑒定結果良好,對屬水平也有一定鑒定效率,對種水平鑒定能力較差(表3).
2.3 微生物強化對群落組成的影響
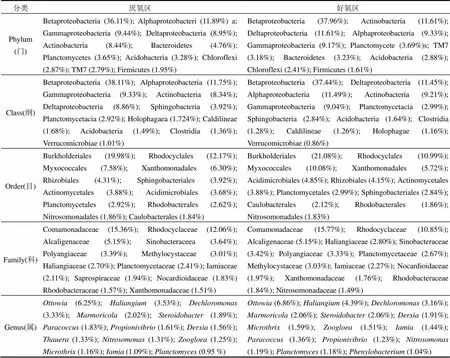
表4列舉了A/O中試裝置活性污泥中的優勢微生物,按照相對豐度降序排列.厭氧區和好氧區在門水平上的優勢微生物組成相種類相同,主要有變形菌門(Betaproteobacteria, Alphaproteobacteria, Gammaproteobacteria, Deltaproteobacteria)、放線菌門(Actinobacteria)、擬桿菌門(Bacteroidetes)、浮霉菌門(Planctomycetes)、酸桿菌門(Acidobacteria)和綠彎菌門(Chloroflexi),且按照豐度排序基本一致;在綱、目水平上,厭氧區和好氧區優勢微生物組成相同,但按照豐度排序差異較大;在科、屬水平上厭氧區與好氧區微生物組成種類基本相似,腐螺旋菌科(Saprospiraceae)和亞硝化單胞菌科(Nitrosomonadaceae)分別是厭氧區和好氧區的優勢科,索氏均屬(Thauera)和苯基桿菌屬(Phenylobacterium)分別是厭氧區和好氧區的優勢屬;厭氧區和好氧區的各科屬微生物豐度差異較大.
注:( )內為相對豐度.
微生物菌劑中最多的是厚壁門(Firmicutes),占細菌總數的53.48%,其次是變形門(Proteobacteria),占細菌總數的46.38%,其中對污泥水解酸化具有強化作用的主要是乳桿菌屬()和醋酸桿菌屬().乳桿菌屬屬于厚壁門(Firmicutes)、桿菌綱(Bacilli)、乳桿菌目(Lactobacillales)、乳桿菌科(Lactobacillaceae),醋酸桿菌屬屬于厚壁門(Firmicutes)、梭菌綱(Clostridia)、梭菌目(Clostridiales)、真桿菌科(Eubacteriaceae).乳桿菌屬是異養菌,耐酸性,能發酵分解糖代謝,終產物中50%以上是乳酸[27],在污水污泥處理中有廣泛的應用.Vellingiri等[28]利用馴化培養的嗜酸乳桿菌降解生活污水中總懸浮固體,同時對COD和BOD5都有很好的降解效果.Oh等[29]通過向厭氧發酵罐接種乳酸桿屬強化初沉污泥水解酸化,SCOD增加59%以上,同時產生了醋酸、乳酸、丙酸、丁酸和戊酸等多種有機酸.醋酸桿菌屬也是一類化能異養菌,耐酸性,能將乙醇氧化成醋酸,并可將醋酸和乳酸氧化成CO2、水和H2等[30].兩種菌屬在污水處理過程中的有機物水解酸化過程中都扮演著重要角色,常用作復合微生物菌劑的主要成分[31-32].對中試裝置活性污泥高通量分析結果顯示,第3,10,21d的強化組(對照組)厭氧區變形門分別為67.95%(64.71%)、66.96%(77.44%)、64.56%(74.66%),厚壁門分別為0.91%(1.37%)、2.39%(1.42%)、3.04%(2.01%).微生物強化使強化組的厚壁門含量增加了2倍多,變形門的增長受到抑制.屬水平上第3,10,21d的強化組(對照組)厭氧區醋酸桿屬相對豐度分別為3%(2%)、7%(2%)、8%(3%),乳桿菌屬相對豐度分別為5%(3%)、10%(4%)、9%(3%).可見菌劑中的功能組分在強化組中都顯著增長,并對活性污泥水解酸化起到了強化作用.
群落解析結果越接近真實,綜合考慮分類層次和鑒定率,對科水平的數據進行深入解析.圖2反映了A/O中試裝置厭氧區和好氧區不同運行階段的科水平的微生物群落組成,活性污泥中科水平微生物群落結構變化明顯.強化組和對照組的厭氧區中高含量物種的總比例變化差明顯.所占比例分別是71.80%、72.10%和69.99%,呈現出穩中微降的變化趨勢;對照組高含量物種第3,10,21d所占比例分別是73.75%、75.03%和75.30%,呈現逐步升高的變化趨勢.這說明外源菌劑的投加改變了強化組厭氧區微生物組成和分布.具體的,Comamonadaceae是A/O裝置活性污泥中含量最多的微生物,強化組中的Comamonadaceae由第3d的15.49%降到第21d的12.23%,而對照組中Comamonadaceae基本不變;強化組中Sinobacteraceae由5.03%降低至3.01%,對照組Sinobacteraceae基本恒定;同樣還有Alcaligenaceae、Haliangiaceae、Polyangiaceae和Saprospiraceae等在強化組中都明顯減少,在對照組中基本不變.其中Alcaligenaceae是一類產堿的好氧菌,它在強化組中含量的減少,是由于外加菌劑中的乳酸桿菌和醋酸桿菌等酸性菌在強化污泥水解酸化時降低了體系的pH值,通過種間競爭的方式抑制了Alcaligenaceae的增殖.Nitrosomonadaceae是亞硝化單胞菌科,在有機氮轉變成硝酸鹽的過程中起著重要作用.Nitrosomonadaceae變化規律說明強化組對蛋白質等有機氮分解能力明顯要強于對照組,這也是強化組的脫氮效果由于對照組的原因.
綜上所述,菌劑中的微生物投加到活性污泥后通過兩方面作用強化污泥減量,一是菌劑中乳桿菌屬和醋酸菌屬等功能微生物自身直接與污泥發生作用,強化了污泥水解酸化,減少污泥產量,二是通過分泌胞外物質(如抗生素)直接作用或通過改變生存環境(如pH值)等方式間接作用,促進或抑制反應體系中其它微生物的生長,改變原活性污泥中的微生物群落結構,影響了污泥產量.
2.4 微生物強化對群落多樣性的影響
多樣性指數和均勻度指數是反映微生物群落變化的綜合量度,Siompson指數反映了群落內微生物多樣性的概率,Shannon指數反映了群落多樣性的高低,Pielou指數反映了群落的均勻程度,三者均是指數越大,微生物分布越均勻,多樣性越高[24-26].圖3反映了科水平上的微生物群落多樣性變化,強化組厭氧區的Siompson指數隨著運行天數增加而升高,由第3d的13.07升高到第21d的14.42,增加了10.33%,而對照組厭氧區的Siompson指數則是先降低后回升,到第21d變為11.91,各個時期Siompson指數均是強化組高于對照組;好氧區Siompson指數則是強化組降低,對照組升高,強化組Siompson指數均低于對照組.強化組厭氧區Siompson指數升高是由于外加菌劑中多種外源微生物的加入增加了其微生物的種類,并改變了各微生物相對含量;而對照組Siompson指數降低則是因為體系中活性污泥取自羅芳污水廠好氧池,進入中試裝置厭氧區后由于pH值、DO和營養物質等生活條件的差異導致微生物群落發生小幅度調整,多樣性降低.好氧區Siompson指數變化規律相反,這是因為菌劑中的功能微生物多是厭氧或兼性厭氧微生物,在好氧區難以大量增殖,對其多樣性影響甚微. Shannon指數變化規律與Siompson指數相似.厭氧區的Pielou指數變化規律與Siompson指數相似,而兩組好氧區Pielou指數波動較小,無明顯差異.本試驗微生物菌劑主要投加到厭氧區,其有效微生物經過適當時間的增殖并與土著微生物充分作用,抑制了部分土著微生物的增殖,使得強化組厭氧區多樣性升高,由此改變微生物群落結構,增加了水解酸化菌的含量,并強化反應器中活性污泥的水解酸化,從而達到污泥減量的目的.
2.5 微生物強化對群落演變的影響
通過聚類分析研究了厭氧區和好氧區不同時期的微生物群落相似性(圖4),發現活性污泥微生物群落在時間尺度上發生明顯的演變,第3d和第5d的樣品聚為一類,第10d和第21d的樣品聚為一類,特別的,第 21d強化組厭氧區獨成一類.空間尺度上強化組和對照組對比來看,其厭氧區變化差異明顯:第3d的強化組和對照組距離最近,相似性最高;對照組第10d和第21d位于中間一支,與對照組第3d相比形成一定分化,d3-Y2與d21-Y2的差異為0.035;強化組厭氧區第3,10, 21d分別位于3個不同支,距離最遠,差異最大, d3-Y1與d21-Y1的差異為0.13;第21d時強化組和對照組群落差異達到0.09.這說明相對于對照組厭氧區,強化組厭氧區落隨時間的推移發生了更為明顯的演變,而外源微生物強化正是這種群落變化的驅動力.
對厭氧區和好氧區不同時期的微生物群落進行了主坐標分析,由圖5可知,強化組和對照組厭氧區的群落明顯分為2個區域,對照組群落沿著坐標1方向演變,而強化組群落是沿著坐標1和坐標2對角線方向演變,并且強化組的演變梯度是對照組的近2倍.強化組和對照組污泥和進水來源相同,中試運行工況如DO、pH值、MLSS等均保持一致,有無菌劑投加是二者唯一的差異,因此可知微生物強化是強化組微生物群落沿著不同于對照組方向進行演變的主要因素.對照組活性污泥由羅芳廠一期AA/O工藝好氧區引入到中試裝置后,該好氧污泥在對照組厭氧區由于DO等工藝參數的改變,其中微生物群落隨之發生演變,即DO的降低(由3~5mg/L降低為不足0.5mg/L)是微生物群落沿坐標1方向的演變驅動力.而微生物強化是微生物群落沿坐標2方向的演變驅動力,強化組微生物群落在2個驅動力綜合作用沿坐標1和坐標2對角線方向演變,并且箭頭更靠近坐標2,說明微生物強化的驅動作用更強.兩組好氧區的微生物群落在坐標1方向上演變較大,對照組在坐標2方向上基本無變化,強化組在坐標2方向上有小幅演變,這是因為微生物菌劑的有效組分是厭氧和兼性厭氧微生物,在好氧區無法大量繁殖,難以正常發揮作用.
選取水溫、pH值、DO和SCOD、TP、TN、氨氮、硝酸鹽氮8個指標,通過典范對應分析(Canonical Correspondence Analysis, CCA)研究科水平上微生物群落演變的關鍵影響因子.F1和F2的特征值分別為0.056和0.033,其解釋率分別為48.12%和28.43%,總解釋率為76.55%.pH值是微生物群落演變的最關鍵影響因子,其次是水溫和DO,而SCOD、TP、TN、氨氮、硝酸鹽氮的影響相對微弱.pH值的影響最大,說明外加菌劑的微生物強化通過影響強化組活性污泥pH值這一生存環境來改變期微生物群落演替方向,使得更多的水解酸化細菌在較低pH值環境中充分繁殖,強化了對活性污泥的水解酸化作用,達到污泥減量的目的.由于試驗是在12月末到1月期間進行,水溫波動幅度較大(8.9~17.9℃),對微生物群落演變產生較大影響.厭氧區DO為0.05~ 0.13mg/L,好氧區DO為3.09~3.75mg/L,DO的影響主要體現在厭氧區和好氧區微生物群落差異上.
3 結論
3.1 微生物強化使A/O中試裝置的MLSS和MLVSS分別減少了26.26%和28.08%,有效減少了剩余污泥產量.強化組的SS、COD、BOD5、TN和氨氮等污染物的去除率增加,但總磷的去除率略微降低,出水水質可穩定達到一級A標準.
3.2 高通量測序對活性污泥微生物群落在門、綱、目、科水平鑒定結果良好,屬水平上也有一定鑒定效率,種水平的鑒定效果較差.微生物菌劑主要成分是厚壁門的乳桿菌屬和醋酸桿菌屬,微生物強化使強化組活性污泥中厚壁門增加了2倍,乳桿菌屬和醋酸菌屬在活性污泥中屬于低含量菌屬,二者在對照組中含量基本不變,在強化組含量明顯上升.
3.3 微生物強化使強化組厭氧區Siompson指數、Shannon指數和Pielou指數均不同程度升高,群落多樣性增加.聚類分析顯示微生物群落按時間順序明顯聚為3簇,強化組厭氧區微生物群落隨時間發生較大演變.主坐標分析顯示強化組和對照組群落明顯聚為2類,其厭氧區群落分別按照不同的方向演變,微生物強化和DO降低的協同作用是其演變的推動力.典范對應分析顯示顯著影響微生物群落結構的環境因子依次為pH值、水溫和DO.
3.4 微生物強化使得活性污泥微生物群落結構發生明顯變化,菌劑通過直接與污泥作用強化污泥水解酸化,或通過改變生存環境(如pH值)等方式間接作用,促進或抑制反應體系中其它微生物的生長,改變原活性污泥中的微生物群落結構,減少污泥產量.
-chloroaniline slows bacterial species turnover rate in bioreactors [J]. Water Research, 2013,12,(47):7109-9119.
陳 雄,韋祎旸,李輝信,等.附著劍菌對三氯聯苯的吸附及降解機制研究 [J]. 中國環境科學, 2015,35(5):1423-1428.
Chang L K, Ibrahim D, Omar I C. A laboratory scale bioremediation of Tapis crude oil contaminated soil by bioaugmentation of Acinetobacter baumannii T30C [J]. African Journal Of Microbiology Research, 2011,9(5):2609-2615.
侯 穎,王 飛,董維亮,等. Rhodococcus sp. T3-1菌株降解乙草胺的特性 [J]. 中國環境科學, 2013,33(10):1785-1790.
Zhu L L, Qiu L P, Zhang L X. Challenge and Development of Sludge Treatment and Disposal in China [J]. Applied Mechanics and Materials, 2014,8,(675-677):665-668.
Wang S B. Sludge Management and Its Development in China [J]. Applied Mechanics and Materials, 2013,11,(665-668):1299- 1302.
Vega E M, Hector G O. Optimizing chemical conditioning for odour removal of undigested sewage sludge in drying processes [J]. Journal Of Environmental Management, 2015,3(150):111- 119.
Murugesan K, Selvam A. Flocculation and dewaterability of chemically enhanced primary treatment sludge by bioaugmentation with filamentous fungi [A]. International Conference on Solid Waste-Innovation in Technology and Management[C]. 2014,9(168):198-203.
Wang Y, Qu P C H, Peng D C. Hydrolysis of excess sludge by bioaugmentation and nutrient removal from supernatant with low cost [C]//Proceedings of the 2011 International Conference on Remote Sensing, Environment and Transportation Engineering, 2011,6:4096-9.
李 俊.微生物菌劑對污水處理廠污泥減量的影響研究 [D]. 重慶:西南大學, 2008.
張海珍.MCMP-II多功能復合微生物制劑用于城市污水處理廠污泥減量的生產性試驗研究 [D]. 重慶:重慶大學, 2008.
黃黎明.微生物制劑IBC強化SBR處理系統的研究 [D]. 廣州:華南理工大學, 2012.
Muyzer G, Dewaal EC, Uitterlinden AG. Profiling of complex microbial-populations by denaturing gradient gel-electrophoresis analysis of polymerase chain reaction-amplified genes-coding for 16S ribosomal-rna [J]. Applied And Environmental Microbiology, 1993,59(3):695-700.
趙 鑫,劉一威,李曉東,等.微生物復合菌劑法污泥原位減量群落動態研究 [J]. 哈爾濱工程大學學報, 2015,36(3):839-845.
薛圓圓,孫寶盛,杜 江,等.貧營養條件下IAMBR污泥微生物群落結構的演變 [J]. 中國環境科學, 2015,35(3):839-845.
陳 燕,劉國華,范 強,等.不同溶解氧條件下A/O系統的除碳脫氮效果和細菌群落結構變化環境科學, 2015,36(7): 2610-2616.
Reuter J A, Spacek D V, Snyder M P. High-Throughput Sequencing Technologies [J]. Molecular Cell, 2015,3(58):586- 597.
Picard C, Fischer A l. Contribution of high-throughput DNA sequencing to the study of primary immunodeficiencies [J]. European Journal of Immunology, 2014,10(44):2854-2861.
Georgiou G, Ippolito G C. Beausang J. The promise and challenge of high-throughput sequencing of the antibody repertoire [J]. Nature Biotechnology, 2014,2,(32):158-168.
McAdam P R, Richardson E J, Fitzgerald J. High-throughput sequencing for the study of bacterial pathogen biology [J]. Current Opinion in Microbiology, 2014,6,(19):106-113.
佳,宋云龍,張金松,等.基于微生物強化的原位污泥減量中試研究 [J]. 給水排水, 2014,40(8):37-43.
GB18918-2002 城鎮污水處理廠污染物排放標準 [S].
Turnbaugh P J, Hamady M, Yatsunenko T, et al. A core gut microbiome in obese and lean twins [J]. Nature, 2009,457(7228): 480–487.
Simpson E H. Measurement of diversity [J]. Nature, 1949, 163:688.
Hill M O. Diversity and evenness: a unifying notation and its consequences [J]. Ecology, 1973,54:427-431.
Pielou E C. The measurement of diversity in different types of biological collections [J]. Journal of Theoretical Biology, 1966, 13:131-144.
Zhang, Y X, Vadlani P V. Lactic acid production from biomass-derived sugars via co-fermentation of Lactobacillus brevis and Lactobacillus plantarum [J]. Journal of Bioscience and Bioengineering, 2015,6(119):694-699.
Vellingiri K, Ramachandran T. Isolation and Purification of Lactobacillus Acidophilus and Analyzing its Influence on Effluent Treatment [J]. International Journal of Engineering and Technology Innovation, 2015,1(5):66-74.
Oh M H, Lee, S M, Hong S H. Monitoring of Lactobacillus sp inoculated in the reactor to evaluate the solubilization efficiency of primary sludge [J]. International Biodeterioration & Biodegradation, 2013,11(85):603-607.
Demler M, Weuster-Botz D. Reaction Engineering Analysis of Hydrogenotrophic Production of Acetic Acid by Acetobacterium woodii [J]. Biotechnology and Bioengineering, 2011,2(108):470- 474.
YIN S, LI S. Ethanol-tolerant acetogenic microorganism composite bacteria used for anaerobic fermentation of medicine residue alcohol comprises Acetobacterium carbinolicum, Clostridium irregularic, Clostridium cellobioparum, and Clostridium glycolicum: China, 103555638-A [P].
Fan H, Qu H, Zhang L. Complex microbial agent useful for treating wastewater, contains Rhodopseudomonas palustris, Acetobacterium, Bacillus subtilis, Metallosphaera sedula, Metallosphaera prunae, Nitrosomonas, Lactococcus, and Flavobacterium: China, 104195090-A [P].
* 責任作者, 教授, zhangjinsong@waterchina.com
Analysis of microbial community in in-situ sludge reduction process by bioaugmentation using high-throughput sequencing technology
SONG Yun-long1, ZHANG Jin-song1*, ZHU Jia2, SHAO Ming-fei1, DENG Ren-jian1
(1.School of Civil and Environment Engineering, Harbin Institute of Technology Shenzhen Graduate School, Shenzhen 518055, China;2.School of Civil and Environmental Engineering, Shenzhen Polytechnic, Shenzhen 518055, China). China Environmental Science, 2016,36(7):2099~2107
High-throughput sequencing was used to parse the microbial community of activated sludge in an in-situ sludge reduction process. Microbial community structure shifted dramatically as the result of bioaugmentation. The abundance of and increased significantly in bioaugmentation group. Community diversity indices including Simpson index, Shannon index and Pielou index rised at different degrees in anaerobic zone of bioaugmentation group. Cluster analysis indicated that microbial communities were obviously grouped into three clusters in chronological order. In anaerobic zone of bioaugmentation group, the community structure shifted more obviously over time. Through principal coordinate analysis, communities of bioaugmentation group and control group were grouped into two distinct areas. Synergistic efforts of bioaugmentation and DO reduction were the driving forces of community shift in bioaugmentation group, while DO reduction was the driving force in control group. Canonical correspondence analysis showed that pH, water temperature and DO were the dominant environmental factors, which affected the microbial community structure significantly.
high-throughput sequencing;bioaugmentation;hydrolytic acidification;microbial community analysis
X172
A
1000-6923(2016)07-2099-09
宋云龍(1983-),男,山東威海人,哈爾濱工業大學博士研究生,主要從事污泥減量及其資源化利用的研究.
2015-11-06
國家高技術研究發展計劃(863計劃)項目(2009AA064704)
