畢赤酵母表達重組牛胰臟羧肽酶A的響應面法優(yōu)化
2016-10-13 13:20:56王曉云徐詩涵梁志宏黃昆侖
生物技術(shù)通報 2016年3期
關(guān)鍵詞:優(yōu)化
王曉云 徐詩涵 梁志宏 黃昆侖
(中國農(nóng)業(yè)大學食品科學與營養(yǎng)工程學院,北京 100083)
畢赤酵母表達重組牛胰臟羧肽酶A的響應面法優(yōu)化
王曉云 徐詩涵 梁志宏 黃昆侖
(中國農(nóng)業(yè)大學食品科學與營養(yǎng)工程學院,北京 100083)
采用Central Composite Design(CCD)實驗設計及響應面分析相結(jié)合的方法,對畢赤酵母表達牛胰臟羧肽酶A的表達條件進行優(yōu)化,旨在獲得牛胰臟羧肽酶A的最佳表達條件。單因素實驗結(jié)果顯示,誘導時間為96 h時,最佳甲醇含量為0.5%、最佳pH值為4、誘導時最佳菌體OD600值為7。在此基礎(chǔ)上,利用CCD實驗設計及響應面分析法進行回歸分析,獲得最優(yōu)表達參數(shù)為:甲醇含量0.5%、誘導pH為5.90、接種前種子培養(yǎng)基OD600為6.54。在優(yōu)化后的條件下進行驗證實驗,得到牛胰臟羧肽酶A的表達量為0.325 mg/mL,與理論值(0.326 mg/mL)相比,相對誤差較小,為0.38%。同時,對表達產(chǎn)物進行了Western Blot鑒定。研究表明,采用響應面法分析優(yōu)化牛胰臟羧肽酶A的表達條件準確可靠,有助于找出最佳條件,為指導其在發(fā)酵罐中進行高密度發(fā)酵生產(chǎn)牛胰臟羧肽酶A奠定基礎(chǔ)。
牛胰臟羧肽酶A;畢赤酵母;響應面
羧肽酶(carboxypeptidases,CPs)是一種專一性地從肽鏈的C端逐個降解、釋放游離氨基酸的一類肽鏈外切酶。主要分為絲氨酸羧肽酶(EC 3.4.16-)、金屬羧肽酶(EC 3.4.17-)和半胱氨酸羧肽酶(EC 3.4.18-)3個亞類。羧肽酶廣泛應用于醫(yī)藥、食品等工業(yè)領(lǐng)域。在食品工業(yè),可用于制備高F值寡肽[1],食品和飼料中赭曲霉素的去除[2]、用作脫苦味劑[3]等。動物來源的羧肽酶主要存在于豬、牛等的胰臟中,如羧肽酶A/B(carboxypeptidaseA/B),其數(shù)量非常有限、價格昂貴、導致其應用受到限制。微生物來源的羧肽酶存在于酵母、曲霉等真菌的液泡中,具有廣闊的應用前景,但后期純化及大量獲得仍然存在困難。因此,借助基因工程策略采用微生物為宿主大量生產(chǎn)重組羧肽酶,有助于進一步降低生產(chǎn)成本、提高產(chǎn)品質(zhì)量、深化酶學性質(zhì)研究、擴展應用范圍。趙瑩[4]完成了豬羧肽酶A1酶原在畢赤酵母中的表達,易靜等[5]完成了人羧肽酶A1的畢赤酵母可溶表達,為進一步研究CPA1 在抗體導向酶-前體藥物療法中的應用奠定了基礎(chǔ)。
赭曲霉毒素A(Ochratoxin A,OTA)是由曲霉屬(Aspergillus spp.)和青霉屬(Penicillium spp.)等霉菌產(chǎn)生的次級代謝產(chǎn)物,屬于聚酮類化合物,主要存在于谷物及其副產(chǎn)品中[6]。OTA具有強烈的腎臟毒性和肝臟毒性,并有致畸、致突變和致癌作用[7],對動物和人體健康有很大的潛在危害,研究OTA的脫毒對人類健康生活至關(guān)重要。1969年P(guān)itiou[8]用薄層層析和光譜方法就發(fā)現(xiàn)羧肽酶(CPA)能將赭曲霉毒素A降解為OTα和L-苯丙氨酸。Deberghes[9]在含有赭曲霉的液體培養(yǎng)基(50 mL)中分別加入100 μL的1、2、5 U的牛胰臟羧肽酶A,結(jié)果發(fā)現(xiàn)1 U羧肽酶A降解率為17%,2 U羧肽酶A降解率為49%,5 U羧肽酶A降解率達了100%。本實驗所用菌由師磊等[10]證明具有降解赭曲霉毒素A效果。
巴斯德畢赤酵母表達系統(tǒng)是應用廣泛而成功的蛋白表達系統(tǒng)。與其他真核蛋白及原核蛋白表達系統(tǒng)相比,具有以下優(yōu)點[11]:(1)生長速度快,易實現(xiàn)高密度發(fā)酵;(2)可生產(chǎn)分泌型蛋白,方便收集及純化;(3)可避免內(nèi)毒素及噬菌體污染;(4)可進行蛋白的翻譯后修飾,包括多肽的折疊、糖基化、甲基化等;(5)有良好的酵母表達載體,如pPIC 9K、pPICZα、pMET B等。外源蛋白的表達條件如誘導時間、pH、碳源、誘導劑濃度等對目標蛋白的產(chǎn)量及活性影響較大。優(yōu)化培養(yǎng)條件可有效提高外源蛋白含量。
響應面法(Response surface methodology,RSM)是以多元二次回歸數(shù)學模型為工具,描述相互作用的實驗因子與響應值之間的關(guān)系的數(shù)據(jù)處理方法。響應面法能有效地找到因素間的最優(yōu)組合,相比于傳統(tǒng)的全因子實驗設計,該法能在更為經(jīng)濟的試驗次數(shù)下,得到精確的統(tǒng)計結(jié)果,具有周期短、精確度高等優(yōu)點。因此,本研究利用單因素實驗,初步優(yōu)化了培養(yǎng)基成分,誘導pH值、誘導前菌體OD600值和甲醇含量。在單因素實驗的基礎(chǔ)上,利用響應面法優(yōu)化實驗對初始誘導pH值、誘導前菌體OD600值和甲醇含量三因素對牛胰臟羧肽酶A表達量的影響進行研究,分析各因素的變化規(guī)律,以期獲得牛胰臟羧肽酶A表達的最佳條件。
1 材料與方法
1.1 材料
重組巴斯德畢赤酵母Pichia pastoris(GSll5/ pPIC9K/proCPA,His+,Mut+,本實驗室構(gòu)建、保存(牛胰臟ProCPA基因(Accession No.NM_174750)由北京奧科鼎盛有限公司依據(jù)P.pastoris對密碼子的偏愛性進行密碼子優(yōu)化);Werstern Blot 實驗中的His 標簽小鼠單克隆抗體、AP標記的山羊抗小鼠IgG抗體均購自Abcam;BCIP/NBP顯色液購自Amresco;培養(yǎng)基試劑購自北京奧博星生物技術(shù)有限責任公司;其余試劑均為分析純。
1.2 方法
1.2.1 菌株活化 將畢赤酵母重組菌株在新鮮YPD平板上劃線,于30℃倒置培養(yǎng)2 d。
1.2.2 種子培養(yǎng) 從YPD平板挑取單菌落接種到50 mLYPD培養(yǎng)基中,30℃和200 r/min 振蕩培養(yǎng)過夜,然后接入50 mL BMGY培養(yǎng)基中繼續(xù)培養(yǎng)至OD600達6.0-8.0。
1.2.3 牛proCPA基因的表達 250 μL種子培養(yǎng)液接種于25 mL BMGY(13. 4 g /L YNB,4 ×10-5g /L生物素,100 mL /L 磷酸緩沖液,10 mL /L 甘油)培養(yǎng)液中。于28℃,250 r /min 的速度振搖至菌濃度為所需的OD600。離心收集菌體棄去BMGY培養(yǎng)液,用25 mL 改良BMMY(13. 4 g /L YNB,4 ×10-5g /L生物素,100 mL /L 磷酸緩沖液,1% 山梨醇,所需的甲醇濃度)培養(yǎng)液。于28℃,250 r /min 的速度振搖,開始誘導表達,每12 h或24 h 補充甲醇。于24、48、72、96、120 h 分別取1 mL 上清液。
1.2.4 發(fā)酵產(chǎn)物分析 每天收集1 mL的發(fā)酵上清液,測量OD值,4℃,10 000 r/min離心5 min,取出上清,-20℃保存,備檢。
1.2.5 表達上清檢測 對上清中的蛋白進行SDSPAGE、Western Blot驗證及純化。利用Bradford法,結(jié)合標準曲線對表達上清中的蛋白含量進行分析。980 μL考馬斯亮藍G250中加入20 μL的表達上清,充分混勻。室溫反應2 min,測定595 nm處的吸光值。依據(jù)標準曲線,計算蛋白濃度。
SDS-PAGE 分析:配制12% 分離膠和5% 濃縮膠,取上述樣品40 μL,加入10 μL 5×Loading Buffer,混合制樣。每孔上樣30 μL,濃縮膠電壓80 V,分離膠電壓120 V。電泳結(jié)束后,ddH2O沖洗后,在考馬斯亮藍染色液中振蕩染色40 min,然后用ddH2O 反復沖洗,放入脫色液(10% 的乙酸)中脫色,直至背景清晰,此過程需不定時更換脫色液。
Western Blot 驗證:將SDS-PAGE電泳結(jié)束后未染色的膠,電轉(zhuǎn)至NC膜上;加入封閉液(5%的BSA溶于TBST中)室溫振蕩孵育1 h;TBST振蕩洗滌6次,每次5 min;加入TBST溶液稀釋的His標簽的鼠源一抗,4℃孵育過夜(12-16 h);TBST振蕩洗滌6次,每次5 min;加入TBST溶液稀釋的AP標記的羊抗鼠二抗,室溫振蕩孵育1 h;TBST振蕩洗滌6次,每次5 min;加入BCIP/NBP顯色液,避光顯色5-10 min,肉眼觀察結(jié)果。
2 結(jié)果
2.1 目的蛋白SDS-PAGE、Western Blot驗證及純化
含重組質(zhì)粒(pPIC9K/proCPA,His+,Mut+)的巴斯德畢赤酵母GS115菌株經(jīng)甲醇誘導后在47 kD左右有明顯的蛋白條帶(圖1),與目的蛋白大小符合。經(jīng)Western Blot分析為目的蛋白(圖2)。
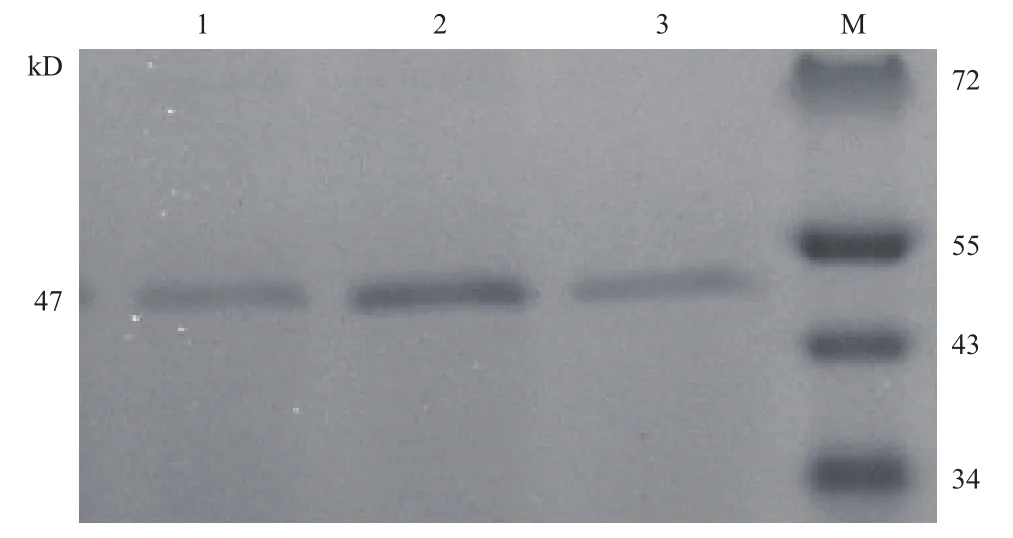
圖1 目的蛋白的SDS-PAGE驗證
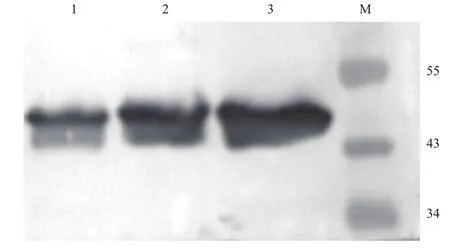
圖2 目的蛋白的Western Blot驗證
2.2 標準曲線
用BSA制作蛋白標準液,根據(jù)不同蛋白濃度繪制蛋白濃度(mg/mL)-吸光度A的標準曲線。經(jīng)數(shù)據(jù)處理分析獲得蛋白質(zhì)標準曲線回歸方程為:y=0.4837x+0.2068,R2=0.9931。
2.3 單因素實驗結(jié)果
2.3.1 甲醇誘導濃度對蛋白表達量的影響 考察甲醇含量分別為1% 每24 h補加、0.5% 甲醇每24 h補加、1% 甲醇每12 h補加時蛋白的表達量。當甲醇濃度為0.5% 時,蛋白濃度最大,且在第4天后漲勢緩慢,第6天時濃度最大(圖3-A)。考慮到工業(yè)生產(chǎn)的時間成本問題,誘導表達時間為4 d。
2.3.2 誘導pH值對蛋白表達量的影響 考察誘導pH分別為3、4、5、6時目的蛋白的表達量。發(fā)現(xiàn)誘導pH值為4時第4天目的蛋白表達量均較高(圖3-B)。因此,單因素實驗中的最適誘導pH值為4。
2.3.3 誘導時菌體OD600對蛋白表達量的影響 考察誘導時菌體OD600值分別為6、7、8時目的蛋白的表達量。誘導時菌體OD600值為7時目的蛋白表達量最高(圖3-C)。因此,選擇菌體OD600值為7作為誘導表達時的最適的菌體OD600值。
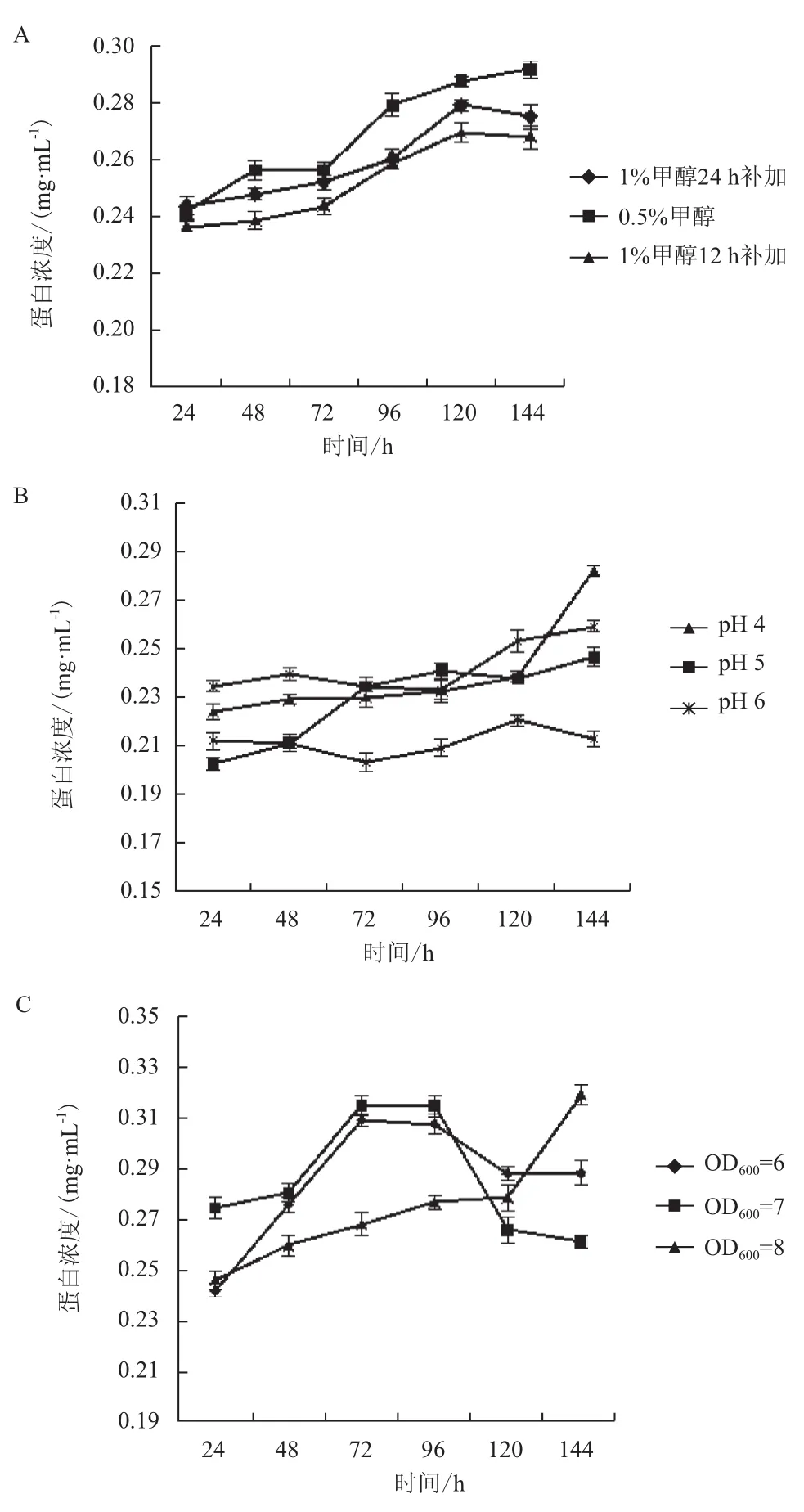
圖3 甲醇濃度(A)、初始誘導pH值(B)、及誘導時菌體OD600(C)對蛋白表達量的影響
2.4 響應面法實驗設計
根據(jù)CCD實驗設計原理,綜合單因素影響實驗結(jié)果,以誘導pH值(pH)、誘導表達前培養(yǎng)基OD600(OD)和甲醇添加量(M)三因素為自變量,以牛胰臟羧肽酶A表達量為響應值,利用CCD組合設計實驗方案,并以-1、0和1 分別代表變量的因素編碼(表1),進行畢赤酵母菌表達牛胰臟羧肽酶A的優(yōu)化。采用Design Expert 8.0.6軟件進行實驗設計。
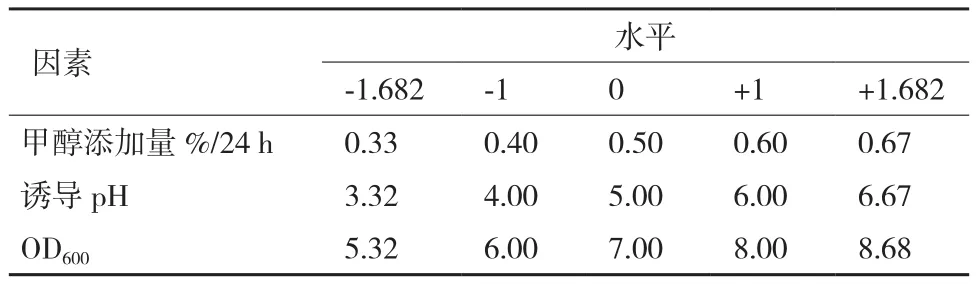
表1 響應面旋轉(zhuǎn)中心組合設計因素水平編碼表
2.5 模型方程的建立及顯著性檢驗
根據(jù)表2 結(jié)果計算各項回歸系數(shù),以這些回歸系數(shù)建立牛胰臟羧肽酶A表達量與誘導pH值(pH)、誘導表達前培養(yǎng)基OD600(OD)和甲醇添加量(M)三個因素的數(shù)學回歸模型。由表2 可以得出二次回歸方程式為:牛胰臟羧肽酶A表達量= -1.23998+3.07479×M+0.18464×pH-0.082985×OD-0.055525×M×pH-0.042232×M×OD-3.15950E-003×pH×OD-2.5376×M2-0.011486×pH2-3.4090E-003×OD2;M、pH 變量的正系數(shù)表明,該變量的正向變化能引起響應值的增加;OD變量負系數(shù)表明,該變量的正向變化能引起響應值的減少。負的二次項系數(shù)表明,方程的拋物面開口向下,具有極大值點,能夠進行最優(yōu)分析。
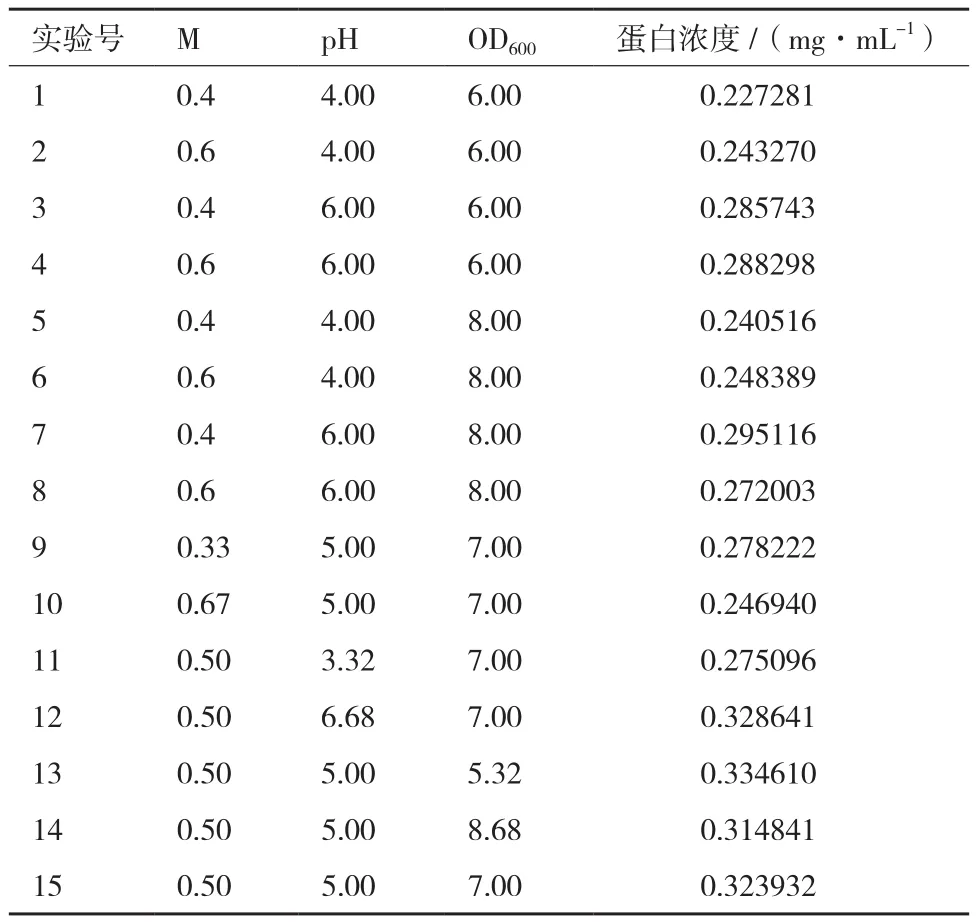
表2 響應面旋轉(zhuǎn)中心組合設計與結(jié)果
回歸模型方差分析(表3)顯示,實驗選用的模型顯著(P<0.05),說明模型是合適的;方差分析中的確定系數(shù)R2=0.801 6,校正確定系數(shù)R2Adj=0.6229。R2Adj可顯示所選模型的擬合程度,即預測值與實測值之間的相關(guān)性,Jalgeker和May建議—個擬合較好的模型R2至少應為0.80[12]。
由表3 可知模型的一次項B影響顯著,A,C影響不顯著。從F 值可以看出,單因素對提取率的影響順序依次是B>A>C,即pH>甲醇添加量>OD。交互項影響不顯著,二次項A2影響極顯著,表明該影響因素對表達量的影響不是簡單的線性關(guān)系。
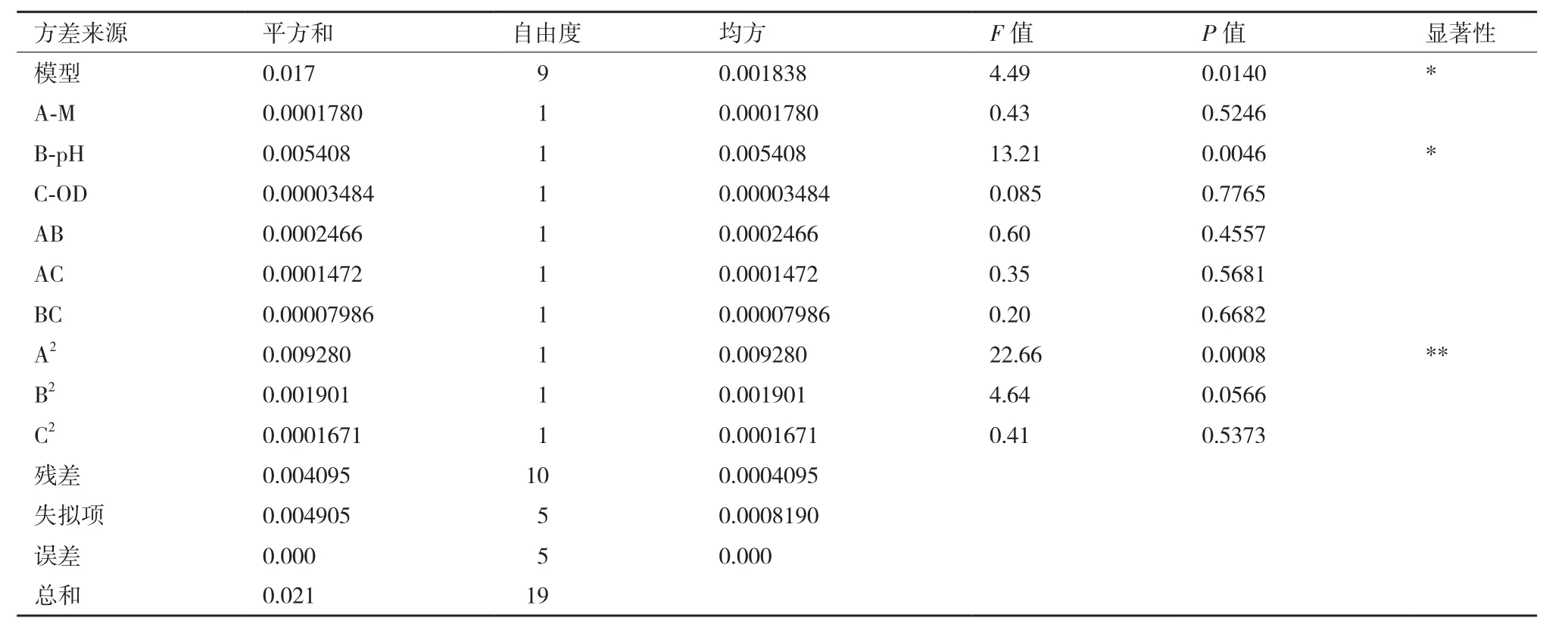
表3 回歸模型方差分析
2.6 響應面交互作用分析與工藝優(yōu)化
對誘導pH值(pH)、誘導表達前培養(yǎng)基OD600(OD)和甲醇添加量(M)三因子,兩兩作交互作用分析,對其作響應面圖,分別見圖4-圖6。等高線的形狀反映交互效應的強弱大小,圓形表示兩因素交互作用不顯著,而橢圓形則表示兩因素交互作用顯著。3張圖中等高線均呈橢圓形,故3個因素兩兩之間交互作用顯著,其中交互作用顯著程度初始誘導pH值與甲醇添加量<誘導pH值與誘導表達前培養(yǎng)基OD600<甲醇添加量與誘導表達前培養(yǎng)基OD600。在求取最優(yōu)提取條件時RSM 能夠綜合考慮各因素對2 個響應值不同的影響程度及趨勢進行綜合評價。可得最佳工藝參數(shù)為:甲醇每24 h含量0.5%、誘導pH為5.90、誘導表達前培養(yǎng)基OD600為6.54,在此優(yōu)化條件下,牛胰臟羧肽酶A蛋白表達量為0.326 mg/mL。為了進一步驗證響應面分析法的可靠性,采用上述最優(yōu)條件進行蛋白表達,3組平行,測得牛胰臟羧肽酶A蛋白表達量分別為:0.324 mg/mL、0.324 mg/mL和0.326 mg/mL,取平均值為0.325 mg/mL,與理論值相比,相對誤差較小,為0.38%。因此,采用響應面法分析優(yōu)化得到的牛胰臟羧肽酶A的參數(shù)準確可靠,可用于實際操作。
3 討論
本研究利用畢赤酵母表達系統(tǒng)成功實現(xiàn)了牛胰臟羧肽酶A的可溶性表達及純化,對表達產(chǎn)物進行了SDS-PAGE及Western Blot驗證。通過單因素及響應面法優(yōu)化了表達條件。羧肽酶作為一大類水解酶,應用及其廣泛,而其來源有限、價格昂貴使其應用受到了限制。借助基因工程手段可以實現(xiàn)羧肽酶的大量生產(chǎn),有助于擺脫來源有限,價格昂貴的限制。畢赤酵母表達系統(tǒng)是一種應用成熟的表達系統(tǒng),其分泌型表達使表達產(chǎn)物實現(xiàn)可溶狀態(tài),廣泛用于各類酶的表達。Damaso等[13]在畢赤酵母中優(yōu)化表達了耐熱木聚糖,Minning等[14]在畢赤酵母中優(yōu)化表達了米根霉脂肪酶。
外源蛋白的表達量受到許多因素的調(diào)節(jié)。由于畢赤酵母對所使用的氨基酸密碼子具有明顯的偏愛性,同義密碼子的表達效率差別明顯[15]。故在表達外源蛋白時,應進行密碼子優(yōu)化和GC含量的調(diào)整[16]。本研究中所使用的表達載體在構(gòu)建時已進行了密碼子的優(yōu)化,且表達量有顯著提高。宿主菌的選擇也至關(guān)重要。目前,用于外源基因表達的菌株種類眾多,如GS115,X-33,KM71 等。為了防止外源蛋白被同時表達的蛋白酶酶解,蛋白酶缺陷型菌株SMD1168(pep4-)、SMD1165(prb1-)和SMD1163(pep4-prb1-)也被廣泛使用[17]。此外,表達載體的選擇,表達載體導入酵母菌基因組,多拷貝菌株的篩選,信號肽的選擇和改造也至關(guān)重要。
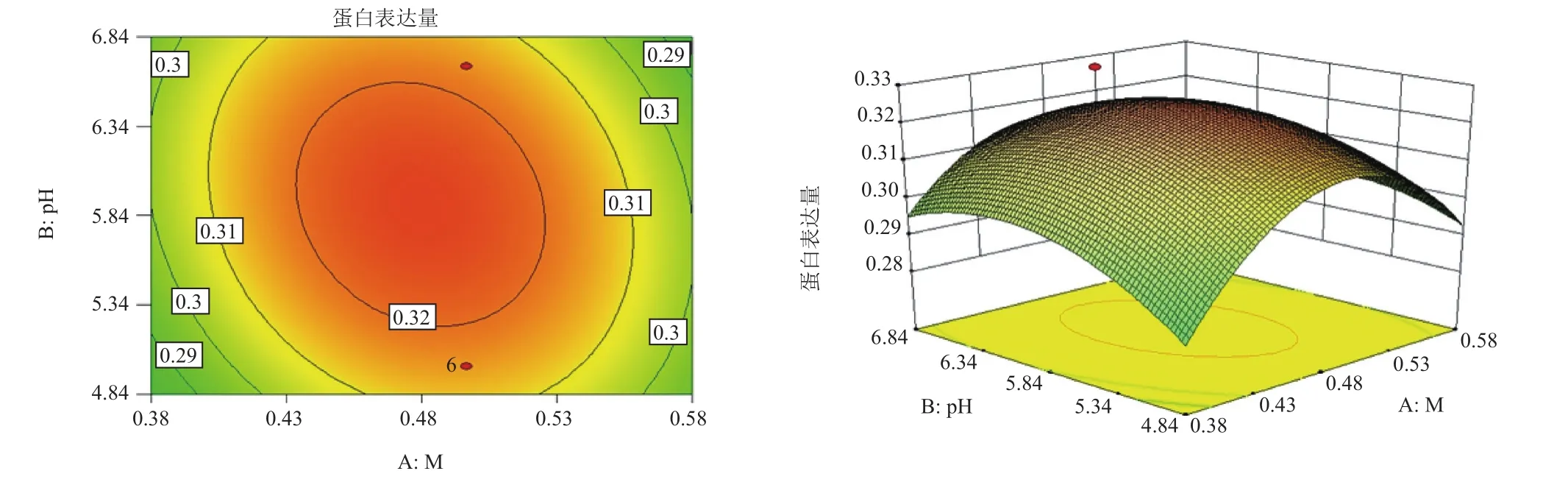
圖4 甲醇添加量和誘導pH值的響應曲面和等高線圖
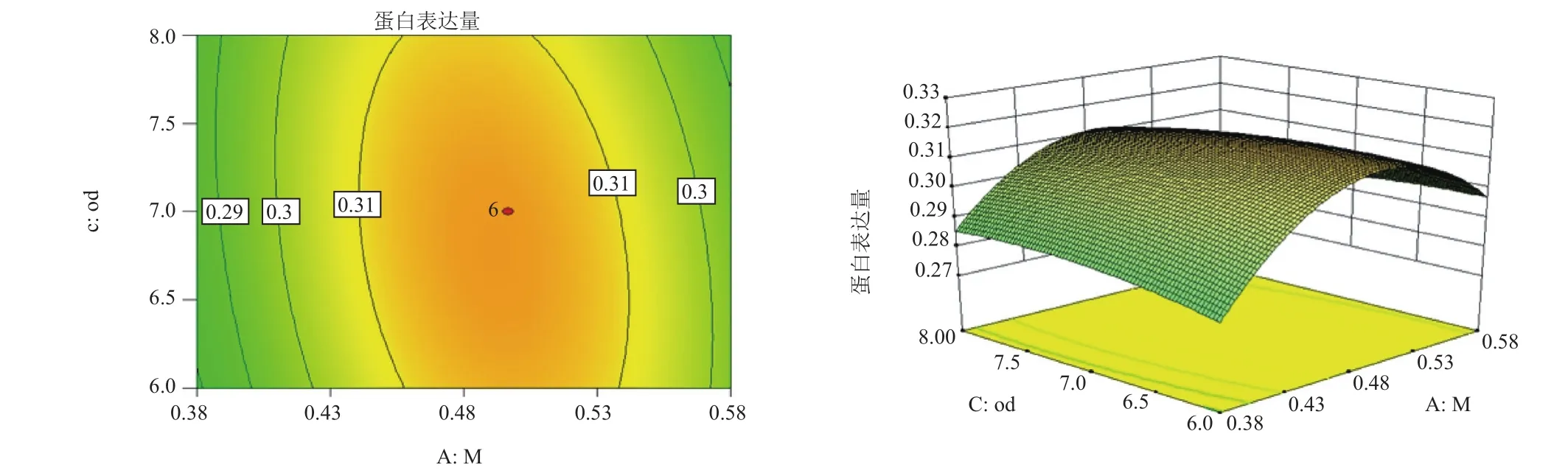
圖5 甲醇添加量和誘導表達前培養(yǎng)基OD600的響應曲面和等高線圖
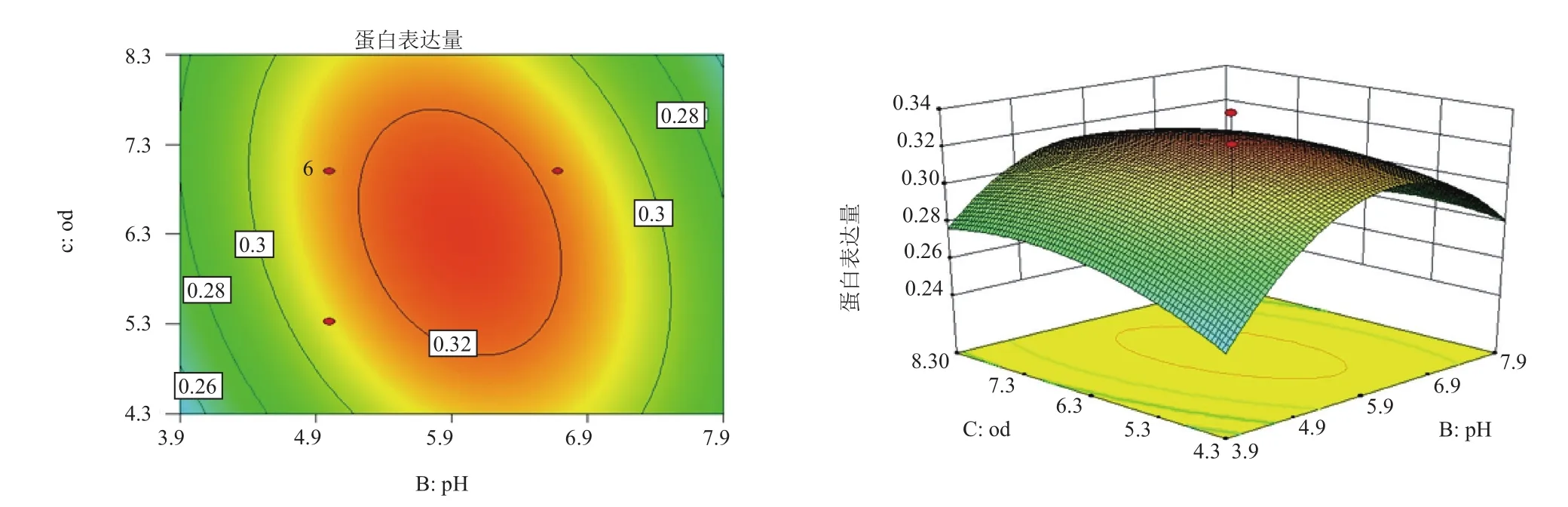
圖6 誘導pH值和誘導表達前培養(yǎng)基OD600的響應曲面和等高線圖
發(fā)酵條件的優(yōu)化,如溶氧、pH、溫度、甲醇濃度、碳源、誘導時間等都是影響蛋白產(chǎn)量的重要條件。pH 值影響細胞的生物量、蛋白表達量、活性及穩(wěn)定性。在pH5.0 時,重組人生長激素的生物量為42 g/L,蛋白表達量為271 mg/L,而當培養(yǎng)基pH 達6.0 時,雖然細胞生物量最高可達53 g/L,但是蛋白表達量僅為24 mg/L[18]。低溫誘導或許可以通過降低蛋白酶活力,增加外源蛋白穩(wěn)定性來提高外源蛋白的產(chǎn)量。合適的甲醇濃度是維持細胞高活性,促進蛋白高表達的關(guān)鍵因素。過低的甲醇濃度不足以完全誘導蛋白的表達,而當甲醇濃度過高時又會對細胞產(chǎn)生毒性,一般超過1%的甲醇濃度就會對酵母細胞產(chǎn)性毒性。本研究中,甲醇濃度為0.5%,每天補加培養(yǎng)基體積0.5%的純甲醇時即可誘導目的蛋白表達,且表達量高于1%的甲醇濃度。另外,當使用甲醇作為唯一碳源時,細胞生長緩慢,混合碳源補加的方法能有效提高表達量。常用的混合碳源有甲醇和山梨醇,甲醇和甘油等。Diego等[19]在培養(yǎng)基中添加山梨醇顯著提高了牛凝乳酶B的表達量。山梨醇的添加有助于降低耗氧量及熱產(chǎn)率[20]。本研究中發(fā)現(xiàn)添加山梨醇可以增加蛋白表達量,故在添加1%山梨醇的BMMY培養(yǎng)基中進行優(yōu)化實驗。一般來說,表達體系中細胞密度增加,蛋白表達量也會相應升高。但較高的細胞密度,溶氧量、營養(yǎng)成分,酵母細胞的代謝產(chǎn)物會影響目的蛋白的產(chǎn)量和穩(wěn)定性。誘導前培養(yǎng)基的OD值表示了酵母細胞的生物量,本研究中,誘導表達前菌體的OD600值分別為6、7、8,當OD600的值為7時,第1天的蛋白表達量明顯高于其他兩組,且在前4天一直高于其他兩組。
4 結(jié)論
本研究采用CCD實驗設計原理設計及響應面分析,建立牛胰臟羧肽酶A表達量的二次多項式數(shù)學模型。經(jīng)檢驗證明該模型是合理可靠的,能夠較好地預測牛胰臟羧肽酶A表達量。
利用模型的響應面及其等高線,對影響牛胰臟羧肽酶A表達量的關(guān)鍵因素及其相互作用進行探討,通過典型分析并考慮到實際操作的便利性得到的最終優(yōu)化條件參數(shù)為甲醇每24 h含量0.5%、誘導pH為5.90、誘導表達前培養(yǎng)基OD600為6.54。同時,在此優(yōu)化條件下,牛胰臟羧肽酶A表達量的理論值為0.326 mg/mL。
采用上述最優(yōu)條件進行蛋白表達,3組平行,測得牛胰臟羧肽酶A蛋白表達量分別為0.324 mg/ mL、0.324 mg/mL和0.326 mg/mL,取平均值為0.325 mg/mL,與理論值相比,相對誤差較小,為0.38%。因此,采用響應面法分析優(yōu)化得到的牛胰臟羧肽酶A的參數(shù)準確可靠,獲得了較高的蛋白表達量。
[1]吳靜, 閔柔, 鄔敏辰, 等. 羧肽酶研究進展[J]. 食品與生物技術(shù)學報, 2012, 31(8):793-801.
[2]Abrunhosa L, Santos L, Venancio A. Degradation of ochratoxin A by proteases and by a crude enzyme of Aspergillus niger[J]. Food Biotechnol, 2006, 20:231-242.
[3]馮紅霞. 羧肽酶菌株的篩選及其酶的分離純化、酶學性質(zhì)的研究[D]. 南京:南京農(nóng)業(yè)大學, 2002.
[4]趙瑩. 豬羧肽酶 A1 酶原的基因克隆及其在畢赤酵母中的表達[D]. 成都:四川農(nóng)業(yè)大學, 2009.
[5]易靜, 郝曉柯, 蘇明權(quán), 等. 人羧肽酶A1活性中心的畢赤酵母可溶表達[J]. 第四軍醫(yī)大學學報, 2006 27(5):389-391.
[6]Doreen D, Huaming W, Gunter S, et al. Structural and functional characterization of ochratoxinase, a novel mycotoxin-degrading enzyme[J]. Biochemistry Journal, 2014, 462:441-452.
[7]Zhihong L, Kunlun H, Yunbo L. Ochratoxin A and ochratoxinproducing fungi on cereal grain in China:a review[J]. Food Additives & Contaminants:Part A, 2015, 32(4):461-470.
[8]Pitout, M. J. The hydrolysis of ochratoxin A by some proteolytic enzymes[J]. Biochemical Harmacology, 1969, 18:485-489.
[9]Deberghes P, Betbeder AM, Boisard F, et al. Detoxification of ochratoxin A, a food contaminant:prevention of growth of Aspergillus ochraceus and its production of ochratoxin A[J]. Mycotoxin Researsh, 1995, 11:37-47.
[10]師磊. 兩柱芽孢桿菌和重組羧肽酶A對赭曲霉毒素A的脫毒研究[D]. 北京:中國農(nóng)業(yè)大學, 2014.
[11]Li PZ, Anumanthan A, Gao XG, et al. Expression of recombinant proteins in Pichia Pastoris[J]. Applied Biochemistry and Biotechnology, 2007, 142:105-124.
[12]Jodekar AM, May AT. Product exceuence through design of experiments[J]. Cereal Foods World, 1987, 32(12):857—868.
[13] Damaso MC, Almeida MS, Kurcenbach E, et al. Optimized expression of a thermostable xylanase from Thermomyces lanuginosus in Pichia pastoris[J]. Applied and Environment Microbiology, 2003,69(10):6064-6072.
[14]Minning S, Serrano A, Ferrer P, et al. Optimization of the high-level production of Rhizopus oryzae lipase in Pichia pastoris[J]. J Biotechnol, 2001, 86:59-70.
[15] 陸永超, 蔣琳. 畢赤酵母高效表達策略概述[J]. 微生物學免疫學進展, 2013, 41(1):70-76.
[16] 盧其湘, 趙樹進. 外源基因在畢赤酵母中表達的優(yōu)化[J]. 氨基酸和生物資源, 2005, 27(4):27-29.
[17] Joan LC, James MC. Heterologous protein expression in the methylotrophic yeast Pichia pastoris[J]. FEMS Microbiology Reviews, 2000, 24:45-66.
[18]Bayraktar E. Effects of pH on human growth hormone production by Pichia pastoris considering the expression levels of regulatory genes[D]. Ankara:Middle East Technical University, 2009.
[19]Diego GN, Matías NR, Martín B, et al. Cloning, expression and optimized production in a bioreactor of bovine chymosin B in Pichia(Komagataella)pastoris under AOX1 promoter[J]. Protein Expression and Purification, 2013, 92:235-244.
[20]Jungo C, Schenk J, Pasquier M, et al. A quantitative analysis of the benefits of mixed feeds of sorbitol and methanol for the production of recombinant avidin with Pichia pastoris[J]. Journal of Biotechnology, 2007, 131:57-66.
(責任編輯 李楠)
Optimization of Expression Conditions of Recombinant Bovine
Pancreatic Carboxypeptidase A in Pichia pastoris by Response Surface Method
WANG Xiao-yun XU Shi-han LIANG Zhi-hong HUANG Kun-lun
(College of Food Science & Nutritional Engineering,China Agricultural University,Beijing 100083)
Central composite design(CCD)and response surface methodology(RSM)were used to optimize the expression conditions of bovine pancreatic carboxypeptidase A. The results of single-factor test showed that the optimal conditions as following:the inducible expression time was 96 hours,the methanol content was 0.5% every 24 h,the initial induction pH4,and OD600before induction expression was 7. Based on the above result,CCD was used to design the tests,and RSM to have regression analysis. The optimal parameters for the expression were obtained as:methanol content was 0.5% every 24 hours,the induction pH 5.90,OD600before inducible expression was 6.54,the expression level after 96 h reached 0.325 mg/mL. Compared with the theoretical value(0.326 mg/mL),the small relative error was 0.38%. Also,the expression of the target protein was confirmed by Western blot. Conclusively,using RSM for optimizing the expression condition of bovine pancreatic carboxypeptidase A was accurate and reliable,and conducive to seek the optimal condition,thus this lays a foundation for guiding the high-density fermentation production of bovine pancreatic carboxypeptidase A in fermenter.
bovine pancreatic carboxypeptidase A;Pichia pastoris;response surface method
10.13560/j.cnki.biotech.bull.1985.2016.03.030
2015-05-14
國家高技術(shù)研究發(fā)展計劃(“863”計劃)(2012AA101609-7)
王曉云,女,碩士研究生,研究方向:微生物脫毒與食品安全;E-mail:5052806592@qq.com
梁志宏,女,博士,副教授,研究方向:微生物與食品安全;E-mail:lzh01@cau.edu.cn
猜你喜歡
房地產(chǎn)導刊(2022年5期)2022-06-01 06:20:14
能源工程(2022年1期)2022-03-29 01:06:28
建材發(fā)展導向(2021年12期)2021-07-22 08:06:48
建材發(fā)展導向(2021年7期)2021-07-16 07:07:52
中學生數(shù)理化(高中版.高二數(shù)學)(2021年12期)2021-04-26 07:43:48
中學生數(shù)理化(高中版.高考數(shù)學)(2021年12期)2021-03-08 01:28:50
今日農(nóng)業(yè)(2020年16期)2020-12-14 15:04:59
消費導刊(2018年8期)2018-05-25 13:20:08
家庭影院技術(shù)(2018年4期)2018-05-09 07:07:41
電子制作(2017年20期)2017-04-26 06:57:45
