septin基因家族的功能及其與人類疾病關(guān)系的研究進(jìn)展*
2016-11-01 08:31:31綜述孫厚良趙麗芳審校
重慶醫(yī)學(xué) 2016年25期
馬 強(qiáng) 綜述,孫厚良,趙麗芳,郝 坡△ 審校
(1.重慶三峽醫(yī)藥高等專科學(xué)校 404120;2.重慶市抗腫瘤天然藥物工程技術(shù)研究中心 404120;3.漯河醫(yī)學(xué)高等專科學(xué)校,河南漯河 462002)
?
·綜述·
septin基因家族的功能及其與人類疾病關(guān)系的研究進(jìn)展*
馬強(qiáng)1,2綜述,孫厚良1,2,趙麗芳3,郝坡1,2△審校
(1.重慶三峽醫(yī)藥高等專科學(xué)校404120;2.重慶市抗腫瘤天然藥物工程技術(shù)研究中心404120;3.漯河醫(yī)學(xué)高等專科學(xué)校,河南漯河 462002)
Septin;基因家族;分類;生理功能;人類疾病
septin是一類具有鳥苷三磷酸酶(GTPase)活性的基因家族,該家族成員在進(jìn)化上相對(duì)比較保守,各家族成員之間除了N端和C端稍有不同外,位于中央的鳥苷三磷酸(GTP)結(jié)合結(jié)構(gòu)域和多堿性氨基酸結(jié)構(gòu)域都是高度保守的[1]。此外,該基因家族的分布也較為廣泛,目前除植物外,在絕大多數(shù)的真核生物中均發(fā)現(xiàn)有septin基因的存在。并且根據(jù)組織表達(dá)和互作蛋白的不同,這些septin基因家族成員可執(zhí)行不同的細(xì)胞功能,包括細(xì)胞極性、胞質(zhì)分裂及細(xì)胞凋亡等一系列重要的生理功能。而septin基因的表達(dá)也會(huì)受到嚴(yán)格的控制,這樣可以維持纖維絲的正確組裝和正常的細(xì)胞功能。但如果是因?yàn)榛蛲蛔兒捅磉_(dá)變化引起septin蛋白改變,就會(huì)導(dǎo)致疾病的發(fā)生,目前人們發(fā)現(xiàn)大多數(shù)septin家族成員的表達(dá)變化與人類多種疾病的發(fā)生有著密切聯(lián)系,如腫瘤的發(fā)生、病原微生物感染、神經(jīng)系統(tǒng)退行性疾病及精子的發(fā)生過程等。但是由于septin 基因家族成員非常龐大,并且它們的結(jié)構(gòu)和功能都比較復(fù)雜,因此,對(duì)這些家族成員與多種疾病關(guān)系的具體作用機(jī)制尚未完全研究清楚。本文將試圖從septin 基因家族的分類、結(jié)構(gòu)、功能及與人類疾病發(fā)生、發(fā)展的關(guān)系方面對(duì)近年來的研究進(jìn)展作一綜述。
1 septin基因家族的分類及結(jié)構(gòu)
septin最早發(fā)現(xiàn)于上世紀(jì)70年代初,它是作為一種參與芽殖酵母(saccharomycescerevisiae)胞質(zhì)分裂過程所必需的蛋白質(zhì)[1]。而人類septin作為基因家族的一部分,盡管它們都具有相似的序列,但功能卻不完全相同,并且這些septin家族成員都來自于一個(gè)共同的祖先[2]。目前,已知的人類septin家族由13個(gè)基因組成(表1),它們是SEPT1~SEPT12及SEPT14,而之前人們所熟知的SEPT13基因目前已被確定是與SEPT7相關(guān)的偽基因,由于這些SEPT基因會(huì)出現(xiàn)多種亞型,因此能編碼幾十種不同的septin蛋白,其所編碼蛋白質(zhì)的相對(duì)分子質(zhì)量在(39~60)×103[2]。此外,根據(jù)系統(tǒng)發(fā)育和進(jìn)化分析的結(jié)果,人們將spetin基因家族成員又分為幾個(gè)不同的亞家族,這些亞家族成員中的SEPT2、SEPT7和SEPT9目前已經(jīng)被證實(shí)具有GTP酶活性,能夠水解鳥苷二磷酸(GDP),而SEPT6亞家族的成員不具備這種能力[3]。這也就意味著GTP水解作用的缺失是一種像人類這樣擁有數(shù)量眾多septin的生物體一個(gè)主要的特征。然而,這些研究結(jié)果背后的原因仍不清楚。GTP的結(jié)合已被證實(shí)有誘導(dǎo)結(jié)合區(qū)構(gòu)象變化的特性,并且其作用機(jī)制被認(rèn)為與Ras蛋白超家族的功能有關(guān)[1,4]。但是,septin蛋白的組裝是否需要GTP的水解及GTP結(jié)合區(qū)的構(gòu)象變化是否會(huì)影響到septin纖維微絲的形成,這些問題在很大程度上仍是個(gè)未知數(shù)。
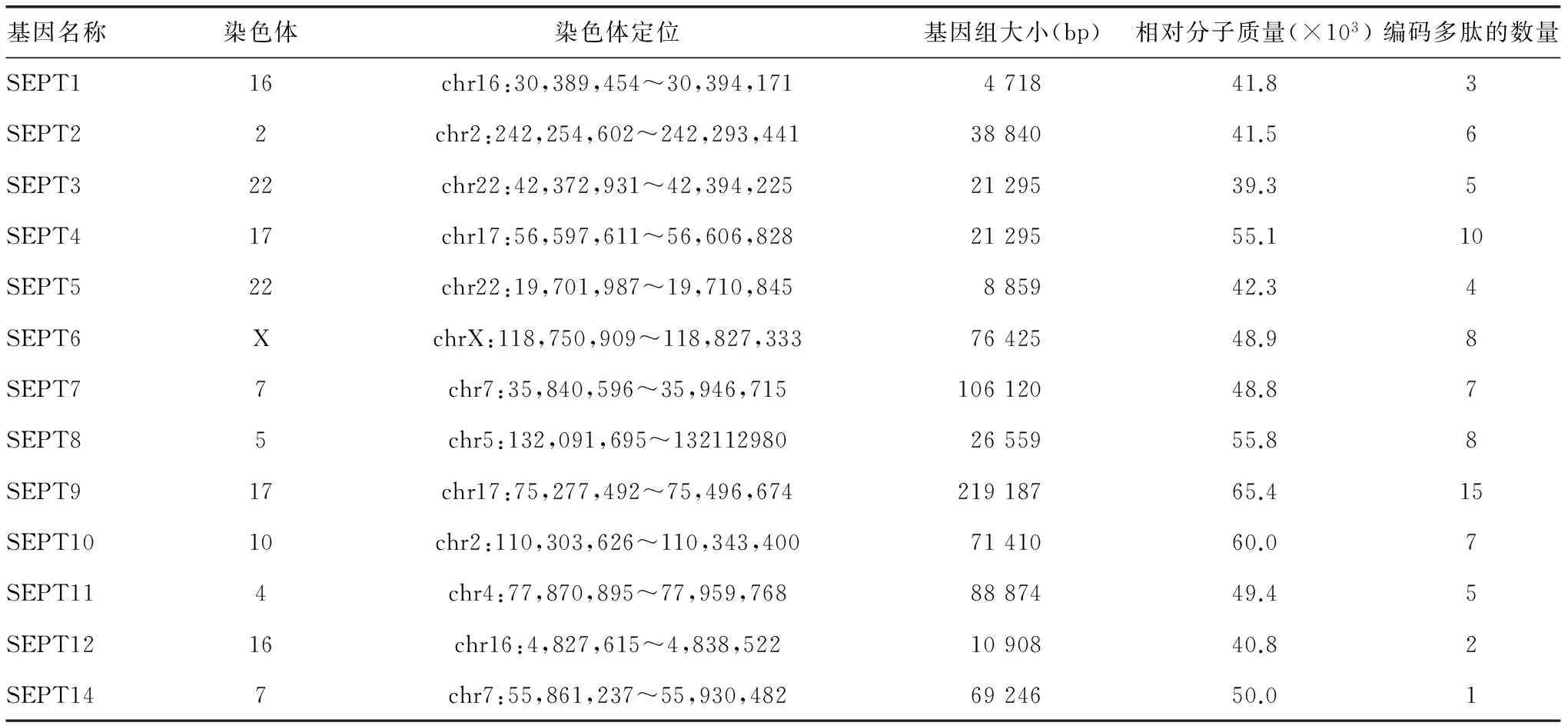
表1 人類septin基因家族成員匯總表
septin蛋白序列N端的多元區(qū)側(cè)翼有一個(gè)高度保守的GTP結(jié)合結(jié)構(gòu)域(圖1),該結(jié)構(gòu)域主要負(fù)責(zé)septin蛋白與細(xì)胞膜之間進(jìn)行互作和結(jié)合。在septin多元區(qū)的上游,含有一個(gè)富含脯氨酸的結(jié)構(gòu)域,該結(jié)構(gòu)域在參與蛋白質(zhì)與蛋白質(zhì)及蛋白質(zhì)與細(xì)胞骨架組分相互作用中顯得尤為重要[5]。此外,septin蛋白結(jié)構(gòu)復(fù)雜性的增加使得自身能同時(shí)編碼多個(gè)復(fù)合的轉(zhuǎn)錄組,并且能產(chǎn)生多種變異剪接本。在人類所有的septin基因家族成員中,除了SEPT14只能編碼一種亞型的多肽外,其余SEPT基因均能編碼2種或2種以上的多肽。最近,隨著微小核糖核酸(miRNAs)和反義轉(zhuǎn)錄本被注釋到septin的基因組區(qū)域,意味著這種嚴(yán)格的轉(zhuǎn)錄調(diào)控機(jī)制可能會(huì)協(xié)調(diào)這個(gè)復(fù)雜基因家族的蛋白表達(dá)。
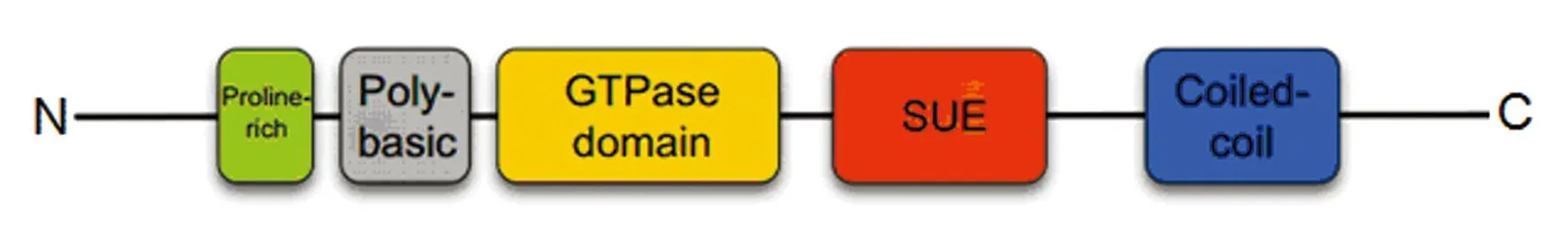
綠色表示脯氨酸富含域(N端可變區(qū));灰色表示多元結(jié)構(gòu)域;黃色表示GTP酶結(jié)構(gòu)域;橙色表示含septin特征元件域;藍(lán)色表示卷曲螺旋(C端可變區(qū))。
圖1septin結(jié)構(gòu)域示意圖
2 septin的生理功能
2.1參與胞質(zhì)分裂septin基因最早發(fā)現(xiàn)于分裂缺陷型的釀酒酵母中,而酵母的胞質(zhì)分裂受阻往往也是由該基因的缺失或突變引起的。在酵母中,septin能定位于母細(xì)胞分裂的芽頸連接處,并且septin已被證明是參與募集許多細(xì)胞質(zhì)分裂相關(guān)蛋白必不可少的招募者[6]。此外,septin環(huán)的組裝過程并非是為細(xì)胞質(zhì)分裂提供所需的能量,而是發(fā)揮必要的調(diào)控作用。septin支架在酵母中的組裝過程需要微管和紡錘極體的定位,并且隨著后續(xù)的分離,經(jīng)過復(fù)制的染色體分別進(jìn)入到母細(xì)胞和子細(xì)胞中。哺乳動(dòng)物的septin與酵母類似,在完成胞質(zhì)分裂的過程中都起著重要的作用。研究表明,通過顯微注射親和純化的SEPT9抗體,再利用小干擾RNA(small interfering RNA,siRNA)使細(xì)胞質(zhì)分裂在早期和晚期階段出現(xiàn)缺陷,最終會(huì)導(dǎo)致雙核細(xì)胞的形成或者子細(xì)胞分離失敗,而SEPT2、SEPT7和SEPT11缺失也與SEPT9的研究結(jié)果一致[7]。要成功實(shí)現(xiàn)細(xì)胞分裂,最重要的就是細(xì)胞質(zhì)分裂和染色體分離之間要建立協(xié)調(diào)的關(guān)系。Spiliotis等[8]分別利用SEPT2和SEPT6對(duì)應(yīng)的抗體進(jìn)行熒光定位,發(fā)現(xiàn)這兩種蛋白質(zhì)都能定位在緊密附著有著絲點(diǎn)的紡錘體微管上。當(dāng)SEPT2和SEPT6通過siRNA干涉后,則會(huì)導(dǎo)致赤道板的染色體丟失、染色體不分離及主軸伸長(zhǎng)。
2.2參與建立細(xì)胞的擴(kuò)散阻礙作用除了在酵母母細(xì)胞芽頸連接處起支架作用外,septin還可通過芽頸限制細(xì)胞表皮因子的交換以起到擴(kuò)散阻礙的作用。和酵母類似,哺乳動(dòng)物細(xì)胞也能在細(xì)胞質(zhì)分裂的地方形成septin環(huán)狀結(jié)構(gòu)。然而,他們的擴(kuò)散阻礙作用并未像酵母一樣得到廣泛的研究。此外,皮層障礙在哺乳動(dòng)物卵裂溝內(nèi)的擴(kuò)散是目前中體(mid-body)已知的方式。septin的擴(kuò)散阻礙作用是由纖毛在septin形成環(huán)狀結(jié)構(gòu)的基礎(chǔ)上建立的[9]。初級(jí)纖毛是脊椎動(dòng)物絕大多數(shù)細(xì)胞頂面的一個(gè)細(xì)胞突起,其功能是作為一種感受器,并在細(xì)胞發(fā)育和組織穩(wěn)態(tài)的信號(hào)通路上起到關(guān)鍵的協(xié)調(diào)作用。人類疾病和已知的纖毛發(fā)育障礙主要是由細(xì)胞器的結(jié)構(gòu)和功能缺陷引起的。在纖毛形成期間纖毛會(huì)利用質(zhì)膜進(jìn)行膜融合,并能把相關(guān)感知和轉(zhuǎn)導(dǎo)多種胞外信號(hào)的獨(dú)特組蛋白保留下來。此外,一種內(nèi)含septin的擴(kuò)散隔膜就是在纖毛膜的基礎(chǔ)上形成的,并且已證明初生纖毛在形成過程中需要保留受體的信號(hào)通路[10]。這種擴(kuò)散阻礙限制了纖毛和外周纖毛膜之間的纖毛膜蛋白的擴(kuò)散,也為septin從真菌到哺乳動(dòng)物的進(jìn)化保守角色提供了一個(gè)經(jīng)典的案例。
2.3參與細(xì)胞極性的形成細(xì)胞極性是生物體和組織發(fā)育及實(shí)現(xiàn)生理功能所必需的基本過程。與酵母同源的哺乳動(dòng)物septin也需要建立和保持細(xì)胞極性,而細(xì)胞極性分化的共同特征就是存在一個(gè)不對(duì)稱的質(zhì)膜組織。在細(xì)胞水平上,建立和保持細(xì)胞極性在不同的生物體中都要遵循共同的過程。與酵母類似,建立哺乳動(dòng)物的細(xì)胞極性可以利用高度保守的Ras和Rho家族[11]。細(xì)胞極性的建立需要囊泡膜沿著供體與膜受體之間連接形成的細(xì)胞骨架軌道進(jìn)行運(yùn)輸。酵母已經(jīng)被證明是一個(gè)研究細(xì)胞極性建立和調(diào)控機(jī)制的很好模型。同時(shí),septin在哺乳動(dòng)物細(xì)胞極性中扮演的角色也初露端倪。Bowen等[12]在體外進(jìn)行犬腎細(xì)胞的極化研究時(shí)發(fā)現(xiàn),SEPT2能指導(dǎo)微管生長(zhǎng)和捕獲的方向,該過程對(duì)于保持微管的正確方向是至關(guān)重要的。先前的研究發(fā)現(xiàn)septin通過與微管蛋白MAP4(microtubule-associated protei)、微管乙酰化作用及微管聚谷酰胺的結(jié)合,從而控制微管的穩(wěn)定[13]。
3 Septin與人類疾病的關(guān)系
3.1Septin與腫瘤越來越多的研究證實(shí),septin基因家族直接參與了腫瘤的發(fā)生、發(fā)展。Septin與腫瘤發(fā)生有關(guān)的報(bào)道首次出現(xiàn)在惡性血液病中。研究表明,在患有急性髓細(xì)胞性白血病(acute myeloid leukemia,AML)的同卵雙胞胎患者體內(nèi),人們發(fā)現(xiàn)參與人類細(xì)胞分裂的周期相關(guān)基因hCDCrel/SEPT9如果出現(xiàn)染色體易位,就很容易和MLL(myeloid/lymphoid ormixed-lineage leukemia)基因形成融合基因或者伴侶基因[14]。后來,人們陸續(xù)發(fā)現(xiàn)SEPT2、SEPT5、SEPT6及SEPT11 4種septin基因都可以與MLL形成類似的融合基因[7,13]。這種融合現(xiàn)象多見于嬰兒型急、慢性粒細(xì)胞性白血病。此外,SEPT9的過量表達(dá)也與某些癌癥的發(fā)生有關(guān)。如人類乳腺癌的發(fā)生跟SEPT9基因產(chǎn)生的一種剪接本SEPT9_v1有直接的關(guān)系,在乳腺癌細(xì)胞及其組織中均能檢測(cè)到該剪接本上調(diào)表達(dá)。而恰恰乳腺癌細(xì)胞的增殖、侵襲及惡性轉(zhuǎn)化的速率與該剪接本過量表達(dá)有密切的關(guān)系。另外,SEPT9 _v1還可穩(wěn)定氨基末端激酶JNK(c-Jun N-terminal kinase)的表達(dá)水平,防止JNK降解,從而促進(jìn)JNK/c-Jun通路的活化,使下游的細(xì)胞周期蛋白CyclinD1表達(dá)上調(diào),促進(jìn)腫瘤細(xì)胞的增殖[15]。
除研究較為深入的SEPT9基因外,其他septin家族成員也與人類腫瘤的發(fā)生有密切的關(guān)系。如人類SEPT1的過量表達(dá)會(huì)使皮膚鱗狀細(xì)胞癌發(fā)生概率增加[16];最新的研究發(fā)現(xiàn),SEPT2的下調(diào)表達(dá)能使過氧化物酶體增殖物激活受體-γ(peroxisome proliferator-activated receptor-γ,PPARγ)活化,從而抑制肝癌細(xì)胞的生長(zhǎng)[17];SEPT3 的表達(dá)則與髓母細(xì)胞瘤等神經(jīng)元來源的腫瘤有關(guān)[18];SEPT4表達(dá)有組織特異性,并且與結(jié)腸癌、前列腺癌、腎細(xì)胞癌和膀胱癌的發(fā)生有關(guān)。此外,人們還發(fā)現(xiàn)SEPT4的兩個(gè)變異剪接本表達(dá)下調(diào)會(huì)抑制結(jié)腸癌細(xì)胞的增殖及惡性轉(zhuǎn)化[19]。
3.2Septin與感染性疾病人們?cè)谘芯克拗髋c微生物的相互作用時(shí),驚奇地發(fā)現(xiàn)septin在細(xì)菌感染和先天免疫中也扮演著很重要的角色。Septin首次與吞噬作用的過程聯(lián)系起來主要是根據(jù)他們已知的調(diào)控膜運(yùn)輸?shù)墓δ堋追Nseptin蛋白都能在小鼠的巨噬細(xì)胞中表達(dá),并且SEPT2和SEPT11都能定位在吞噬體上[20]。此后不久,隨著研究的深入,septin開始出現(xiàn)新的功能,其在保護(hù)宿主免受病原體的感染上起到關(guān)鍵的作用。這項(xiàng)研究主要是圍繞志賀氏桿菌(Shigella)開展,研究發(fā)現(xiàn)septin能聚集組裝成一個(gè)籠狀結(jié)構(gòu),該結(jié)構(gòu)能防御細(xì)菌在細(xì)胞間進(jìn)行擴(kuò)散[21]。膜動(dòng)力學(xué)的調(diào)控對(duì)于T淋巴細(xì)胞的遷移也起到至關(guān)重要的作用。利用小發(fā)卡RNA(small hairpin RNA,shRNA)消除septin的細(xì)胞骨架,會(huì)導(dǎo)致T細(xì)胞的形態(tài)結(jié)構(gòu)發(fā)生變化,此時(shí)T細(xì)胞會(huì)出現(xiàn)明顯的皺縮及多余的尖端突起[22]。此外,其他病原體如單增李斯特菌(Listeriamonocytogenes)和立克次氏體(Rickettsia)在入侵宿主細(xì)胞時(shí)同樣會(huì)受到septin蛋白形成的類似籠狀結(jié)構(gòu)的組織防御[9,23],這種結(jié)構(gòu)可以將病原體包裹在內(nèi)部,從而有利于體內(nèi)的溶菌酶對(duì)其進(jìn)行降解清除。
3.3Septin與神經(jīng)系統(tǒng)疾病研究表明,septin基因家族部分成員的功能失調(diào)與阿爾茨海默病、帕金森病及遺傳性神經(jīng)痛性肌萎縮等神經(jīng)系統(tǒng)疾病的發(fā)生和發(fā)展有著直接的關(guān)系。Macedo等[24]基于分子流行病學(xué)的研究發(fā)現(xiàn),SEPT3第11號(hào)外顯子的一個(gè)多態(tài)性位點(diǎn)與阿爾茨海默病發(fā)生有關(guān);Walden等[25]研究發(fā)現(xiàn),SEPT5的其中一個(gè)剪接本SEPT5_v2作為一種結(jié)合蛋白可與SEPT5另一個(gè)變異體v1結(jié)合,并在常染色體隱性青少年帕金森病腦組織中積聚。此外,遺傳性神經(jīng)痛性肌萎縮(Hereditary neuralgic amyotrophy,HNA)作為一種罕見的常染色體顯性遺傳性疾病,目前已被證實(shí)該病的發(fā)生與SEPT9的突變有直接的關(guān)系[26]。此外,患有帕金森病、癡呆癥及多發(fā)系統(tǒng)萎縮等疾病的患者中樞神經(jīng)系統(tǒng)均可看見大量的萊維小體(Lewy body),該小體已被確認(rèn)為此類神經(jīng)系統(tǒng)疾病患者腦內(nèi)的特征性標(biāo)志物,值得注意的是,SEPT4參與了該小體的形成[27]。
3.4Septin與男性不育癥Septin家族可能與男性不育癥有關(guān)。在已發(fā)現(xiàn)的13個(gè)人類septin成員中,SEPT1、SEPT4、SEPT6、SEPT7及SEPT12均被證明在精子細(xì)胞的分化、成熟及精子的運(yùn)動(dòng)功能中發(fā)揮著重要的作用。其中,SEPT4能定位于精子的環(huán)結(jié)構(gòu)。在精子發(fā)生過程中,SEPT4能促進(jìn)精子環(huán)結(jié)構(gòu)的形成。研究表明,把雄性小鼠SEPT4基因敲除后,會(huì)造成小鼠精子環(huán)結(jié)構(gòu)形成缺陷,進(jìn)而使鞭毛形態(tài)及運(yùn)動(dòng)功能受到影響,最終導(dǎo)致不育癥。2010年Chao等[28]報(bào)道1例畸形精子無力癥患者,其精子的鞭毛環(huán)結(jié)構(gòu)缺失率達(dá)97%,進(jìn)一步的定位結(jié)果顯示,SEPT7信號(hào)在該患者的精子鞭毛環(huán)結(jié)構(gòu)處較弱,因此臨床上也把SEPT7作為檢測(cè)精子成熟的生物標(biāo)記。除此之外,SEPT6和SEPT12的表達(dá)缺陷也會(huì)造成精子環(huán)結(jié)構(gòu)的紊亂,出現(xiàn)精子無力現(xiàn)象,導(dǎo)致不育[29]。
4 展 望
Septin基因首次在酵母中被發(fā)現(xiàn)距今已過去40多年的時(shí)間,在哺乳動(dòng)物細(xì)胞中的研究也過去了有20年的時(shí)間,在這期間,人們對(duì)septin進(jìn)行了大量而深入的研究工作,也取得了很多研究成果,尤其是作為支架蛋白的septin絲狀結(jié)構(gòu)為深入研究septin的功能提供了大量的證據(jù)。目前的研究表明,septin參與了很多重要的生理過程,其中包括細(xì)胞分裂、細(xì)胞極性形成、細(xì)胞的擴(kuò)散阻礙、囊泡運(yùn)輸、胞吞胞吐和細(xì)胞凋亡等。在這些過程中septin可能扮演了一種腳手架蛋白的角色,其功能可能是招募或阻隔相關(guān)的細(xì)胞功能蛋白來參與完成上述的一系列生理過程,但由于septin基因家族結(jié)構(gòu)的特殊性和成員的復(fù)雜性,其詳細(xì)的生理功能還有待進(jìn)一步深入研究。此外,septin的異常表達(dá)與人類疾病的發(fā)生、發(fā)展有直接的關(guān)系。這些人類疾病包括腫瘤發(fā)生、神經(jīng)功能障礙和病原微生物感染及男性不育癥等。隨著后續(xù)研究的深入,septin作為一種蛋白載體發(fā)揮功能的方式也逐漸得到了眾多研究者的認(rèn)同,這些septin蛋白要么加強(qiáng)功能蛋白間的相互作用,要么阻隔二者間的互作,通過這兩種作用方式來調(diào)節(jié)細(xì)胞信號(hào)通路的活化或抑制。正是由于它們的特殊性和重要性,septin基因家族與人類疾病的關(guān)系才越來越受到人們的關(guān)注,并取得了很多重要進(jìn)展,這也為今后的研究奠定了基礎(chǔ),未來的研究重點(diǎn)可能會(huì)更多的集中到septin蛋白在細(xì)胞吞噬中所發(fā)揮的作用上,此外,繼續(xù)對(duì)septin進(jìn)行基因水平上的探索也是今后研究的趨勢(shì),而這些septin功能的研究和闡釋必將會(huì)對(duì)人類疾病的診斷和治療產(chǎn)生重要的意義。
[1]Weirich CS,Erzberger JP,Barral Y.The septin family of GTPases:architecture and dynamics[J].Nat Rev Mol Cell Biol,2008,9(6):478-489.
[2]Russell SE,Hall PA.Septin genomics:a road less travelled[J].Biol Chem,2011,392(8/9):763-767.
[3]Zent E,Wittinghofer A.Human septin isoforms and the GDP-GTP cycle[J].Biol Chem,2014,395(2):169-180.
[4]Sirajuddin M,Farkasovsky M,Zent E,et al.GTP-induced conformational changes in septins and implications for function[J].Proc Natl Acad Sci U S A,2009,106(39):16592-16597.
[5]Montagna C,Bejerano-Sagie M,Zechmeister JR,et al.Mammalian septins in health and disease[J].Int J Nano Med,2014,9:1883-1889.
[6]Hanrahan J,Snyder M.Cytoskeletal activation of a checkpoint kinase[J].Mol Cell,2003,12:663-673.
[7]Estey MP,Di Ciano-Oliveira C,Froese CD,et al.Distinct roles of septins in cytokinesis:SEPT9 mediates midbody abscission[J].J Cell Biol,2010,191(4):741-749.
[8]Spiliotis ET,Kinoshita M,Nelson WJ.A mitotic septin scaffold required for mammalian chromosome congression and segregation[J].Science,2005,307(5716):1781-1785.
[9]Saarikangas J,Barral Y.The emerging functions of septins in metazoans[J].EMBO Rep,2011,12(11):1118-1126.
[10]Hu Q,Milenkovic L,Jin H,et al.A septin diffusion barrier at the base of the primary cilium maintains ciliary membrane protein distribution[J].Science,2010,329(5990):436-439.
[11]McCaffrey LM,Macara IG.Widely conserved signaling pathways in the establishment of cell polarity[J].Cold Spring Harb Perspect Biol,2009,1(2):a001370.
[12]Bowen JR,Hwang D,Bai X,et al.Spiliotis ET.Septin GTPases spatially guide microtubule organization and plus end dynamics in polarizing epithelia[J].J Cell Biol,2011,194(2):187-197.
[13]Spiliotis ET.Regulation of microtubule organization and functions by septin GTPases[J].Cytoskeleton,2010,67(6):339-345.
[14]Santos J,Cerveira N,Correia C,et al.Coexistence of alternative MLL-SEPT9 fusion transcripts in an acute myeloid leukemia with t(11;17)(q23;q25)[J].Cancer Genet Cytogenet,2010,197(1):60-64.
[15]Gonzalez ME,Makarova O,Peterson EA,et al.Up-regulation of SEPT9_v1 stabilizes c-Jun-N-terminal kinase and contributes to its pro-proliferative activity in mammary epithelial cells[J].Cell Signal,2009,21(4):477-487.
[16]Mizutani Y,Ito H,Iwamoto I,et al.Possible role of a septin,SEPT1,in spreading in squamous cell carcinoma DJM-1 cells[J].Biol Chem,2013,394(2):281-290.
[17]Cao LQ,Shao ZL,Liang HH,et al.Activation of peroxisome proliferator-activated receptor-γ (PPARγ) inhibits hepatoma cell growth via downregulation of SEPT2 expression[J].Cancer Lett,2015,359(1):127-135.
[18]Sellin ME,Stenmark S,Gullberg M.Cell type-specific expression of SEPT3-homology subgroup members controls the subunit number of heteromeric septin complexes[J].Mol Biol Cell,2014,25(10):1594-1607.
[19]Fuchs Y,Brown S,Gorenc T,et al.Sept4/ARTS regulates stem cell apoptosis and skin regeneration[J].Science,2013,341(6143):286-289.
[20]Huang YW,Yan M,Collins RF,et al.Mammalian septins are required for phagosome formation[J].Mol Biol Cell,2008,19:1717-1726.
[21]Mostowy S,Bonazzi M,Hamon MA,et al.Entrapment of intracytosolic bacteria by septin cage-like structures[J].Cell Host Microbe,2010,8(5):433-444.
[22]Tooley AJ,Gilden J,Jacobelli J,et al.Amoeboid T lymphocytes require the septin cytoskeleton for cortical integrity and persistent motility[J].Nat Cell Biol,2009,11(1):17-26.
[23]Reed SC,Lamason RL,Risca VI,et al.Rickettsia actin-based motility occurs in distinct phases mediated by different actin nucleators[J].Curr Biol,2014,24(1):98-103.
[24]Macedo JN,Valadares NF,Marques IA,et al.The structure and properties of septin 3:a possible missing Link in septin filament formation[J].Biochem J,2013,450(1):95-105.
[25]Walden H,Martinez-Torres RJ.Regulation of parkin E3 ubiquitin ligase activity[J].Cell Mol Life Sci,2012,69(18):3053-3067.
[26]Leshinsky-Silver E,Ginzberg M,Dabby R,et al.Neonatal vocal cord paralysis-an early presentation of hereditary neuralgic amyotrophy due to a mutation in the SEPT9 gene[J].Eur J Paediatr Neurol,2013,17(1):64-67.
[27]Ageta-Ishihara N,Yamakado H,Morita T,et al.Chronic overload of SEPT4,a parkin substrate that aggregates in Parkinson′s disease,causes behavioral alterations but not neurodegeneration in mice[J].Mol Brain,2013,6(1):35.
[28]Chao HC,Lin YH,Kuo YC,et al.The expression pattern of SEPT7 correlates with sperm morphology[J].J Assist Reprod Genet,2010,27(6):299-307.
[29]Kuo YC,Lin YH,Chen HI,et al.SEPT12 mutations cause male infertility with defective sperm annulus[J].Hum Mutat,2012,33(4):710-719.
10.3969/j.issn.1671-8348.2016.25.039
重慶市基礎(chǔ)與前沿研究計(jì)劃項(xiàng)目(cstc2014jcyjA80019);重慶三峽醫(yī)專自然科學(xué)重點(diǎn)項(xiàng)目(2016xzz03)。
馬強(qiáng)(1986-),講師,碩士,主要從事生物化學(xué)與分子生物學(xué)的研究。
,E-mail:hpo1979@126.com。
R363
A
1671-8348(2016)25-3570-04
2016-03-18
2016-05-26)
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
大科技·百科新說(2021年6期)2021-09-12 02:37:27
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
好孩子畫報(bào)(2020年5期)2020-06-27 14:08:05
科技傳播(2019年22期)2020-01-14 03:06:54
意林·全彩Color(2019年6期)2019-07-24 08:13:50
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
中國(guó)科技論壇(2017年7期)2017-07-25 08:49:53
奧秘(2015年2期)2015-09-10 07:22:44

- 重慶醫(yī)學(xué)的其它文章
- 持續(xù)質(zhì)量改進(jìn)與醫(yī)院感染率的分析
- 應(yīng)用PDCA循環(huán)管理提高肝膽外科住院患者滿意度的研究
- 系統(tǒng)化健康教育對(duì)經(jīng)皮肺穿檢查患者干預(yù)效果的分析
- 反思日記在準(zhǔn)實(shí)習(xí)護(hù)生外科綜合實(shí)訓(xùn)教學(xué)中的效果分析
- 《急救護(hù)理》網(wǎng)絡(luò)資源教學(xué)平臺(tái)用戶評(píng)價(jià)指標(biāo)體系的構(gòu)建與實(shí)踐研究*
- 口腔實(shí)習(xí)生醫(yī)患溝通技巧的情景教學(xué)探索*