降低日糧蛋白質含量并補充氨基酸對泌乳母豬生產性能、乳腺吸收效率和轉運體基因表達的影響
2016-11-08 04:08:20ManjarinZamoraWuSteibelKirkwoodTaylorWilsPlotzTrifiloTrottier
中國飼料 2016年15期
關鍵詞:效率
Manjarin R,Zamora V,Wu G,Steibel J P,4,Kirkwood R N,Taylor N P,Wils-Plotz E,Trifilo K,Trottier N L
(1.密歇根州立大學動物科學學院,東蘭辛48824;2.墨西哥州Montecillo研究生院,墨西哥56230;3.德克薩斯州農工大學動物科學和營養學部,學院站77843;4.密歇根州立大學漁業和野生動物系,東蘭辛48824;5.美國密歇根州立大學大型動物臨床科學系,東蘭辛48824)
中國·豬營養國際論壇
降低日糧蛋白質含量并補充氨基酸對泌乳母豬生產性能、乳腺吸收效率和轉運體基因表達的影響
Manjarin R1,Zamora V2,Wu G3,Steibel J P1,4,Kirkwood R N5,Taylor N P1,Wils-Plotz E1,Trifilo K1,Trottier N L1
(1.密歇根州立大學動物科學學院,東蘭辛48824;2.墨西哥州Montecillo研究生院,墨西哥56230;3.德克薩斯州農工大學動物科學和營養學部,學院站77843;4.密歇根州立大學漁業和野生動物系,東蘭辛48824;5.美國密歇根州立大學大型動物臨床科學系,東蘭辛48824)
中國豬營養國際論壇是由美國動物科學學會、上海亙泰實業集團和上海優久生物科技有限公司聯合主辦,以“凝聚全球科研力量,驅動豬業創新思維”為宗旨,力邀全球一流的機構、專家和學者,傾力打造一個動物營養領域具有國際性、前沿性和權威性的論壇。該論壇每兩年舉辦一屆,聚焦行業發展中的熱點、難點,通過專家學者和企業領導者之間進行開放建設性的學術探討、理論研究和實踐經驗交流,整合全球動物營養領域最新的技術和研究成果,推動行業發展,創造和提升產業價值。
www.asaschina.org
為驗證降低日糧蛋白質水平并補充晶體氨基酸是否可以提高氨基酸用于泌乳母豬產奶的使用效率,試驗選用3種日糧,蛋白質含量分別為9.5%(不足型)、13.5%(理想型)和17.5%(標準型)飼喂泌乳母豬,各日糧氨基酸水平相同。檢測乳腺氨基酸動靜脈濃度差(A-V)、氨基酸運輸效率(A-V/A×100)、氨基酸轉運載體和乳蛋白基因的轉錄豐度。在第7和18天,收集采食后動脈、乳腺靜脈血液和乳腺組織樣品。使用定量PCR測定氨基酸轉運載體b0,+AT(SLC7A9)、Y+LAT2(SLC7A6)、ATB0,+(SLC6A14)、CAT-1(SLC7A1)、CAT-2b(SLC7A2)以及β酪蛋白(CSN2)和α乳清蛋白(LALBA)的轉錄豐度。結果表明,隨著蛋白質含量從不足型到標準型逐漸增加,仔豬日增重呈曲線上升(線性和二次方,P<0.03)。第7天,隨粗蛋白質含量增加,賴氨酸和精氨酸A-V、運輸效率呈二次方增加(P<0.05)。第18天,隨粗蛋白質含量增加,賴氨酸A-V呈增加趨勢(線性,P=0.08)。第7天,隨著粗蛋白質含量增加,異亮氨酸和纈氨酸A-V增加(呈線性,分別為P=0.05和P=0.08)。第18天,異亮氨酸和纈氨酸濃度增加(呈線性,分別為P=0.04和P=0.07)。第7天,血液支鏈AA(BCAA):賴氨酸(Lys)比值呈二次方降低(P<0.05)。SLC7A9、SLC7A6、SLC6A14、SLC7A1、SLC7A2、CSN2和LALBA的基因表達不受日糧影響。總之,日糧蛋白質含量從17.5%下降到13.5%,并補充晶體氨基酸并不影響仔豬日增重,氨基酸轉運載體或乳蛋白基因的表達,但可以增加泌乳第7天乳腺運輸效率和賴氨酸與精氨酸動靜脈濃度差。這種增加與血液BCAA∶Lys比值下降有關,表明氨基酸運輸至乳腺細胞時,陽離子型AA和BCAA之間存在競爭機制。
氨基酸;效率;基因;哺乳期;母豬;轉運載體
為了使豬更好地利用日糧蛋白質,越來越多的研究關注最佳日糧氨基酸(AA)水平(NRC,2012)。有研究表明,日糧AA過量或不足引起的AA不平衡可導致哺乳母豬氮利用率降低,乳蛋白合成和窩增重受限(Pérez-Laspiur等,2009;Guan等,2004)。對于生長豬,減少日糧粗蛋白質(CP)并補充晶體AA,可以增加賴氨酸利用效率,并降低N排泄量(Otto等,2003)。然而,關于氨基酸利用率在細胞水平上的作用機制還不是很清楚。日糧AA胞內可利用性是由位于細胞膜上允許氨基酸進入細胞內的蛋白質載體控制(Bro?r,2008;Palacín等,1998;Shennan等,1997)。
賴氨酸(Lys)是哺乳母豬第一限制性氨基酸(NRC,1998),因此涉及賴氨酸攝取的轉運蛋白可能對整個乳腺日糧蛋白質利用率發揮關鍵作用。曾報道為了滿足奶需求(Manjarín等,2011)和日糧氨基酸可利用性(Pérez-Laspiur等,2009),使編碼Lys轉運載體的基因轉錄豐度增加。因此,由乳腺動脈循環提取AA的比例反映了AA轉運速率相對于動脈AA可用度。本試驗中,AA轉運效率被定義為AA提取率。
我們推測,豬乳腺轉運AA效率增加,與日糧CP減少并補充晶體AA相關,這種增加是通過編碼Lys轉運載體的乳腺基因mRNA豐度增加來調控。本試驗目的:(1)檢測減少日糧蛋白質,并補充晶體AA,是否可以通過乳腺奶產量來使Lys利用效率增加;(2)對編碼特定乳腺Lys轉運蛋白和主導乳腺合成乳蛋白的基因表達進行定量分析。
1 材料與方法
本研究中所有動物試驗程序由密歇根州立大學動物護理學院和使用委員會批準(AUF號為10 /08-162-00)。
1.1動物和組織收集試驗選用24頭經產母豬(長×大),3個重復(組)。每組被定義為一個產仔周期,第一組和第二組包含9頭母豬,第三組包含6頭母豬。試驗中3頭母豬由于采食量過低或導管相關問題被踢除;因此,第一組和第二組分別包含8頭和7頭母豬。整個研究過程中所有母豬單獨飼養在20℃分娩欄中。預產期前一周,母豬被隨機分配給三種日糧:9.5%的CP(不足型;N=8),13.5%的CP(理想型;N=6)和17.5%的CP(標準型;N=7)。分娩前所有母豬每天兩次飼喂,共2.5 kg日糧(飼喂基礎)。分娩后的第二天當作哺乳期第1天,分別在8∶00和16∶00飼喂母豬1 kg飼料。哺乳期第2和3天,分別飼喂母豬3、4 kg。其他時間,每天最多飼喂5.5 kg,以確保所有母豬干物質采食量均等。整個哺乳期每天記錄母豬采食量。自由飲水。泌乳期第2和18天記錄母豬體重。總重約15 kg的仔豬在出生后24 h內完成交叉寄養,每窩均分8頭仔豬。哺乳期第2和18天記錄窩重,第21天斷奶。
1.2日糧處理和飼料營養分析不足型、理想型和標準型日糧中分別含有0.50%、0.81%、1.01%的標準回腸可消化(SID)Lys(表1)。通過向標準日糧中添加淀粉和蔗糖降低CP含量,以得到理想型和不足型日糧,但整個日糧保持豆粕玉米比例不變。理想型日糧中添加晶體AA以滿足哺乳10頭仔豬的母豬對必需氨基酸需求和AA∶Lys比,預測仔豬日增重為200 g/d,母豬采食量為4.61 kg/d,哺乳期體重變化為-10 kg(NRC,1998)。標準型日糧中添加晶體L-賴氨酸,使其比理想型高30%(即為1.01%和0.81%SID)。標準日糧中賴氨酸量按照行業標準選擇(安大略省,圭爾夫大學;美國農業部,農業科學研究院)。為了滿足NRC(1998)AA∶Lys比例,并維持與理想型日糧相同的AA配方,標準日糧中添加晶體AA。不足型日糧中添加L-賴氨酸以滿足0.50%的SID賴氨酸,從而實現不足型、理想型和標準型日糧之間SID賴氨酸相近。對于標準型日糧,添加晶體AA以確保與理想型日糧有相同的AA∶Lys比例。此外,日糧中添加大豆油以減少粉塵并改善適口性,Solka-Floc(紐約,國際纖維有限公司)用來平衡整個日糧纖維水平。
每組制備新鮮日糧,避免長期儲存3個不同批次的混合日糧。從每包取樣,充分混合。所有混合樣品用樣品研磨機(Cyclotec 1093;Foss Teca-tor,Eden Prairie,MN)磨細。使用燃燒測定儀(FP-2000,LECO Corp.,St.Joseph,MI)分析每個樣品的N含量,并取平均值。混合飼料樣品中氨基酸濃度用陽離子交換色譜分析,柱后茚三酮衍生和定量分析(農業試驗站,密蘇里大學,哥倫比亞大學,MO)。AA計算值和分析值見表2。
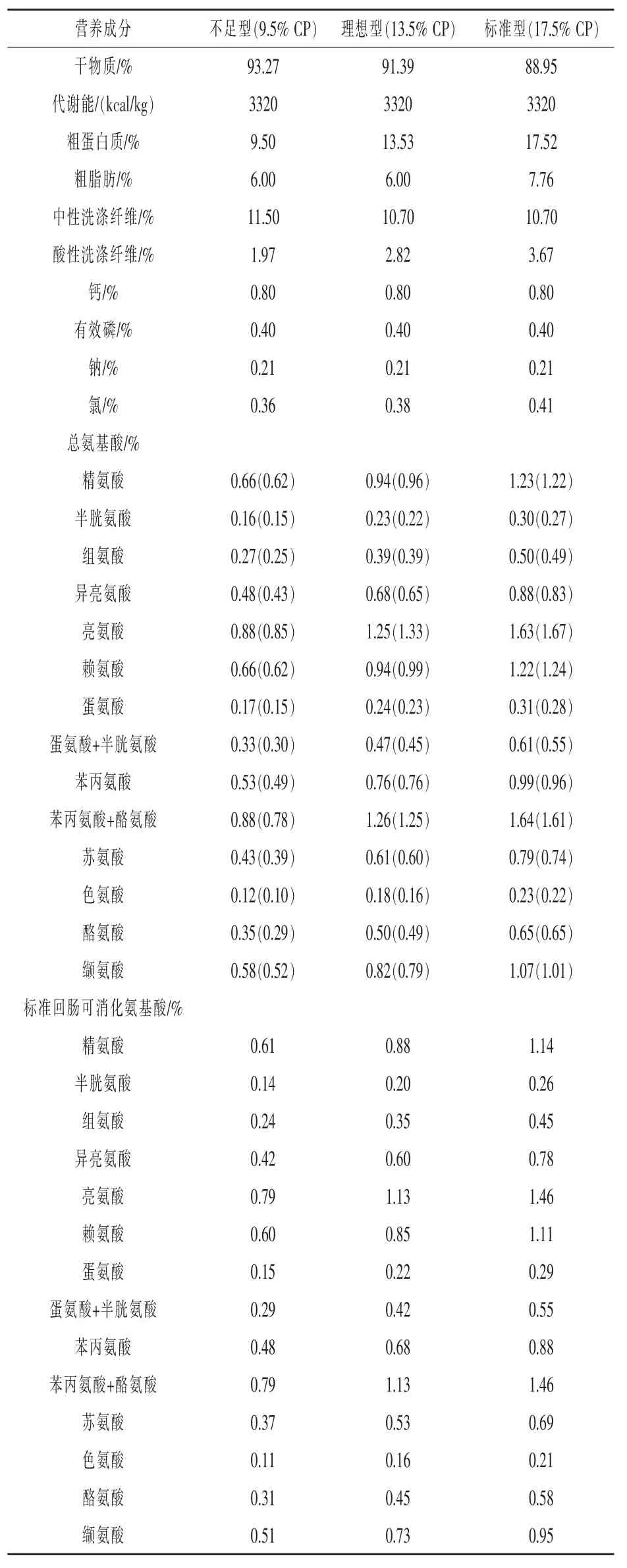
表2 試驗日糧能量和營養成分計算值和分析值(飼喂基礎)
1.3基因表達分析
1.3.1樣品采集哺乳期第7天(早期)和第18天(峰值)所有母豬采食后3.5 h,根據Kirkwood等(2007)描述的方法,從第一和第二胸腺對乳腺實質組織進行活體檢查。乳腺組織收集期間,仔豬被隔離在配備有保溫燈的相鄰欄。活檢后,將乳腺組織快速冷凍在液氮中,-80℃保存。活檢后3 h,仔豬被送回母豬欄,并允許哺乳。
1.3.2RNA提取和cDNA合成用試劑盒從乳腺組織提取核糖核酸(PerfectPure RNA細胞與組織試劑盒;PRIME,馬里蘭州蓋瑟斯堡)。對分離的RNA進行定性和定量測定(安捷倫科技帕洛阿爾托,加利福尼亞安捷倫生物分析儀2100與核糖核酸6000納米芯片實驗室)。所有樣品RNA完整性數值范圍為8.7~10。每個樣品使用2 μg總RNA作為反轉錄模板合成互補DNA(SuperscriptⅢ反轉錄酶和寡(dT)15-18引物;英杰公司,卡爾斯巴德,加利福尼亞州)。用分光光度法對最終cDNA濃度進行定量分析(NanoDrop 1000;Thermo Scientific,Waltham,MA),然后工作原液稀釋至10 ng/μL,并于-20℃儲存。
1.3.3引物設計參考基因和靶基因的引物序列分別見表3和表4。本試驗中,每個靶基因被稱為所編碼蛋白質的通用名稱。因此,SLC7A9將被稱為b0,+AT,SLC7A7稱為y+LAT1,SLC7A6稱為y+LAT2,SLC6A14稱為ATB0,+,SLC7A1稱為CAT-1,SLC7A2稱為CAT-2b,CSN2稱為β-酪蛋白,LALBA稱為α-乳蛋白。依據先前的研究選擇備選參考基因(Tramontana等,2008;Bionaz和Loor,2007),但本試驗所用的引物與已經發表的文章里反轉錄定量PCR(RT-qPCR)的優化效率不同。引物設計依據美國國家生物技術信息中心公開可用的豬cDNA和表達序列標簽序列(貝塞斯達,MD),使用缺省設置的軟件數據庫(引物設計軟件3.0版)。設計的引物與已發表的豬、人、牛和大鼠的基因組序列比對,核苷酸序列配對顯示出顯著對齊(即高的查詢覆蓋率),而其他物種所提到的蛋白比對被丟棄。本研究沒有對引物對擴增子進行序列測定。形成引物二聚體的判斷是基于RT-qPCR反應后解離曲線中單峰的存在。引物設計不是跨兩個外顯子結合。然而,提取RNA所用的方法為下一步柱上DNase處理的DNA裂解消除了基因組DNA(PerfectPure RNADNase,蓋瑟斯堡,馬里蘭州)。使用引物優化矩陣來優化引物對濃度(Mikeska和Dobrovic,2009),并用相對標準曲線來確定效率(Yuan等,2006)。用所有樣品的RNA庫合成cDNA建立標準曲線,cDNA含量為(一式兩份):40、20、10、5、2.5 ng。每個基因RT-qPCR反應效率由Yuan等(2006)描述的標準曲線斜率計算,公式為(10-1/斜率-1)×100。引物的特異性雜交通過RT-qPCR擴增末端解離曲線中特有峰的存在驗證。所有RT-qPCR模板包含非模板鏈,以驗證引物擴增不含污染的DNA。
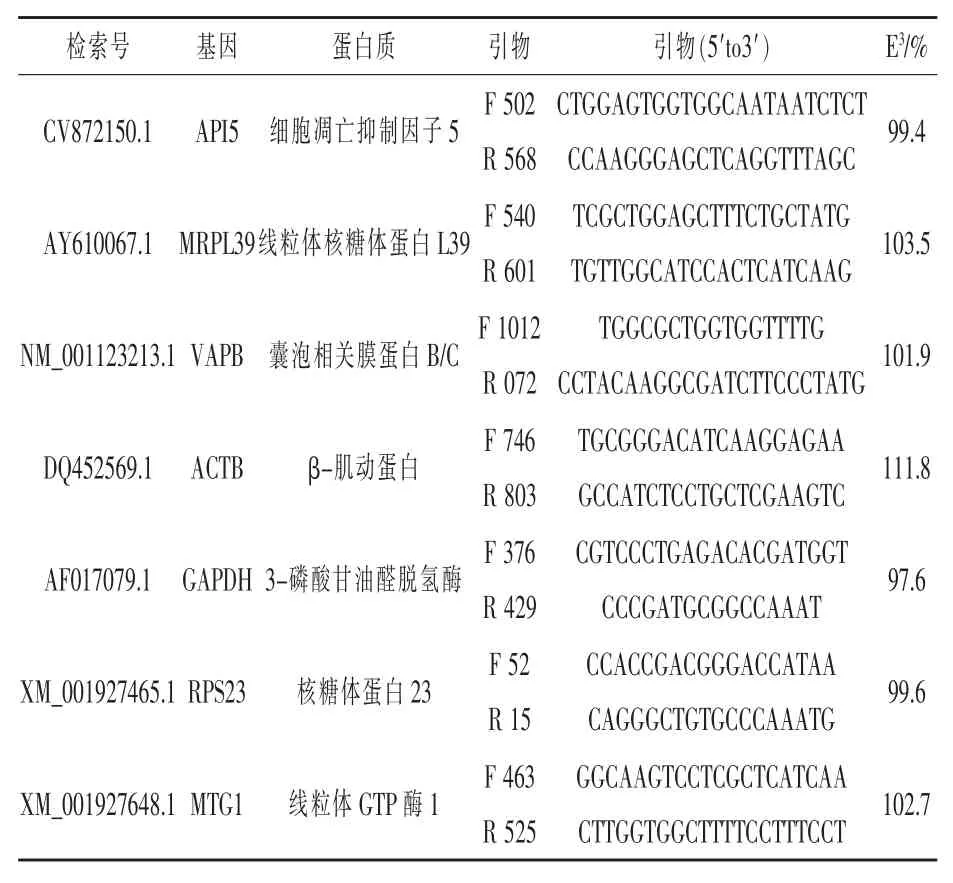
表3 反轉錄定量PCR(RT-qPCR)參考基因引物信息
1.3.4參考基因選擇使用相對標準曲線(Larionov等,2005)作為RT-qPCR方法來測定潛在參考基因的mRNA豐度。如Vandesompele等(2002)所描述,標準曲線中相對mRNA量直接被輸入到軟件(geNorm,http://medgen.ugent.be/~jvdesomp/genorm),選擇分析設置中最穩定的參考基因,簡單地說,每個基因的表達穩定值被確定為所有參考基因中每個基因的平均成對變異,而所用的參考基因數目由2個連續的歸一化因子之間的配對變異分析值計算。此類歸一化因子是基于n和n+1最佳參考基因表達的幾何平均值(Vandesompele等,2002)。
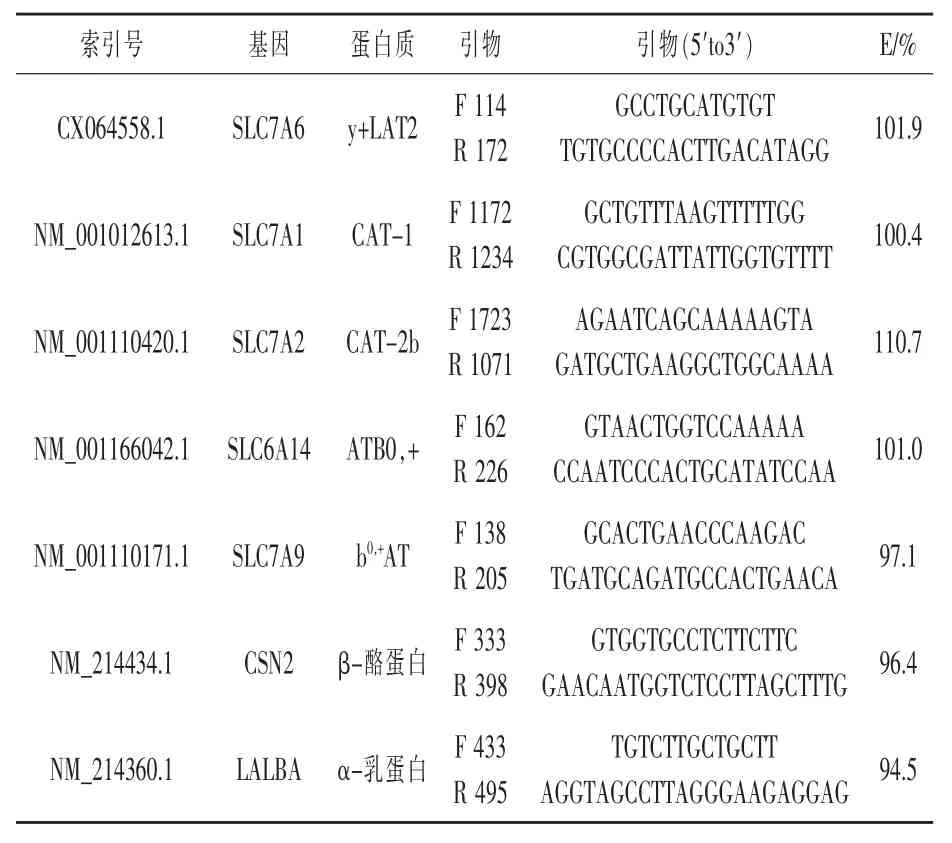
表4 反轉錄定量PCR(RT-qPCR)靶基因引物信息
1.3.5RT-qPCR分析反轉錄定量PCR反應使用美國應用生物系統公司MicroAmp光學96孔反應板進行。向每個孔中加入3 μL(30 ng)模板cDNA,12.5μL SYBR熒光標記物(Applied Biosystems公司),10 μmol/L正向和反向引物各6 μL,3.5 μL經焦碳酸二乙酯處理不含核苷酸的水(DEPC-水)(Fisher Scientific公司,新澤西州)。將板密封,于15℃、400 g離心1 min,置于系統中(ABI PRISM7000序列檢測系統,Applied Biosystems公司)。擴增程序包括2個初始步驟(50℃,2 min;95℃,10 min),接著進行40次循環(95℃,15 s;60℃,1 min)和解離曲線(95℃,15 s;60℃,1 min;95℃,15 s)。用7000 RQ序列檢測系統軟件2.2.1版本(應用生物系統公司)分析數據。
1.3.6RT-qPCR數據標準化靶基因表達值的標準化是按照以下公式計算:
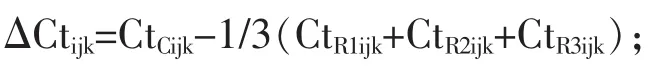
式中:ΔCtijk為第j頭母豬、哺乳第i階段、第k天日糧標準化目標基因的表達;CtCijk、CtRijk分別是目標基因和參考基因閾值循環。基因表達結果顯示ΔCt值往往令人困惑,因為ΔCt的增加反映了mRNA豐度降低。因此,在統計分析之前或之后從常量中減去ΔCt值,使得對數據的解釋更簡單,從而增加的ΔCt值與增加的mRNA豐度相對應,而不改變P值或SE值。為了進一步簡化對結果的解釋,這個常數選擇為大于所有基因中任何ΔCt值的最大數值。因此,在圖中所有的值都是正的。
1.3.7AA轉運至乳腺效率分析如Trottier等(1997)所述,每組日糧處理的5頭母豬在哺乳期第3天和第4天插入乳腺靜脈和頸動脈導管。每12 h用無菌肝素化(20 U/mL)的鹽溶液(0.9%)導管沖洗,以維持肝素環境。采樣前一晚飼喂母豬1 kg料,以保證第二天早上日糧被完全消耗。哺乳期第7天和第18天,所有母豬在8∶00喂2 kg飼料,從8∶30到11∶30每隔30 min采集血液樣品。用注射器同時采動脈和靜脈血(各8 mL),并轉移到樣品管(內含0.147 mL的75 g/L EDTA溶液;Becton Dickinson公司,富蘭克林湖,新澤西州)。前3 mL被丟棄,以消除被肝素稀釋的影響。血液樣品在600 g離心10 min,并將血漿于-20℃儲存。通過鄰苯二甲酸衍生后,使用HPLC檢測血漿中的游離AA(Li等,2011;Wu和Meininger,2008)。
將每天3.5 h收集的母豬血漿樣品進行混合,每個混合樣當作一個樣品單位。計算出第7和18天的AA轉運效率作為AA循環占整個乳腺的百分比:
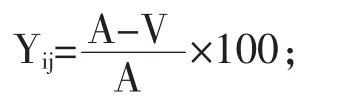
式中:Yij為j頭母豬哺乳第i階段AA運輸效率;A-V是動脈和靜脈AA濃度之差。乳腺總DNA定量。
根據Labarca和Paigen的方法(1980)使用冷高氯酸提取乳腺組織中總DNA(Sigma-Aldrich公司,圣路易斯,密蘇里州)。提取后,通過雙苯酰亞胺分光光度法(Hoechst 33258,Sigma-Aldrich公司)和酶標儀(Bio-Tek FL600,360/460 nm)對DNA進行定量分析,結果顯示為μg/mg乳腺組織。)。
1.4數據分析使用SAS統計軟件單變量程序下Shapiro-Wilk檢驗做正態分布分析。使用線性混合模型,將日糧、泌乳階段和互作效應作為固定因子,母豬和分組作為隨機因子,母豬初始體重和胎次作為協變量,分析仔豬ADG、母豬體重損失、窩增重、母豬飼料和蛋白采食量、乳腺組織DNA濃度、ΔCt、血漿AA水平、A-V和轉運效率以及血漿支鏈AA(BCAA)。該統計模型是:
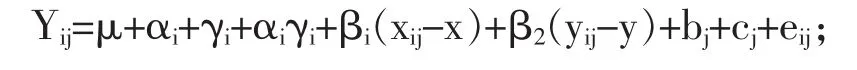
式中:Yij是哺乳階段(i)測得的變量(j),μ是總平均值,αi是i階段日糧的固定效應,γi是第i個泌乳階段的固定效應,αiγi是第i階段日糧和泌乳之間的交互作用,βi是與協變量母豬初始體重相關變量的回歸系數,xij表示為第j頭母豬在哺乳期第i階段的初始體重,x是所有母豬初始體重的平均數,β2是與協變量母豬肽次相關變量的回歸系數,yij是第j頭母豬第i泌乳階段的肽次,y是整期肽次平均值,bj是母豬的隨機效應,cj是欄的隨機效應,eij是試驗誤差。
使用線性和二次方對照分析日糧CP攝入、哺乳天數及其交互作用。交互作用不顯著時,以主效應的P值作為結果。數據表示為最小二乘方平均值±標準誤。P≤0.05表示差異顯著,P≤0.1表示有差異趨勢。
2 結果
由表5可見,日糧CP攝入不影響采食量,哺乳期第7天到第17天,飼料和日糧CP攝入均明顯升高(P<0.01)(結果未顯示)。不同日糧間第1天或第17天母豬體重和產仔數沒有明顯差異。仔豬日增重(線性,P=0.03;二次方,P=0.01)和窩增重(線性,P=0.07;二次方,P=0.01)隨著日糧CP水平增加而增加。日糧或哺乳階段對乳腺DNA濃度沒有影響。
參考基因MTG1、MRPL39和VAPB具有最小平均值(穩定值),因此被選擇為豬乳腺組織中最穩定的基因模型(圖1)。連續歸一化因子之間的成對變異分析低于0.15臨界值,參考基因的最佳數目是3。第四基因的加入增加了(V2/3=0.11,V3/ 4=0.12)成對變異(圖2)。編碼基因b0,+AT、y+LAT2、ATB0,+、CAT-1、CAT-2b、β酪蛋白、α乳蛋白的表達不受日糧影響(表6)。
動脈必需AA濃度、A-V濃度差、整個乳腺運輸效率分別見表7、表8和表9。動脈非必需AA濃度、A-V濃度差、整個乳腺運輸效率分別見表10、表11和表12。哺乳第7和8天,隨著CP濃度增加,動脈纈氨酸濃度呈線性增加(P<0.01),而亮氨酸呈曲線增加(線性,P=0.001;二次方,P<0.01)(表7)。第7天賴氨酸(線性,P=0.06)和異亮氨酸(線性和二次,P=0.08)有增加的趨勢,第18天賴氨酸呈線性增加(P=0.01),異亮氨酸呈曲線增加(線性,P=0.02;二次方,P=0.08)。哺乳第7天(P=0.09)和18天(P=0.07)精氨酸呈線性增加趨勢。
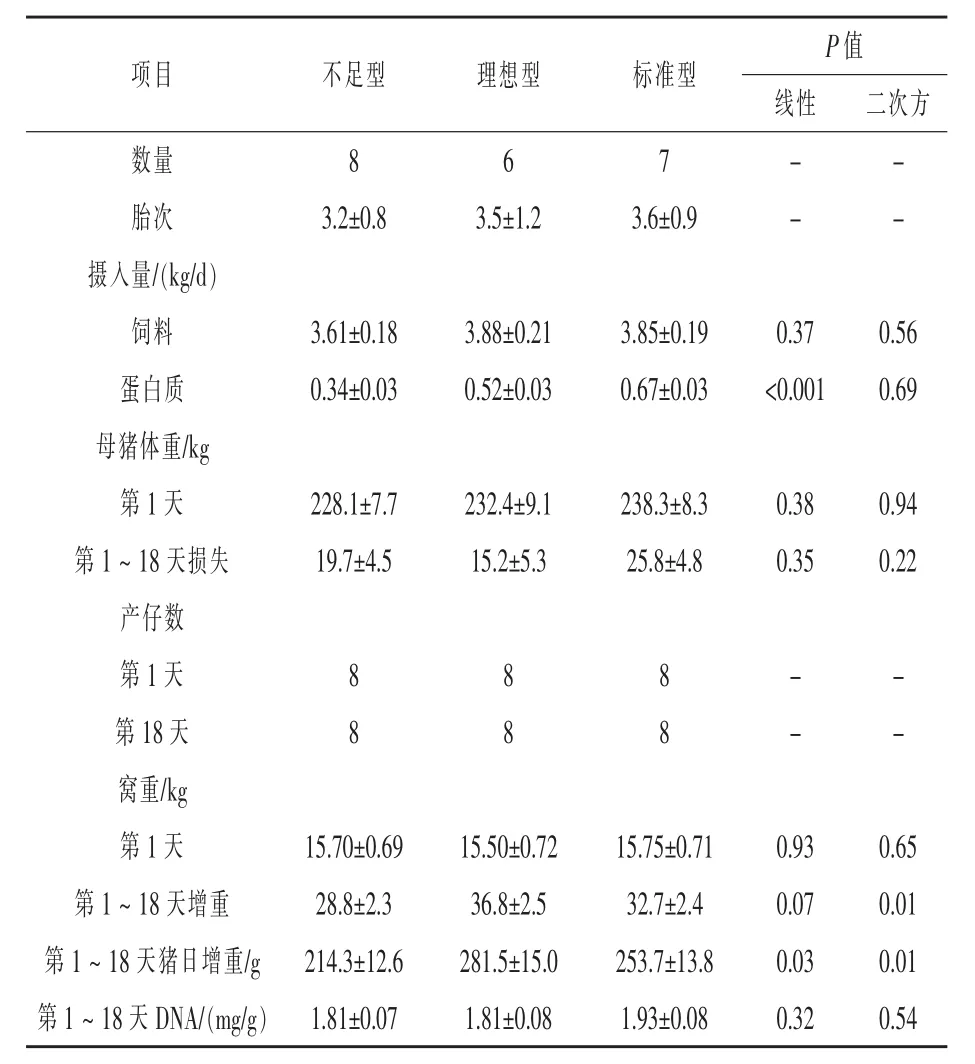
表5 日糧蛋白質水平對母豬、仔豬性能和乳腺組織DNA濃度的影響

圖1 參考基因API5、MRPL39、VAPB、MTG1、RPS21、ACTB和GAPDH的平均表達穩定性值

圖2 歸一化因子NFn和NFn+1之間的成對變異系數(Vn/Vn+1),來確定標準化參考基因的最佳數目
第7天,賴氨酸和精氨酸的動靜脈濃度差(P<0.03)和傳輸效率(P<0.05)明顯呈二次方增加,而隨著CP比例增加異亮氨酸動靜脈濃度差(P= 0.05)和運輸效率(P=0.08)呈線性增加趨勢(表8和表9)。同樣的,隨著日糧CP增加纈氨酸動靜脈濃度差呈線性增加趨勢(P=0.08)。第18天,隨著CP比例增加,賴氨酸和纈氨酸動靜脈濃度差呈線性增加趨勢(P<0.08),亮氨酸的動靜脈濃度差明顯呈線性增加趨勢(P=0.04),蘇氨酸動靜脈濃度差呈曲線增加趨勢(線性,P=0.06;二次方,P= 0.09)。第18天整個日糧運輸效率沒有改變。
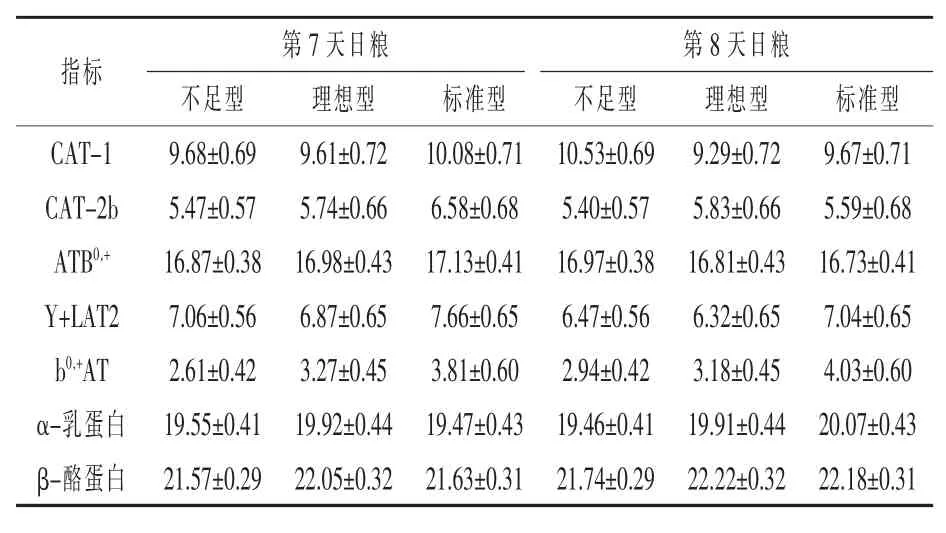
表6 通過反轉錄定量PCR(RT-qPCR)對編碼母豬乳腺組織中AA轉運蛋白和乳蛋白基因的mRNA豐度的定量分析
動脈BBCA∶Lys比例見表13。第7天隨著CP水平增加,血漿總BCAA∶Lys、Leu∶LYS和Ile∶Lys先降低后增加(二次方,P<0.03),同樣,血漿Val∶Lys也先下降再升高(線性,P=0.05;二次方,P= 0.09)。哺乳第7和18天不同日糧血漿Arg:Lys保持不變。
3 討論
大多數研究關注于優化哺乳母豬日糧賴氨酸攝取來最大化窩增重(Kim等,1999;Yang等,2000;Dourmad等,1998;NRC,1998),但很少有研究關注調節賴氨酸用于乳蛋白產量的利用效率這一機制。可以設計一個試驗方案,以最優化賴氨酸利用效率用于窩增重和減少氮素損失到環境中。第一步,賴氨酸用于奶產量的利用效率涉及乳腺細胞賴氨酸攝取,位于細胞膜上轉運蛋白介導的過程(Shennan和Peaker,2000;Shennan等,1997)。本研究評估飼喂NRC(1998)提出的最佳必需AA時,整個乳腺賴氨酸運輸是否受日糧CP影響。本研究中,在日增重、母豬體重損失、采食量的基礎上,預測不足型、理想型和標準型日糧的SID Lys需求量分別為0.82%,0.91%和0.85%(NRC,2012)。日糧包含0.60%、0.85%和1.11%的SIDLys分別表明不足型、理想型和標準型日糧中Lys供應為明顯缺乏、少量缺乏和過量。然而,尤其是對理想型日糧來說,這種預測并不是基于經驗而得出降低日糧粗蛋白質含量,很可能是本研究中理想型日糧需要增加賴氨酸利用率。例如,不足型、理想型和標準型日糧中凈賴氨酸攝取(數據未示出)與平均日增重比率估計分別為0.099、0.093和0.095。此研究中觀察到的日增重對應日糧蛋白質結果與Pérez-Laspiur等(2009)和Guan等(2004)報道類似。然而,在研究中,賴氨酸需要量僅達到18%日糧蛋白質;因此,日糧蛋白質水平超過曾報道的添加量(18%),日增重才能最大化。與此研究相似,Guan等(2004)報道,在最大平均日增重時,動靜脈濃度差、賴氨酸和精氨酸的提取效率最大。先前關于賴氨酸和精氨酸增加與較高的日增重相對應,這一機制研究是有限的。Pérez-Laspiur等(2009)報道隨著日糧Lys和CP從不足到過量逐漸增加,乳腺組織編碼CAT-2b的基因(SLC7A2)表達量呈線性降低。本試驗中,我們進一步探討了在飼喂理想型和低蛋白質日糧時,負責Lys和Arg轉運的其他基因是否與日糧Lys和Arg可利用率相對應。
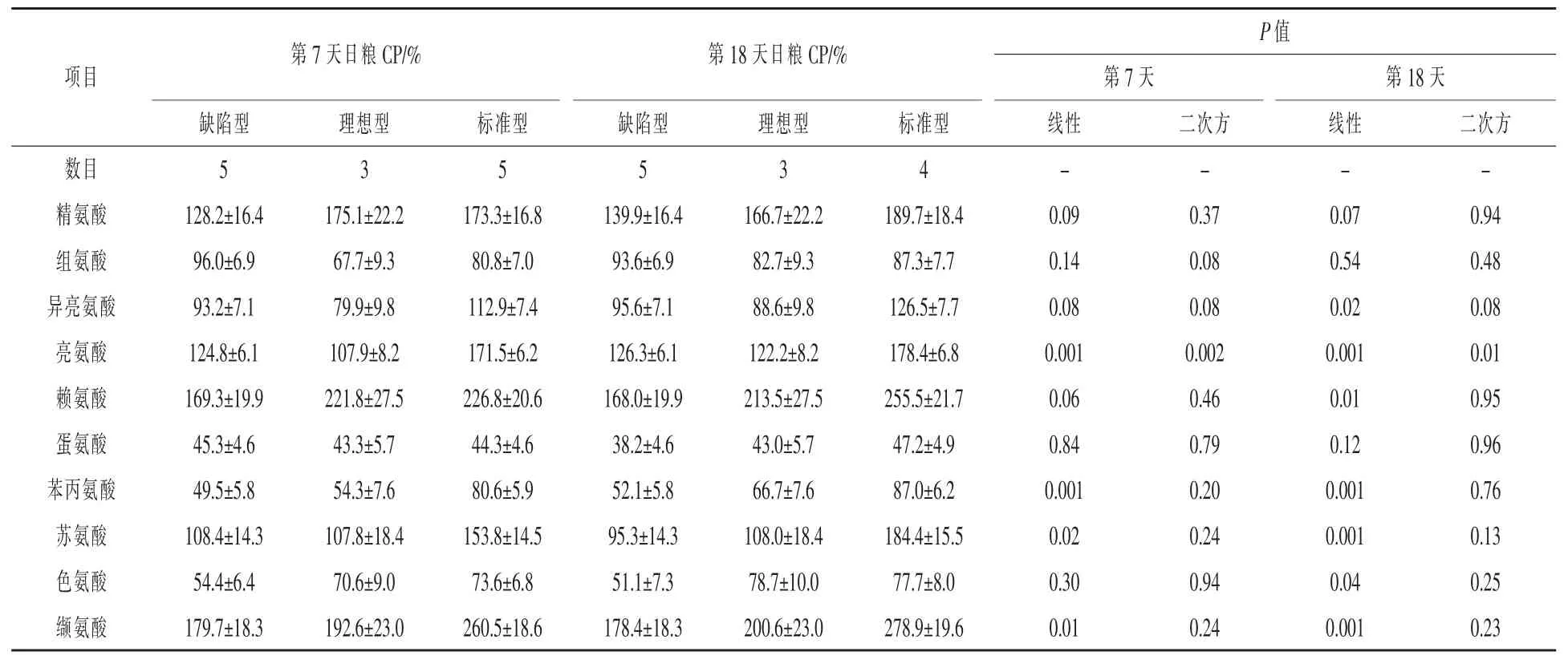
表7 日糧蛋白質水平對哺乳期第7天和第18天母豬動脈AA濃度的影響μmol/L
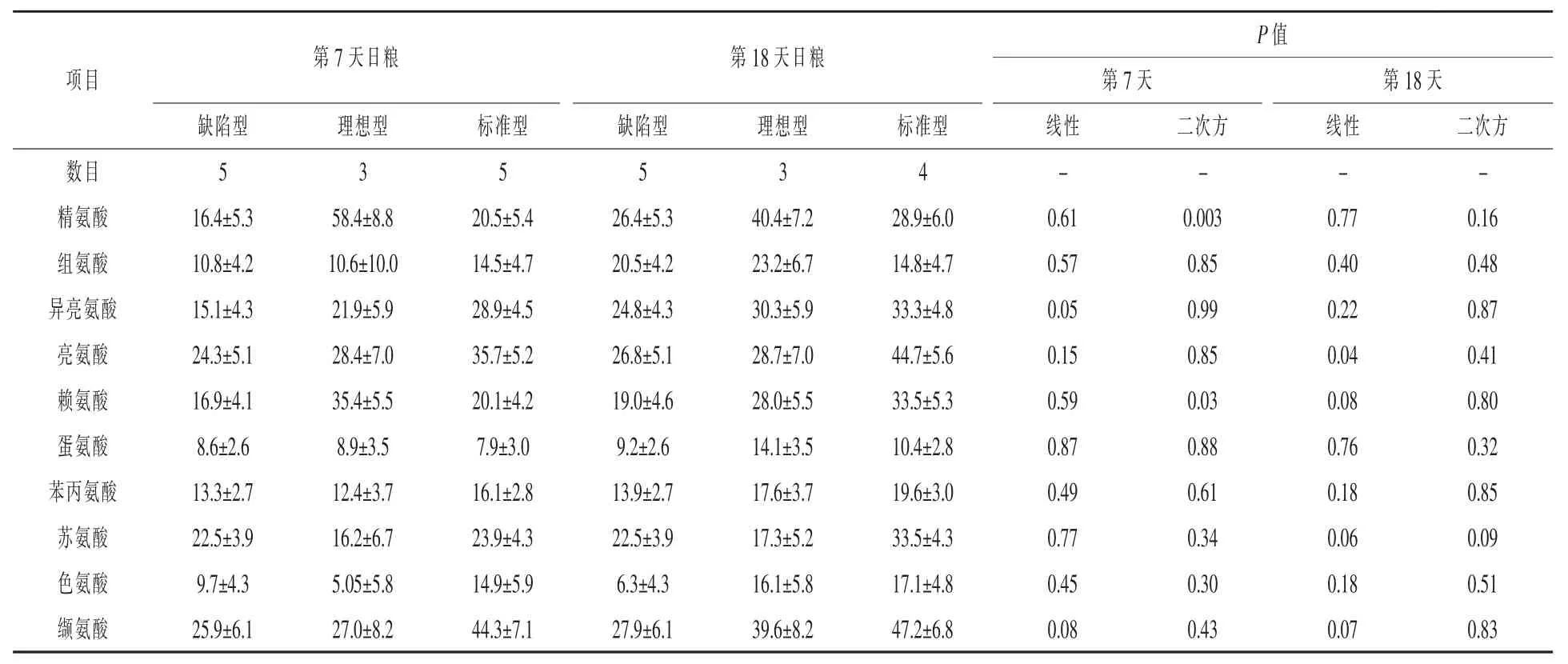
表8 日糧蛋白質水平對哺乳期第7天和第18天母豬乳腺動靜脈AA之差的影響μmol/L
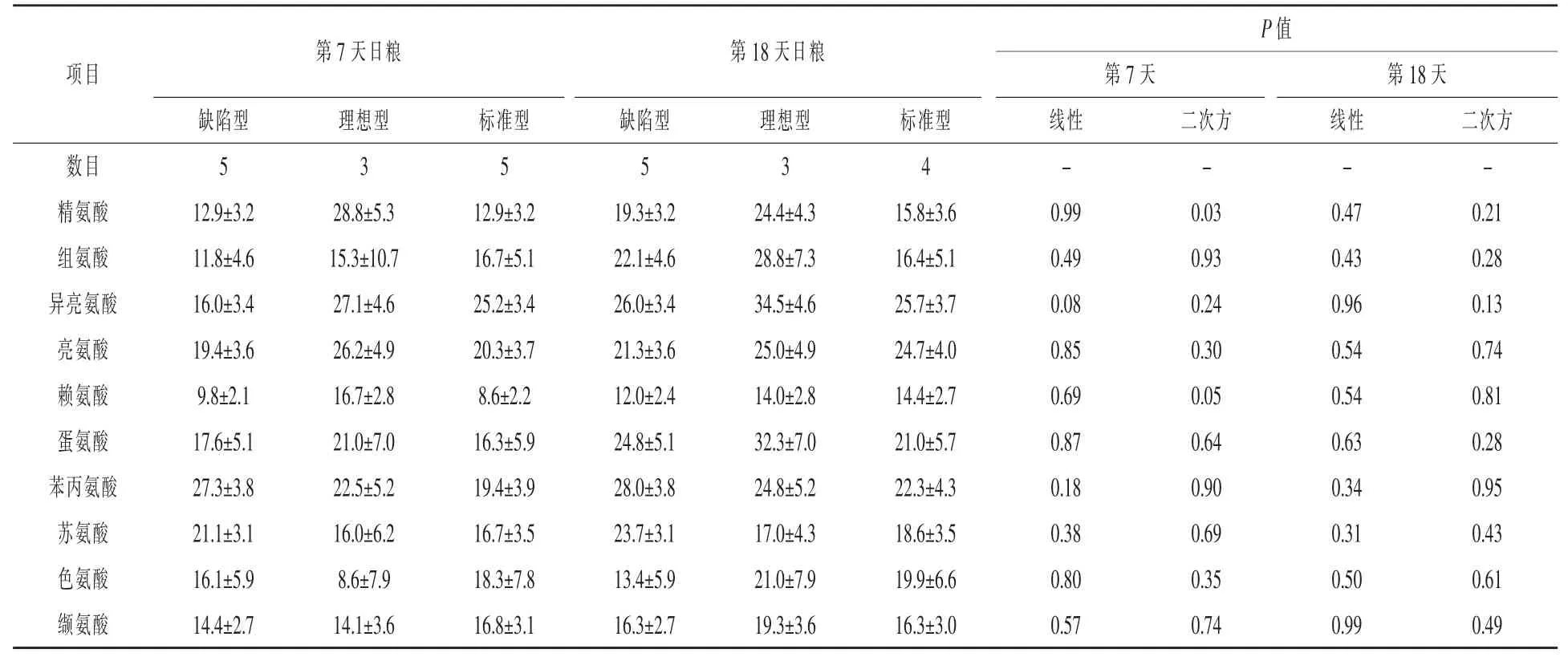
表9 日糧蛋白質水平對哺乳期第7和天第18天母豬乳腺AA運輸效率影響%
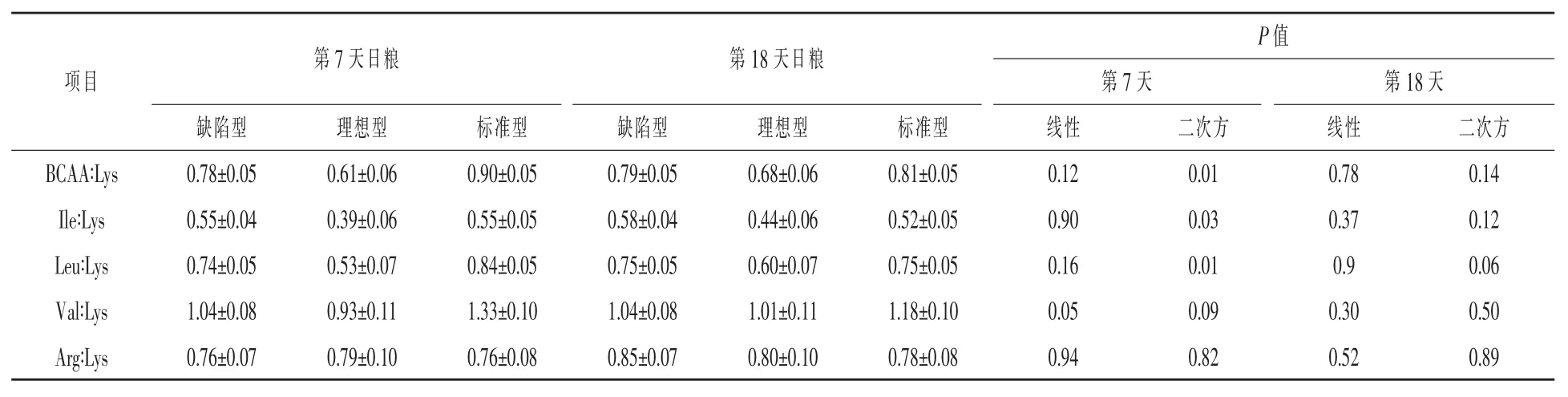
表10 哺乳期第7天和第18天日糧粗蛋白濃度對母豬乳腺動脈AA比率的影響
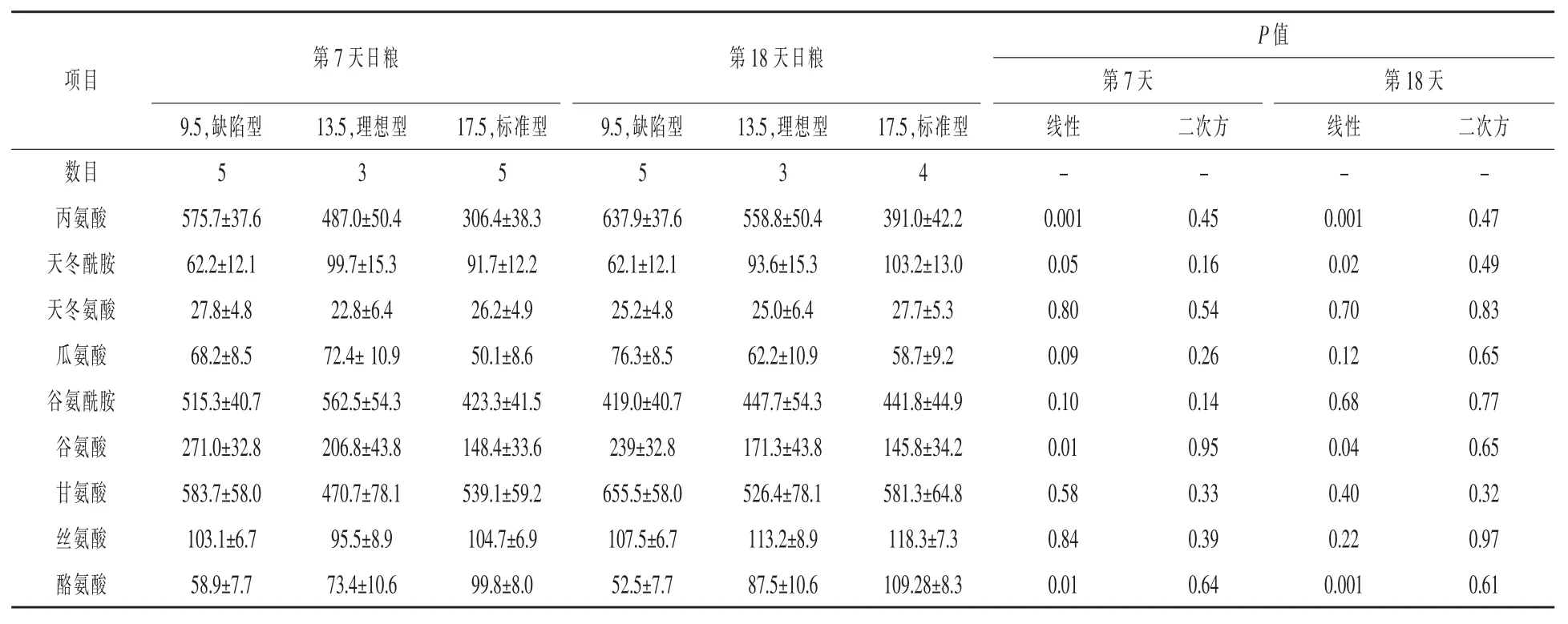
表11 日糧蛋白質水平對哺乳期第7天和第18天母豬動脈非必需AA濃度的影響μmol/L
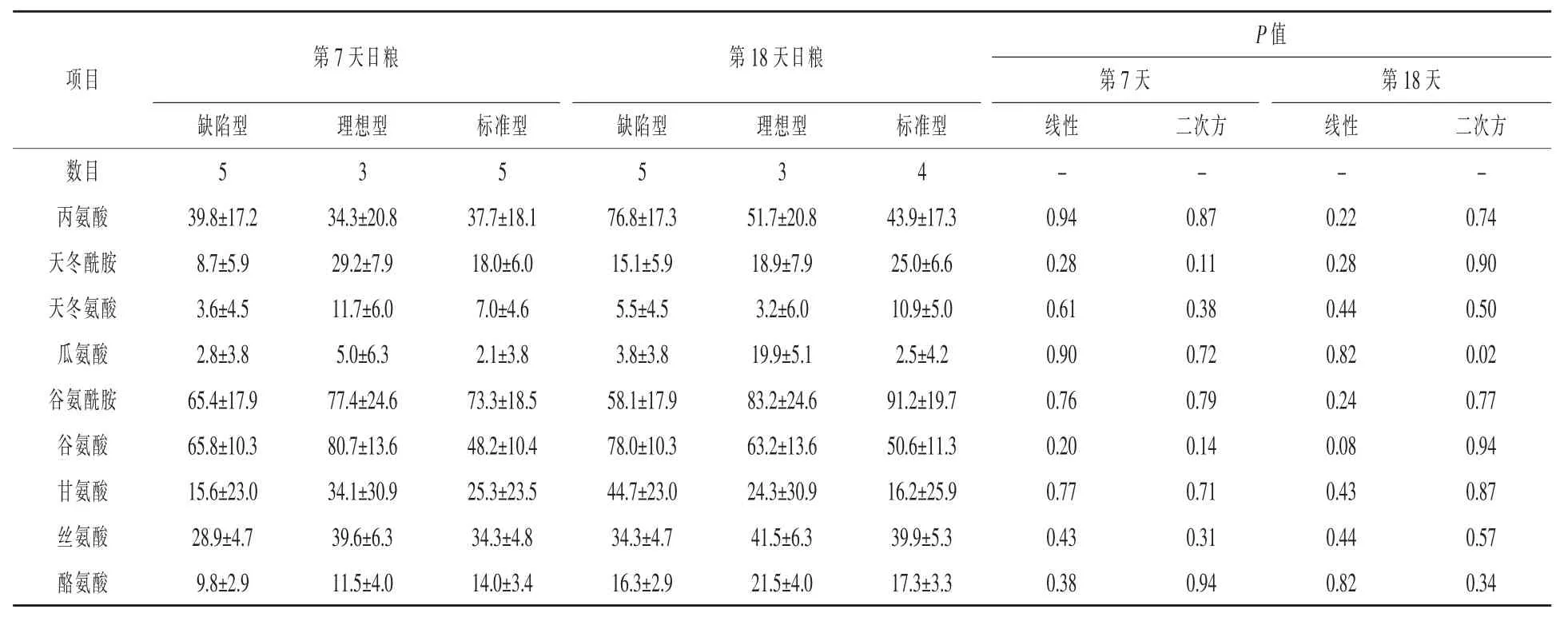
表12 日糧蛋白質水平對哺乳期第7天和第18天母豬乳腺動靜脈非必需AA差異的影響μmol/L
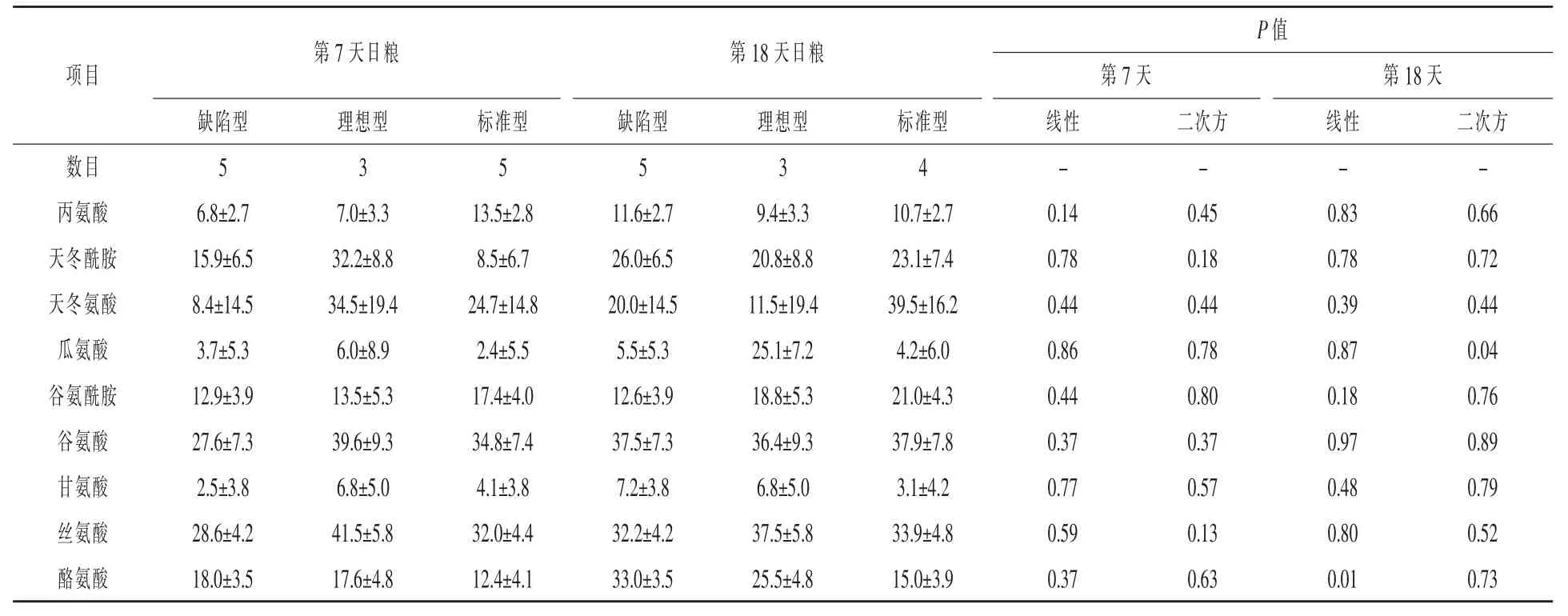
表13 日糧蛋白質水平對哺乳期第7天和第18天母豬乳腺非必需AA運輸效率的影響%
已知促進賴氨酸轉運到哺乳動物上皮細胞中的蛋白質包括y+系統中AA轉運蛋白家族特異性陽離子型AA(Shennan和Peaker,2000;Palacín等,1998),其中2個已經確定在哺乳母豬乳腺組織中,即CAT-1和CAT-2b(Pérez-Laspiur等,2009、2004)。賴氨酸也可通過與BCAA共享系統,如B0,+、b0,+和y+L系統,這些系統中,已有報道蛋白質ATB0,+、b0,+AT和y+LAT2在豬乳腺組織中表達,并在預翻譯水平調節(Manjarín等,2011;Pérez-Laspiur等,2009、2004)。在本研究中,隨著蛋白質從17.5%(標準型)降低為13.5%(理想想),Lys和Arg運輸效率增加,但不同日糧和整個泌乳階段CAT-1、CAT-2b、ATB0,+、b0,+和y+LAT2基因的表達不變。運輸效率的這種變化很可能與乳腺細胞數增加無關,因為不同日糧和整個哺乳期乳腺DNA濃度保持不變。在體內試驗(Pérez-Laspiur等,2009)和體外試驗(Satsu等,1998)表明,隨著CP攝入降低,CAT-2b和ATB0,+基因表達具有適應性調節。然而,本研究中,總賴氨酸(分析值)從1.24%(標準型)降到0.99%(理想型)或0.62%(不足型),可能不足以引起AA轉運蛋白基因表達的改變。Pérez-Laspiur等(2009)研究表明,日糧總賴氨酸自適應調節值為0.60%,與本試驗中不足型日糧氨基酸數值接近,但本試驗中SID賴氨酸濃度比Pérez-Laspiur等(2009)試驗中低。
同樣地,哺乳期第7天和第18天所有日糧中主導-α-乳清蛋白和β酪蛋白的轉錄物豐度保持不變,表明乳蛋白基因表達的變化發生在哺乳期早期或者在轉錄后(也就是蛋白質翻譯或表達)。事實上,我們先前的研究顯示豬乳腺組織賴氨酸轉運蛋白和乳蛋白基因是正調節,與產前和泌乳第5天發生的奶需求相對應(Manjarín等,2011)。此外,大鼠肌肉組織的研究表明,日糧AA可能通過激活哺乳動物納巴霉素(mTOR)的靶物質來調節mRNA翻譯和蛋白合成(Hundal和Taylor,2009;Kimball和Jefferson,2007;O’Connor等,2003)。在這方面,奶牛上研究表明mTOR途徑在泌乳期整個蛋白質翻譯和調節中起關鍵作用(Burgos等,2010;Hayashi等,2009;Hayashi和Proud,2007),mTOR途徑蛋白激酶的基因表達已在豬乳腺中被證實(Manjarín等,2011)。因此,本研究中觀察到的賴氨酸和精氨酸轉運效率和仔豬日增重的增加,可能是由于AA轉運蛋白基因在蛋白水平的正向調節。
乳腺中Lys運輸效率的變化與BCAA∶Lys比值的變化相反。與日糧蛋白質9.5%和17.5%相比,日糧蛋白質13.5%哺乳期第7天飼喂理想AA配方,乳腺BCAA∶Lys比值降低,Lys轉運效率升高。與此相反,泌乳第18天,所有日糧動脈BCAA∶Lys比值和Lys轉運效率保持不變。所有日糧SID BCAA∶Lys為常量時,動脈BCAA∶Lys比值變化的生理機制尚不清楚。飼喂理想型日糧的母豬可能BCAA外周組織攝入量較高,導致動脈BCAA:Lys比值降低。BCAA的代謝主要是發生在肝外組織,如肌肉和脂肪組織(Herman等,2010),是因為肝細胞缺乏參與支鏈氨基酸氧化的BCAA氨基轉移酶(Li等,2009;Nelson和Cox,2008)。事實上,與飼喂標準型和不足型日糧的母豬相比,飼喂理想型日糧的母豬體重損失少,表明骨骼肌蛋白活性低。
在泌乳母豬(Guan等,2002)和奶牛(Bequette等,1996)上可見,蛋白質攝入量增加,導致乳腺支鏈氨基酸運輸增加,而乳蛋白含量沒有增加。因此Bequette等(1996)推測乳腺過量吸收的支鏈氨基酸可能在組織中被氧化,有可能降低日糧氨基酸轉變為乳蛋白的利用效率。乳腺支鏈氨基酸氧化分解代謝,可通過氨基酸競爭活動對賴氨酸和精氨酸運輸進入乳腺細胞的效率產生負面影響。例如,氨基酸轉運蛋白ATB0,+,介導上皮細胞上陽離子和中性氨基酸的胞內運輸(Bro?r,2008),顯示了對支鏈氨基酸的更大親和力(Sloan和Mager,2013)。正如所提到的,本試驗和先前的研究(Pérez-Laspiur等,2009、2004)表明在豬乳腺組織中ATB0,+轉錄明顯較高。盡管所有日糧SID BCAA∶Lys比值相似,但哺乳期第7天標準型和不足型日糧動脈BCAA∶Lys比值明顯比理想型大,因此,不足型和標準型日糧動脈BCAA∶Lys比值的增加可能通過轉運蛋白ATB0,+增加BCAA相對于Arg和Lys的競爭吸收優勢,從而導致豬乳腺賴氨酸和精氨酸轉運效率降低。陽離子和中性AA共享氨基酸轉運蛋白y+LAT2,它在豬乳腺細胞也是高度表達的(Manjarín等,2011)。相對于ATB0,+,y+LAT2是陽離子和大分子中性AA之間的必需交換工具,促進陽離子氨基酸從細胞流出(Bro?r,2008)。在這方面,高水平的亮氨酸和纈氨酸可分別抑制賴氨酸攝取,使賴氨酸從大鼠乳腺外植體(Calvert和Shennan,1996;Shennan等,1994)和豬乳腺組織(Guan等,2002)流出增加。因此,除了通過ATB0,+優先攝取BCAA,不足型和標準型日糧動脈BCAA∶Lys比值的增加可能通過轉運蛋白y+LAT2促進Lys和Arg從乳腺細胞外流來交換BCAA,導致豬乳腺賴氨酸和精氨酸運輸效率下降。
哺乳期第7天飼喂理想型日糧的豬精氨酸運輸效率提高表明精氨酸對豬乳腺重要的作用。例如,Guan等(2004)報道,日糧蛋白質從24%過量值降低至18%需求量,動靜脈精氨酸濃度差增大。Nielsen等(2002)報道,乳腺攝取精氨酸的增加與窩產仔數增大相關。精氨酸被認為不能夠限制母豬乳蛋白合成(NRC,1998)。然而,近期研究表明,參與到乳腺組織的眾多功能AA直接與產奶量相關(Kim和Wu,2009)。精氨酸為血管擴張劑一氧化氮合成的底物(Wu和Morris,1998),血液流向乳腺是乳腺AA攝取增加的主要驅動變量,這與Nielsen等(2002)關于窩產仔數的研究結果相一致。然而,本研究中,理想型日糧中精氨酸較高的運輸效率,似乎與更大血流無關。血流量依據菲克原理(Guan等,2004)估算,奶中平均賴氨酸濃度為0.386%(賴氨酸占純乳蛋白7.01%,純乳蛋白占奶的5.41%)。Noblet和Etienne(1989)根據窩增重和仔豬初始體重計算,不足型、標準型和理想型日糧產奶量分別為每天8095 L、6188 L和5671 L。這些估算值與Guan等(2004)報道結果類似,飼喂蛋白不足或過量的日糧,母豬血流量更多。Guan等(2004)認為,乳腺通過增加血流量來補償賴氨酸動靜脈濃度差下降,促進非限制性氨基酸的吸收,從而提高代謝效率,并有可能增加能量需求。因此,不足型日糧可能是能量限制乳蛋白的合成,導致活性AA運輸降低。如Mateo等(2008)所述對于精氨酸提取,保持日糧精氨酸添加可以促進母豬產奶和仔豬的生長,因此乳腺組織精氨酸提取增加可以導致乳腺細胞精氨酸可用性增加。氨基酸轉運蛋白CAT-1是精氨酸進入乳腺組織的唯一通道。CAT-1對精氨酸和賴氨酸運輸具有高度特異性(Sloan和Mager,1999)。研究表明,乳腺組織中CAT-1 mRNA高豐度表達(Manjarín等,2011)。另外,精氨酸還參與多胺的合成,多胺是蛋白質合成和泌乳的調節物質(Meininger和Wu,2002;Wu和Morris,1998),脯氨酸是仔豬的一種必需氨基酸(Ball等,1986;Wu等,2011)。
綜上所述,日糧蛋白質從17.5%下降至13.5%,并補充晶體氨基酸,從而實現“理想”氨基酸配方,并沒有影響哺乳期仔豬日增重和編碼氨基酸轉運蛋白和乳蛋白的基因mRNA豐度。然而,與17.5%的日糧蛋白相比,泌乳早期(第7天)飼喂13.5%蛋白的日糧乳腺Lys和Arg運輸效率增加,血漿BCAA∶Lys比值降低。結果表明,日糧AA用于窩生長的效率與編碼乳蛋白或AA轉運蛋白的基因轉錄無關,可能與乳腺細胞膜上陽離子和支鏈氨基酸之間存在競爭性抑制有關。這些結果有助于我們理解飼喂最佳氨基酸平衡的日糧,以及N利用率提高的潛在機制,為農場更好地利用哺乳母豬理想蛋白質模型提供依據。
[1]Ball R O,Atkinson J L,Bayley H S.Proline as an essential amino acid for the young pig[J].British Journal of Nutrition,1986,55(3):659~668.
[2]Bequette B J,Metcalf J A,Wray-Cahen D,et al.Leucine and protein metabolism in the lactating dairy cow mammary gland:responses to supplemental dietary crude protein intake[J].Journal of Dairy Research,1996,63(2):209~222.
[3]Bionaz M,Loor J J.Identification of reference genes for quantitative realtime PCR in the bovine mammary gland during the lactation cycle[J].Physiological Genomics,2007,29(29):312~319.
[4]Burgos S A,Dai M,Cant J P.Nutrient availability and lactogenic hormones regulate mammary protein synthesis through the mammalian target of rapamycin signaling pathway[J].Journal of Dairy Science,2010,93(1):153~161.
[5]Calvert D T,Shennan D B.Evidence for an interaction between cationic and neutral amino acids at the blood-facing aspect of the lactating rat mammary epithelium[J].Journal of Dairy Research,1996,63(1):25~33.
[6]Dourmad J Y,Noblet J,Etienne M.Effect of protein and lysine supply on performance,nitrogen balance,and body composition changes of sows during lactation[J].Journal of Animal Science,1998,76(2):542~550.
[7]Guan X,Bequette B J,Calder G,et al.Amino acid availability affects amino acid flux and protein metabolism in the porcine mammary gland[J].Journal of Nutrition,2002,132(6):1224~1234.
[8]Guan X,Pettigrew J E,Ku P K,et al.Dietary protein concentration affects plasma arteriovenous difference of amino acids across the porcine mammary gland[J].Journal of Animal Science,2004,82(10):2953~2963.
[9]Hayashi A A,Nones K,Roy N C,et al.Initiation and elongation steps of mRNA translation are involved in the increase in milk protein yield caused by growth hormone administration during lactation[J].Journal of Dairy Science,2009,92(5):1889~1899.
[10]Hayashi A A,Proud C G.The rapid activation of protein synthesis by growth hormone requires signaling through mTOR[J].American Journal of Physiology Endocrinology and Metabolism,2007,292(292):E1647~1655.
[11]Herman M A,Pengxiang S,Peroni O D,et al.Adipose tissue branched chain amino acid(BCAA)metabolism modulates circulating BCAA levels[J]. Journal of Biological Chemistry,2010,285(15):11348~11356.
[12]Hundal H S,Taylor P M.Amino acid transceptors:gate keepers of nutrient exchange and regulators of nutrient signaling[J].American Journal of Physiology Endocrinology and Metabolism,2009,296(4):E603~613.
[13]Kim S W,Hurley W L,Han I K,et al.Changes in tissue composition associated with mammary gland growth during lactation in sows[J].Journal of Animal Science,1999,77(9):2510~2516.
[14]Kimball S R,Jefferson L S.New functions for amino acids:effects on gene transcription and translation[J].American Journal of Clinical Nutrition,2007,83(83):500~507.
[15]Kirkwood R N,Laspiur J P,Moore J B,et all.Mammary gland biopsy does not affect lactation performance in sows[J].Canadian Journal of Animal Science,2007,87(2):281~284.
[16]Labarca C,Paigen K.A simple,rapid,and sensitive DNA assay procedure[J].Analytical Biochemistry,1980,102(2):344~352.
[17]Larionov A,Krause A,Miller W.A standard curve based method for relative real time PCR data processing[J].Bmc Bioinformatics,2005,6(6):967~981.
[18]Li P,Knabe D A,Kim S W.Lactating porcine mammary tissue catabolizes branched-chain amino acids for glutamine and aspartate synthesis[J].Journal of Nutrition,2009,139(8):1502~1509.
[19]Li X,Rezaei R,Li P,et al.Composition of amino acids in feed ingredients for animal diets[J].Amino Acids,2011,40(40):1159~1168.
[20]Manjarin R,Steibel J P,Zamora V,et al.Transcript abundance of amino acid transporters,-casein,and-lactalbumin in mammary tissue of periparturient,lactating,and postweaned sows[J].Journal of Dairy Science,2011,94(7):3467~3476.
[21]Mateo R D,Wu G Y,Moon H K,et al.Effects of dietary arginine supplementation during gestation and lactation on the performance of lactating primiparous sows and nursing piglets[J].Journal of Animal Science,2008,86(4):827~835.
[22]Meininger C J,Wu G Y.Regulation of endothelial cell proliferation by nitric oxide[J].Methods in Enzymology,2002,352(24):280~295.
[23]Mikeska T,Dobrovic A.Validation of a primer optimisation matrix to improve the performance of reverse transcription-quantitative real-time PCR assays[J].Bmc Research Notes,2009,2(1):1~5.
[24]Nielsen T T,Trottier N L,Stein H H,et al.The effect of litter size and day of lactation on amino acid uptake by the porcine mammary glands[J].Journal of Animal Science,2002,80(9):2402~2411.
[25]Noblet J,Etienne M.Estimation of sow milk nutrient output[J].Journal of Animal Science,1989,67(67):3352~3359.
[26]Nutrient Requirements of Swine[M].1998.
[27]Nutrient Requirements of Swine[M].2012.
[28]O'Connor P M J,Kimball S R,Agus S,et al.Regulation of translation initiation by insulin and amino acids in skeletal muscle of neonatal pigs[J]. American Journal of Physiology Endocrinology and Metabolism,2003,285(1):E40~53.
[29]Otto E R,Yokoyama M,Ku P K,et al.Nitrogen balance and ileal amino acid digestibility in growing pigs fed diets reduced in protein concentration[J]. Journal of Animal Science,2003,81(7):1743~1753.
[30]Palacín M,Estévez R,Bertran J,et al.Molecular biology of mammalian plasma membrane amino acid transporters[J].Physiological Reviews,1998,78(4):969~1054.
[31]Pérez-Laspiur J P,Burton J L,Weber P S D,et al.Dietary protein intake and stage of lactation differentially modulate amino acid transporter mRNA abundance in porcine mammary tissue[J].Journal of Nutrition,2009,139(9):1677~1684.
[32]Pérez-Laspiur J P,Burton J L,Weber P S D,et al.Short communication:amino acid transporters in porcine mammary gland during lactation[J].Journal of Dairy Science,2004,87(10):3235~3237.
[33]Satsu H,Watanabe H,Arai S,et al.System B0,(+)-mediated regulation of lysine transport in Caco-2 human intestinal cells[J].Amino Acids,1998,14(4):379~384.
[34]Shennan D B,Mcneillie S A,Jamieson E A,et al.Lysine transport in lactating rat mammary tissue:evidence for an interaction between cationic and neutral amino acids[J].Acta Physiologica Scandinavica,1994,151(4):461~466.
[35]Shennan D B,Millar I D,Calvert D T.Mammary-tissue amino acid transport systems[J].Proceedings of the Nutrition Society,1997,56(1A):177~191.
[36]Shennan D B,Peaker M.Transport of milk constituents by the mammary gland[J].Physiological Reviews,2000,80(3):925~951.
[37]Sloan J L,Mager S.Cloning and functional expression of a human Na(+)and Cl(-)-dependent neutral and cationic amino acid transporter B(0+)[J]. Journal of Biological Chemistry,2013,8(7):410~414.
[38]Tramontana S,Bionaz M,Sharma A,et al.Internal controls for quantitative polymerase chain reaction of swine mammary glands during pregnancy and lactation[J].Journal of Dairy Science,2008,91(8):3057~3066.
[40]Trottier N L,Shipley C F,Easter R A.Plasma amino acid uptake by the mammary gland of the lactating sow[J].Journal of Animal Science,1997,75(5):1266~1278.
[41]Vandesompele J,Preter K D,Pattyn F,et al.Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes[J].Genome Biology,2002,3(7):research0034.0031-research0034.0011.
[42]Wu G Y,Bazer F W,Burghardt R C,et al.Proline and hydroxyproline metabolism:implications for animal and human nutrition[J].Amino Acids,2011,40(4):1053~1063.
[43]Wu G Y,Meininger C J.Analysis of Citrulline,Arginine,and Methylarginines Using High-Performance Liquid Chromatography[J].Methods in Enzymology,2008,440:177~189.
[44]Wu G Y,Morris S M.Arginine metabolism:Nitric oxide and beyond.Biochemical Journal,1998,336(1):1~17.
[45]Yang H,Pettigrew J E,Johnston L J,et al.Effects of dietary lysine intake during lactation on blood metabolites,hormones,and reproductive performance in primiparous sows[J].Journal of Animal Science,2000,78(4):1001~1009.
[45]Yuan J S,Reed A,Feng C,et al.Statistical analysis of real-time PCR data[J].Bmc Bioinformatics,2006,7(4):563~569.■
10.15906/j.cnki.cn11-2975/s.20161508
S816.7
A
1004-3314(2016)15-0032-12
猜你喜歡
瘋狂英語·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數理化(高中版.高考數學)(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經濟(2017年6期)2017-07-12 09:27:16
中國衛生(2016年9期)2016-11-12 13:27:54
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國洗滌用品工業(2015年7期)2015-02-28 19:02:38
電子設計工程(2015年12期)2015-02-27 12:06:10
中國衛生(2014年11期)2014-11-12 13:11:32
