黃河口及鄰近水域小型鳀鯡魚類數量分布及其與環境因子的關系
2016-11-11 01:23:30李敏紀毓鵬徐賓鐸薛瑩任一平
海洋學報 2016年10期
李敏,紀毓鵬,徐賓鐸*, 薛瑩,任一平
(1.中國海洋大學 水產學院, 山東 青島 266003)
?
黃河口及鄰近水域小型鳀鯡魚類數量分布及其與環境因子的關系
李敏1,紀毓鵬1,徐賓鐸1*, 薛瑩1,任一平1
(1.中國海洋大學 水產學院, 山東 青島 266003)
小型鳀鯡魚類多為浮游生物食性種類,又是多種高營養層次魚類的餌料生物,作為生態系統中的重要類群,其數量分布對多種生物均具有重要影響。本文根據2013年6月、8月、10月和2014年2月、4月、5月在黃河口及鄰近水域進行的漁業資源和環境調查數據,分析了該海域小型鳀鯡魚類資源數量分布特征,并用廣義可加模型研究了其數量分布與時空和環境因子等之間的關系。黃河口及鄰近水域小型鳀鯡魚類相對資源量呈現明顯的季節變化,夏、秋季相對資源量明顯高于冬、春季。小型鳀鯡魚類相對資源量具有明顯的空間變化,且在不同月份呈現不同的空間分布格局;在4月、5月、6月小型鳀鯡魚類主要分布于近岸水域,在8月、10月小型鳀鯡魚類分布外移且范圍擴大。GAM分析表明,影響黃河口及鄰近水域小型鳀鯡魚類相對資源量時空分布的主要環境因子包括表層鹽度、表層水溫、水深和浮游植物豐度等。小型鳀鯡魚類相對資源量隨表層鹽度的升高呈現升高的趨勢,在鹽度達到30時,小型鳀鯡魚類相對資源量處于最高水平。隨表層水溫的升高相對資源量有升高趨勢,當表層水溫達到15℃后,小型鳀鯡魚類相對資源量增加趨勢減緩且豐度較大。小型鳀鯡魚類相對資源量隨浮游植物豐度的增大總體呈現上升趨勢,而隨水深的變化呈現先降低后升高的趨勢。黃河口及鄰近水域小型鳀鯡魚類相對資源量的時空變化與鳀鯡魚類產卵、索餌和越冬洄游有關,受到季節變化、黃河徑流變化以及黃河調水調沙等引起的海洋環境因子變動的影響。
時空分布;相對資源量;環境因子;廣義可加模型
1 引言
魚類對海洋環境因素的適應性和局限性決定了其洄游和分布水域,深入研究魚類數量分布及其與相關海洋環境因子的關系,將會為其漁況分析、漁場尋找和漁業資源合理利用等提供理論基礎[1]。隨著計算機技術和統計學的發展,有關漁業捕撈對象的數量分布與環境因子之間關系的研究越來越多,但多數是針對單魚種的研究,例如方氏云鳚、鳀魚、黃鯽和鮐魚等[2—5],而生態位相近的魚類功能群的數量分布與環境因子關系的研究尚不多見。
20世紀80年代以來,海域高強度開發、海洋環境退化以及過度捕撈等人類活動造成黃河口水域的漁業資源生產潛力嚴重下降[6]。漁業資源結構發生很大變化,帶魚(Trichiuruslepturus)、小黃魚(Pseudosciaenapolyactis)和鲆鰈類等底層魚類日益衰退,鳀魚(Engraulisjaponicus)、斑鰶(Clupanodonpunctaus)等小型中上層鳀鯡魚類成為該海域優勢種[7—8]。
鳀鯡魚類屬于脊索動物門(Chordata),硬骨魚綱(Osteichthyes),鯡形目(Clupeiformes),鯡科(Clupeidae)和鳀科(Engraulidae)。其中,黃河口小型鳀鯡魚類中鯡科主要包括青鱗小沙丁魚(Sardinellazunasi)和斑鰶(Clupanodonpunctaus)等;鳀科主要包括鳀魚(Engraulisjaponicus)、赤鼻棱鳀(Thrissakammalensis)、黃鯽(Setipinnagilberti)和中頜棱鳀(Thrissamustax)等[9—10]。小型鳀鯡魚類性成熟早,僅1年即可性成熟,資源補充迅速;其多為暖水種或暖溫種,繁殖季節、繁殖習性和食性等相似[11—12]。小型鳀鯡魚類既是多種高營養級經濟魚類的餌料,又可作為漁業捕撈對象,在食物網中起著承上啟下的作用。因此,小型鳀鯡魚類是黃河口水域生態系統的關鍵類群[13—14]。但近年來,海洋環境惡化和過度捕撈等人為因素造成小型鳀鯡魚類漁業資源出現不同程度的衰退,太平洋鯡(Clupeapallasii)和青鱗小沙丁魚等種類的漁業產量自2000年以來均有不同程度的下降[14—15]。隨著環境因素的變化,生物群落的種類組成會有顯著變化[16],但以食物資源利用方式為基礎的功能群組成卻相對穩定[17]。功能群比單種類能更有效地反映生境梯度變化和生態系統功能[18]。因此,環境因子的改變引起的功能群的數量變動,更能體現生態系統對外界環境變動的實際響應。
作為重要魚類功能群,小型鳀鯡魚類數量及空間分布的變動可能會對生態系統中多種生物產生重要影響。因此,需要分析小型鳀鯡魚類數量分布及其與環境因子的關系。本文根據2013—2014年6個航次的漁業資源和環境調查數據,研究小型鳀鯡魚類數量分布及其與相關環境因子的關系,以期為黃河口及鄰近水域小型鳀鯡魚類資源養護與可持續利用提供理論依據。
2 材料與方法
2.1數據來源
本文中小型鳀鯡魚類相對資源量和環境因子數據來自2013年6月、8月、10月,2014年2月、4月、5月在黃河口及其鄰近海域進行的漁業資源和環境調查。本次調查范圍為37.60°~38.20°N,119.00°~119.80°E之間的海域,調查站位的設計以黃河入海口為中心,向外呈輻射狀設置5條斷面,中間3條斷面各有4個站位,兩側2條斷面各設置3個站位,河口附近的站位較密集,調查站位設置18個站點(圖1)。調查船為單拖漁船,船只功率為260 kW,調查網具網口寬度為8 m,網囊囊目為20 mm。漁業資源調查每站平均拖網時間為1 h,拖速為2.0 ~3.0 kn。同步調查的環境因子包括水溫、鹽度、深度和pH等,其中溫度、鹽度、深度采用CTD溫鹽深儀(XR-420)測定。樣品的采集和測定根據《海洋調查規范》(GB/T 12763.6—2007)進行[19]。在數據分析前對原始調查數據進行預處理,將各站位鳀鯡魚類的相對資源量均換算成拖網時間1 h、拖速2.0 kn的漁獲數值。
2.2數據分析
相關分析表明表層水溫和底層水溫相關性極顯著,且以往對鳀魚等小型鳀鯡魚類的研究中皆選取表層水溫作為環境因子[3]。夏季河口受沖淡水的影響,表層鹽度變化更明顯。因此,本研究選取表層水溫、表層鹽度、表層溶解氧和水深作為環境因子;葉綠素a、浮游動物豐度和浮游植物豐度作為生物因子;月份作為時間因子;經度、緯度作為空間因子。在分析相對資源量時空變化與相關因子的關系前,首先對因子進行Pearson相關性分析,顯著相關的兩個因子之間選擇其一。初步篩選出7個因子,包括經度、緯度、表層水溫、表層鹽度、水深、浮游動物豐度和浮游植物豐度。
利用GAM模型對黃河口小型鳀鯡魚類相對資源量和選取的相關環境因子進行分析,GAM模型的一般表達式如下[20]:
(1)
式中,Y為相對資源量,以單位網次漁獲量(單位:g/h)表示,即各調查站位拖網時間1 h及拖速2 kn時的漁獲質量;xj表示解釋變量,即各站位的影響因子;α是適合函數的截距,ε表示殘差,fi(xj)表示各自變量的任意單變量函數,為樣條平滑函數(Spline smoothing)。模型分析的誤差函數均為正態分布,連接函數為自然對數[21]。
將初步篩選出的7個因子代入GAM模型,進一步篩選出影響小型鳀鯡魚類數量分布的主要因子。根據AIC準則,在AIC最小的單因子預測函數的基礎上按順序加入其他因子,進而得到AIC值最小的雙因子預測模型,再依照上面的過程不斷重復,直到繼續添加新的因子不會減小AIC值為止,所得AIC值最小的模型即為擬合效果最好的模型[21—22]。

圖1 黃河口及鄰近海域漁業資源與環境調查站位Fig.1 Map of bottom trawl survey stations of fishery resources in Huanghe River Estuary and its adjacent waters
利用F檢驗評估預測變量的顯著性[21—22]。
模型構建過程均在R統計軟件(version: 2.15)的gam軟件包中實現[22],并利用Sufer11軟件繪制黃河口及鄰近海域鳀鯡魚類相對資源量的空間分布圖。
3 結果
3.1相對資源量的月變化
黃河口及鄰近水域小型鳀鯡魚類相對資源量呈現明顯的月變化,8月相對資源量最大,其次是10月,兩個月份相對資源量均在1 000 g/h以上;2月在該水域未捕到鳀鯡魚類(表1)。鳀鯡魚類相對資源量具有明顯的空間變化,且在不同月份呈現不同的空間分布規律。其中,2月該海域未捕獲到鳀鯡魚類;4月在119.25°~119.35°E的河口東部有少量分布;5月開始鳀鯡魚類的分布范圍擴大,在5個斷面均有分布,在遠離河口的東北部近海分布相對較多;6月在正對河口、朝向東北方向的2個斷面上鳀鯡魚類分布明顯減少;8月鳀鯡魚類在18個站位均有分布,除河口區海域分布較少外,其他站位分布較其他月份更為均勻,且相對資源量明顯增加;10月鳀鯡魚類主要集中在河口區域,河口以北的4個斷面,在遠離河口的海域鳀鯡魚類分布較少,而河口以東斷面上遠離河口的海域,鳀鯡魚類分布較多(圖2)。

表1 黃河口小型鳀鯡魚類相對資源量的月變化(平均值±標準誤差)
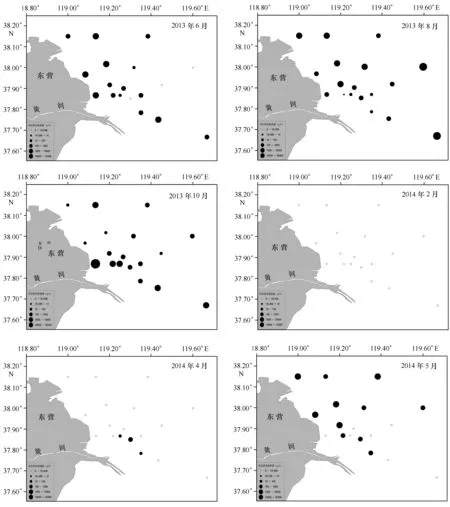
圖2 黃河口及鄰近海域小型鳀鯡魚類相對資源量的空間分布Fig.2 Spatial distribution of relative abundance of small-sized fish species in Clupeidae and Engraulidae in Huanghe River Estuary and its adjacent waters
3.2不同因子對黃河口小型鳀鯡魚類數量分布的影響
表2列出了黃河口小型鳀鯡魚類數量分布的影響因子的GAM篩選過程。根據AIC原則篩選后的GAM最優模型最終表達式如下:
Ln(Y+1)=α+S1(sss)+S2(depth)+
(2)
式中,Y是鳀鯡魚類相對資源量,sss是表層鹽度(sea surface salinity)、depth是水深(depth)、phy是浮游植物豐度(phytoplankton abundance)、sst是表層水溫(sea surface temperature)、lon是經度(longitude)。各因子的偏差解釋率分別為19.84%、5.73%、20.66%、5.92%和6.14%,所選因子對鳀鯡魚類相對資源量的累計解釋偏差為58.29%。其中貢獻最大的因子為浮游植物豐度,其次為表層鹽度,對相對資源量影響程度較小的因子包括水深、表層水溫和經度。GAM方差分析表明,表層鹽度對小型鳀鯡魚類相對資源量變化的影響極顯著(P<0.001),水深和浮游植物豐度對鳀鯡魚類相對資源量變化的影響顯著(P<0.05)(表3)。GAM模型的Q-Q殘差圖表明,預測值接近正態分布,模型擬合程度較高(圖3)。
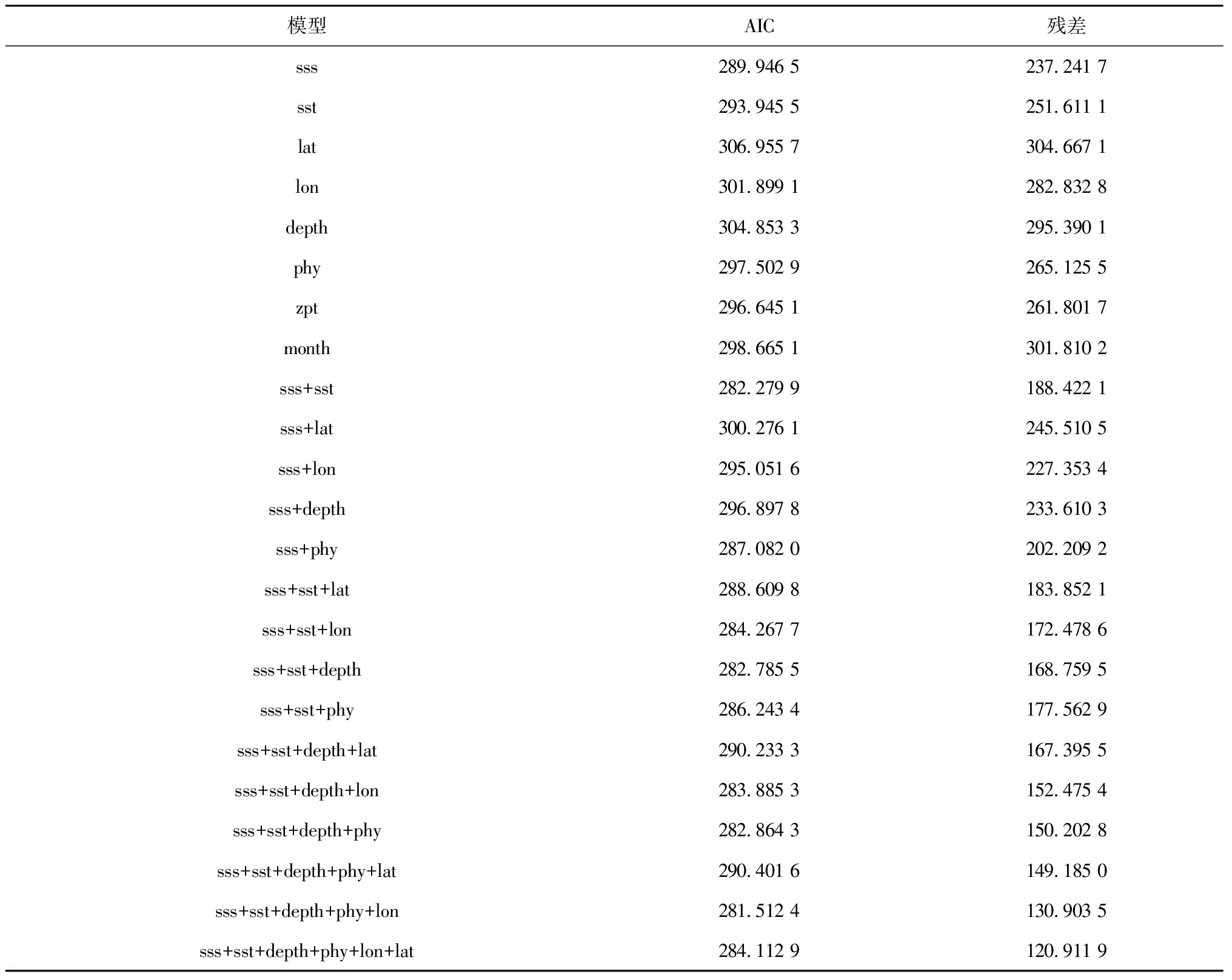
表2 黃河口小型鳀鯡魚類相對資源量分布的影響因子篩選過程
注: sst 表示表層水溫;sss 表示表層鹽度;depth為水深;phy表示浮游植物豐度;zpt表示浮游動物豐度;month 表示月份;lon 為調查站位的經度;lat 為調查站位的緯度。
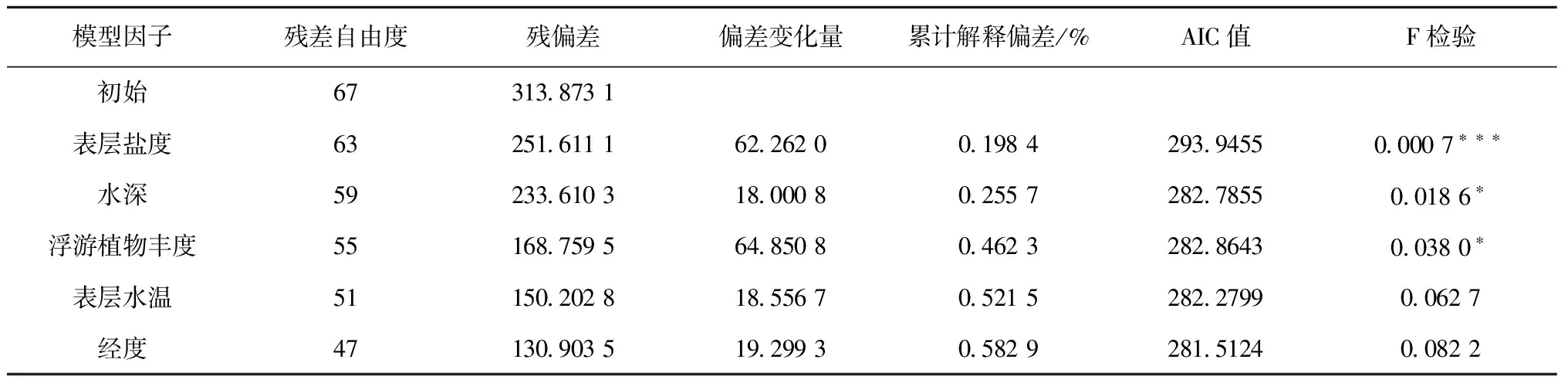
表3 GAM模型擬合結果的偏差分析
注:*P<0.05,***P<0.001。
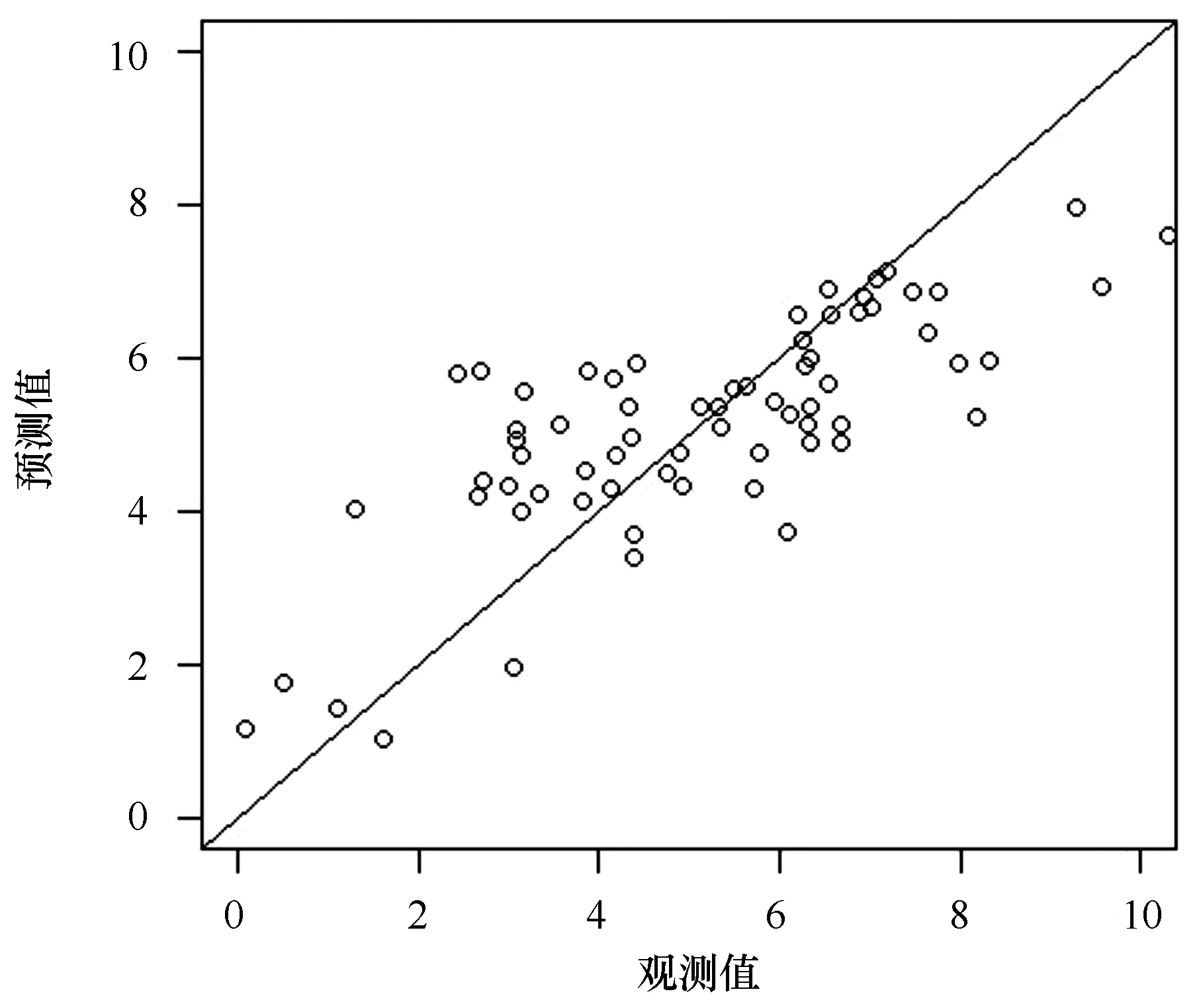
圖3 GAM模型的Q-Q 殘差圖Fig.3 Q-Q residual plot of GAM
圖4表明了黃河口及鄰近水域小型鳀鯡魚類相對資源量隨表層鹽度、表層水溫、水深、浮游植物豐度和經度的變化情況。本研究中,表層鹽度變化范圍為8~30,小型鳀鯡魚類的相對資源量隨表層鹽度的增加呈上升趨勢,當表層鹽度達到30時,相對資源量最大。在海水表層水溫較低(小于10℃)時,小型鳀鯡魚類有少量分布,主要分布水溫范圍為13~27℃。在表層水溫達到15℃時,鳀鯡魚類相對資源量出現峰值;在小于15℃時,相對資源量隨水溫的上升明顯增大;大于15℃時上升趨勢變緩。當水深9~17 m時,鳀鯡魚類相對資源量較小且較穩定;小于9 m時,相對資源量隨深度增加而減小;大于17 m,相對資源量隨水深的增加而增大。鳀鯡魚類相對資源量隨浮游植物豐度的變化呈現先下降后上升的趨勢,但除個別值外,浮游植物豐度的增加并未對鳀鯡魚類相對資源量的變化造成明顯影響。鳀鯡魚類相對資源量隨經度的變化總體呈上升趨勢,在119°~119.5°E范圍內,相對資源量略有波動,呈先上升后下降的趨勢;在119.5°E以外的海區,相對資源量呈增大趨勢。黃河口及鄰近水域小型鳀鯡魚類的相對資源量隨環境因子的變化呈現明顯的空間變化,表層鹽度與表層水溫對鳀鯡魚類相對資源量的增大有積極影響,鳀鯡魚類相對資源量隨水深和浮游植物豐度的變化都呈現先降低后升高的趨勢。
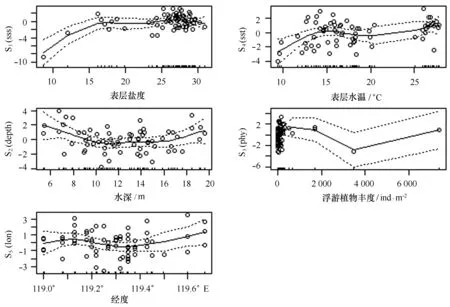
圖4 解釋變量對黃河口小型鳀鯡魚類相對資源量的影響Fig.4 Effects of explanatory variables on relative abundance of small-sized fish species in Clupeidae and Engraulidae in Huanghe River Estuary
4 討論
黃河口及其鄰近水域小型鳀鯡魚類的數量分布呈現明顯的季節變化,夏、秋季相對資源量明顯高于冬、春季。鳀魚、青鱗小沙丁魚、斑鰶和黃鯽等小型鳀鯡魚類春季洄游至渤海各海灣及河口繁殖[11,23—25],隨著水溫的升高,產卵群體的生物量逐步增加,到7月、8月新生群體補充進來,加之6-8月的伏季休漁大大減少了捕撈死亡率,造成了8月小型鳀鯡魚類相對資源量最高。再者,從食物關系的角度,夏季藍點馬鮫等洄游性魚類到達該水域的種類較少[26—27],鳀鯡魚類等小型魚類被捕食的機會減少,也是造成8月鳀鯡魚類相對資源量達到峰值的原因之一。
春季鳀鯡魚類分布范圍逐漸增大,夏、秋季節鳀鯡魚類分布最廣,由河口向外海擴展。其中,8月和10月分布范圍擴大,且分布較均勻。這是由于夏、秋季節水溫升高、鳀鯡魚類攝食能力增強,由近岸游向外海[28]。其次,夏、秋季降雨增多,沖淡水勢力強,河口區及鄰近水域鹽度較低,8月河口區及鄰近水域存在大量低鹽區。對應GAM模型的分析結果,高鹽區鳀鯡魚類相對資源量較大,鹽度達到30時,鳀鯡魚類相對資源量最高,這與夏、秋季節鳀鯡魚類由近岸向離岸深水區移動的現象一致。相關研究表明該海區優勢種鳀魚適合生長的鹽度31、斑鰶適合生長的鹽度26.2,本研究結果與上述適宜鹽度值接近[11,29]。
鳀鯡魚類的相對資源量和分布范圍在調查海域內也呈現明顯的空間變化。全年調查中,鳀鯡魚類相對資源量在近岸水域明顯高于離岸水域,且除2月以外,河口區全年分布頻率高于離岸水域。由于黃河口改道造成沙嘴前沿出現強潮流帶,將沙嘴向海中推進,并使潮流帶進一步強化[30]。而黃河入海徑流攜帶的淡水、泥沙以及營養鹽的長期相互作用使得近岸海域水深較淺、全年平均水溫較高、水中營養鹽較豐富。獨特的生態環境,為浮游生物提供了有利的生長條件[31],為以浮游生物為食的鳀魚等優勢種類提供了大量的餌料生物[31—32],有利于其產卵后在附近海域索餌、育肥[11,28]。
GAM模型大多用于解決非線性問題,在分析各因子對鳀鯡魚類相對資源量的影響時,雖然浮游植物豐度對鳀鯡魚類相對資源量的影響顯著,但模型的擬合結果并不十分理想,在GAM模型中的偏差解釋率較大。這與浮游植物種群增長速度快且生長周期短所造成的數據不連續有關[33]。另外,鳀鯡魚類主要攝食硅藻中的圓篩藻、曲舟藻、舟形藻等幾種浮游植物而非全部種類也可能是造成誤差的原因[11,28]。再者,眾多研究[11—12]表明,浮游動物是鳀鯡魚類的重要餌料組成部分,但在本文的模型篩選過程中,浮游動物豐度不是影響該海域小型鳀鯡魚類數量分布的顯著因子,這可能與本次調查中85%的調查站位的浮游動物豐度小于2 000個/m3有關,浮游動物豐度梯度較小有關。
除此之外,一定程度上黃河口及鄰近海域環境還受到渤海-萊州灣沿岸流的影響。沿岸流由渤海西南部沿岸的海河、黃河徑流入海形成,除渤海灣沿岸水春季沿渤海岸向北流動外,其他季節均與萊州灣沿岸低鹽水匯合[34],對黃河口及鄰近海域鹽度的變化也造成一定影響。夏、秋季節降雨增多,黃河徑流量增大,黃河口附近沿岸海域淡水堆積較多,河流從陸上帶來的泥沙和有機質的數量比較多[35],其中攜帶的有機碎屑也為斑鰶等碎屑食性的鳀鯡魚類提供豐富的餌料[11]。另外,鳀鯡魚類多棲息于軟沙底質水域底層[36],而黃河口入海口附近常年受到入海徑流攜帶的泥沙堆積作用,底質多為粉砂質沙[37],為鳀鯡魚類提供了較為理想的棲息場所。查明小型鳀鯡魚類數量分布對于水溫的響應關系,對于全球氣候變化引起的水溫升高情況下的小型鳀鯡魚類資源的利用具有理論指導意義。除了本研究中提到的上述影響因子外,底質類型等因子也可能對鳀鯡魚類的分布有一定影響。在進一步的研究中,需綜合考慮上述因子的影響。
[1]陳新軍. 漁業資源與漁場學[M]. 2版. 北京: 海洋出版社, 2014.
Chen Xinjun. Fishery Resources and Fishery Oceanograghy[M]. 2nd ed. Beijing: China Ocean Press, 2014.
[2]李敏, 李增光, 徐賓鐸, 等. 時空和環境因子對海州灣方氏云鳚資源豐度分布的影響[J]. 中國水產科學, 2015, 22(4): 812-819.
Li Min, Li Zengguang, Xu Binduo, et al. Effects of spatiotemporal and environmental factors on the distribution and abundance ofPholisfangiin Haizhou Bay using a generalized additive model[J]. Journal of Fishery Sciences of China, 2015, 22(4): 812-819.
[3]牛明香, 李顯森, 趙庚星. 黃海中南部越冬鳀魚空間分布及其與水溫年際變化的關系[J]. 應用生態學報, 2012, 23(2): 552-558.
Niu Mingxiang, Li Xiansen, Zhao Gengxing. Spatial distribution of winteringEngraulisjaponicusand its relationship with the inter-annual variations of water temperature in central and southern Yellow Sea[J]. Chinese Journal of Applied Ecology, 2012, 23(2): 552-558.
[4]劉勇, 程家驊, 李圣法. 東海區黃鯽數量分布特征的分析研究[J]. 海洋漁業, 2004, 26(4): 255-260.
Liu Yong, Cheng Jiahua, Li Shengfa. A study on the distribution ofSetipinnatatyin the East China Sea[J]. Marine Fisheries, 2004, 26(4): 255-260.
[5]鄭波, 陳新軍, 李綱. GLM和GAM模型研究東黃海鮐資源漁場與環境因子的關系[J]. 水產學報, 2008, 32(3): 379-386.
Zheng Bo, Chen Xinjun, Li Gang. Relationship between the resource and fishing ground of mackerel and environmental factors based on GAM and GLM models in the East China Sea and Yellow Sea[J]. Journal of Fisheries of China, 2008, 32(3): 379-386.
[6]張繼民, 劉霜, 尹韋翰, 等. 黃河口區域綜合承載力評估指標體系初步構建及應用[J]. 海洋通報, 2012, 31(5): 496-501.
Zhang Jimin, Liu Shuang, Yin Weihan, et al. Primary indicators of integrated carrying capacity in the region of the Yellow River Estuary and its application[J]. Marine Science Bulletin, 2012, 31(5): 496-501.
[7]朱鑫華, 繆鋒, 劉棟, 等. 黃河口及鄰近海域魚類群落時空格局與優勢種特征研究[J]. 海洋科學集刊, 2001(43): 141-151.
Zhu Xinhua, Miao Feng, Liu Dong, et al. Spatio-temporal pattern and dominant component of fish community in the Yellow River Estuary and its adjacent waters[J]. Studia Marina Sinica, 2001(43): 141-151.
[8]Jin Xianshi. Long-term changes in fish community structure in the Bohai Sea, China[J]. Estuarine, Coastal and Shelf Science, 2004, 59(1): 163-171.
[9]陳大剛. 黃渤海漁業生態學[M]. 北京: 海洋出版社, 1991.
Chen Dagang. Fisheries ecology of Yellow Sea and Bohai Sea[M]. Beijing: China Ocean Press, 1991.
[10]張世義. 中國動物志: 硬骨魚綱 鱘形目 海鰱目 鯡形目 鼠鮚目[M]. 北京: 科學出版社, 2001.
Zhang Shiyi. China fauna: Osteickthyes, Acipenseriformes, tarpon mesh, Clupeiformes, mouse oyster mesh[M]. Beijing: Science Press, 2001.
[11]陳大剛. 斑鰶[J]. 水產科技情報, 1975(9): 29-30.
Chen Dagang. The spotted gizzardshad[J]. Fisheries Science & Technology Information, 1975(9): 29-30.
[12]鄧景耀, 姜衛民, 楊紀明, 等. 渤海主要生物種間關系及食物網的研究[J]. 中國水產科學, 1997, 4(4): 1-7.
Deng Jingyao, Jiang Weimin, Yang Jiming, et al. Species interaction and food web of major predatory species in the Bohai Sea[J]. Journal of Fishery Sciences of China, 1997, 4(4): 1-7.
[13]焦燕, 陳大剛, 劉群, 等. 萊州灣小型鳀鯡魚類的生物學特征[J]. 水產學報, 2001, 25(4): 323-329.
Jiao Yan, Chen Dagang, Liu Qun, et al. Biological characteristics of some small species in Engraulidae and Clupeidae[J]. Journal of Fisheries of China, 2001, 25(4): 323-329.
[14]任一平, 劉群, 李慶懷, 等. 青島近海小型鳀鯡魚類漁業生物學特性的研究[J]. 海洋湖沼通報, 2002(1): 69-74.
Ren Yiping, Liu Qun, Li Qinghuai, et al. Biological characteristics of some small species in Engraulidae and Clupeidae[J]. Transactions of Oceanology and Limnology, 2002(1): 69-74.
[15]農業部漁業局. 中國漁業統計年鑒[M]. 北京: 中國農業出版社, 2013.
Ministry of Agriculture Bureau. China Fishery Statistical Yearbook[M]. Beijing: China Agriculture Press, 2013.
[16]Garrison P G. Spatial and dietary overlap in the Georges Bank groundfish community[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2000, 57(8): 1679-1691.
[17]張波, 李忠義, 金顯仕. 渤海魚類群落功能群及其主要種類[J]. 水產學報, 2012, 36(1): 64-72.
Zhang Bo, Li Zhongyi, Jin Xianshi. Functional groups of fish assemblages and their major species in the Bohai Sea[J]. Journal of Fisheries of China, 2012, 36(1): 64-72.
[18]Gaudêncio M J, Cabral H N. Trophic structure of macrobenthos in the Tagus Estuary and adjacent coastal shelf[J]. Hydrobiologia, 2007, 587(1): 241-251.
[19]國家質量監督檢驗檢疫總局. GB/T 12763.6-2007, 海洋調查規范 第6部分: 海洋生物調查[S]. 北京: 中國標準出版社, 2008.
General Administration of Quality Supervision, Inspection and Quarantine. GB/T 12763.6-2007, Specifications for oceanographic survey-part 6: marine biological survey[S]. Beijing: China Standards Press, 2008.
[20]Hastie T J, Tibshirani R J. Generalized Additive Models[M]. London: Chapman and Hall, 1990.
[21]Akaike H. Information theory and an extension of the maximum likelihood principle[C]// Petrov B N, Csaki F. Proceedings of the 2nd International Symposium on Information Theory. Budapest: Akademiai Kiado, 1973.
[22]Burnham K P, Anderson D R. Model selection and multimodel inference: a practical information-theoretic approach[M]. New York: Springer Verlag, 2002.
[23]葉懋中, 章隼. 黃渤海區鳀魚的分布、洄游和探察方法[J]. 水產學報, 1965, 2(2): 27-34.
Ye Maozhong, Zhang Sun. The distribution and behaviour ofEngraulisJaponicustemminck and Schlegel and its detection in the North China Seas (Hwang-Hai and Po-Hai)[J]. Journal of Fisheries of China, 1965, 2(2): 27-34.
[24]張孟海. 渤海南部黃鯽幼魚的季節分布研究[J]. 齊魯漁業, 1994(5): 4-6.
Zhang Menghai. On seasonable distribution of the juvenile ofSetipinnatatyin the southern Bohai Sea[J]. Shandong Fisheries, 1994(5): 4-6.
[25]張孟海, 孫同秋, 李永明, 等. 渤海南部黃鯽季節分布的研究[J]. 齊魯漁業, 1995(4): 10-13.
Zhang Menghai, Sun Tongqiu, Li Yongming, et al. On seasonal distribution ofSetipinnatatyin southern Bohai Sea[J]. Shandong Fisheries, 1995(4): 10-13.
[26]鄧景耀, 金顯仕. 萊州灣及黃河口水域漁業生物多樣性及其保護研究[J]. 動物學研究, 2000, 21(1): 76-82.
Deng Jingyao, Jin Xianshi. Study on fishery biodiversity and its conservation in Laizhou Bay and Yellow River Estuary[J]. Zoological Research, 2000, 21(1): 76-82.
[27]金顯仕, 趙憲勇, 孟田湘, 等. 黃、渤海生物資源與棲息環境[M]. 北京: 科學出版社, 2005.
Jin Xianshi, Zhao Xianyong, Meng Tianxiang, et al. Biological resources and habitat of Yellow and Bohai Seas[M]. Beijing: Science Press, 2005.
[28]唐啟升, 葉懋中. 山東近海漁業資源開發與保護[M]. 北京: 農業出版社, 1990.
Tang Qisheng, Ye Maozhong. The exploitation and conservation of nearshore fisheries resources off Shandong[M]. Beijing: Agriculture Press, 1990.
[29]李峣, 趙憲勇, 張濤, 等. 黃海鳀魚越冬洄游分布及其與物理環境的關系[J]. 海洋水產研究, 2007, 28(2): 104-112.
Li Yao, Zhao Xianyong, Zhang Tao, et al. Wintering migration and distribution of anchovy in the Yellow Sea and its relation to physical environment[J]. Marine Fisheries Research, 2007, 28(2): 104-112.
[30]顧玉荷, 修日晨. 渤海海流概況及其輸沙作用初析[J]. 黃渤海海洋, 1996, 14(1): 1-6.
Gu Yuhe, Xiu Richen. On the current and storm flow in the Bohai Sea and their role in transporting deposited silt of the Yellow River[J]. Journal of Oceanography of Huanghai & Bohai Seas, 1996, 14(1): 1-6.
[31]田家怡. 黃河三角洲附近海域浮游植物多樣性[J]. 海洋環境, 2000, 19(2): 38-42.
Tian Jiayi. Diversity of phytoplankton in the sea near Yellow River Delta[J]. Marine Environmental Science, 2000, 19(2): 38-42.
[32]孫鵬飛. 萊州灣及黃河口水域漁業資源結構特征與漁業生態系統健康評價的初步分析[D]. 上海: 上海海洋大學, 2014.
Sun Pengfei. Preliminary analysis of fishery resource structure and health assessment of fishery ecosystem in the Laizhou Bay and Yellow River Estuary[D]. Shanghai: Shanghai Ocean University, 2014.
[33]趙亮. 渤海浮游植物生態動力學模型研究[D]. 青島: 中國海洋大學, 2002.
Zhao Liang. A modeling study of the phytoplankton dynamic in the Bohai Sea[D]. Qingdao: Ocean University of China, 2002.
[34]李先超. 山東近岸海域(黃海部分)生態環境現狀及演變特征研究[D]. 青島: 中國海洋大學, 2011.
Li Xianchao. The study on the current ecological environment situation and its developmental feather in off-shore area of Shandong Province (focus on Yellow Sea)[D]. Qingdao: Ocean University of China, 2011.
[35]李澤剛. 黃河口附近海區水文要素基本特征[J]. 黃渤海海洋, 2000, 18(3): 20-28.
Li Zegang. Basic featuers of hydrologic elements in the sea area near the Yellow River Estuary[J]. Journal of Oceanography of Huanghai & Bohai Seas, 2000, 18(3): 20-28.
[36]朱元鼎, 張春霖, 成慶泰. 東海魚類志[M]. 北京: 科學出版社, 1963.
Zhu Yuanding, Zhang Chunlin, Cheng Qingtai. The Fishes of the East China Sea[M]. Beijing: Science Press, 1963.
[37]王文海, 夏東興, 高興辰, 等. 中國海灣志·第四分冊·山東半島南部和江蘇省海灣[M]. 北京: 海洋出版社, 1993.
Wang Wenhai, Xia Dongxing, Gao Xingchen, et al. Annals of China’s Bay (book four): Bays in Southern Shandong Peninsula and Jiangsu Province[M]. Beijing: Science Press, 1993.
Spatio-temporal distribution of small-sized fish species in Clupeidae and Engraulidae and its relationships with environmental factors in Huanghe River Estuary and its adjacent waters
Li Min1, Ji Yupeng1, Xu Binduo1, Xue Ying1, Ren Yiping1
(1.CollegeofFisheries,OceanUniversityofChina,Qingdao266003,China)
The small-sized fish species in Clupeidae and Engraulidae are important components in marine ecosystem, as most of them feed on plankton and are food resources for fishes with high trophic levels. Regarding the important ecological roles, their spatiotemporal distribution has an important influence on a variety of organisms. The distribution of this taxon and its relationship with environmental factors was analyzed using generalized additive models (GAM) based on the data collected from the bottom trawl surveys conducted in June, August, October 2013 and February, April, May 2014 in the Huanghe River Estuary and its adjacent waters. The relative abundance of the small-sized fish species in Clupeidae and Engraulidae varied remarkably among seasons, and the relative abundance was higher in summer and autumn and lower in spring and winter. This taxon had significant spatial variation and showed different distribution patterns among months. These species aggregated in the coastal waters of the estuary in April, May and June and moved outward and expanded their distribution in August and October. GAM analysis showed that the spatio-temporal distribution of these species were influenced by several main factors, including sea surface salinity and sea surface temperature, depth and phytoplankton abundance in the Yellow River Estuary. Surface salinity had the most significant effect on the relative abundance of the species in Clupeidae and Engraulidae. The relative abundance was positively corrected with surface salinity and maintained at a high level with surface salinity of 30. The surface water temperature also had a positive effect on the relative abundance of the species in Clupeidae and Engraulidae, and the abundance maintained at a high level when SST was higher than 15℃. The phytoplankton abundance was positively correlated with fish abundance, whereas depth showed a concave-shaped correlation. It is inferred that the distribution of fish species in Clupeidae and Engraulidae could be ascribed to their spawning, feeding behaviors and overwintering migration, driven by the seasonal changes in marine environment and other environmental changes caused by the runoff and sand-washing operation of the Huanghe River.
spatio-temporal distribution; relative abundance; environmental factor; generalized additive model
10.3969/j.issn.0253-4193.2016.10.006
2016-01-21;
2016-05-09。
公益性行業(農業)科研專項經費(201303050);青島海洋科學與技術國家實驗室鰲山科技創新計劃項目(2015ASKJ02)。
李敏(1990—),女,山東省德州市人,博士研究生,主要從事漁業資源與生態學的研究。E-mail:haimi0609@163.com
徐賓鐸,男,主要研究漁業資源與生態學和漁業生物學等。E-mail:bdxu@ouc.edu.cn
S931
A
0253-4193(2016)10-0052-10
李敏,紀毓鵬,徐賓鐸,等. 黃河口及鄰近水域小型鳀鯡魚類數量分布及其與環境因子的關系[J].海洋學報,2016,38(10):52-61,
Li Min, Ji Yupeng, Xu Binduo, et al. Spatio-temporal distribution of small-sized fish species in Clupeidae and Engraulidae and its relationships with environmental factors in Huanghe River Estuary and its adjacent waters[J]. Haiyang Xuebao,2016,38(10):52-61, doi:10.3969/j.issn.0253-4193.2016.10.006
