應用CRISPR/Cas9系統在G401細胞株中敲除p21基因
2016-11-12 03:48:11趙秀娟陳萬標張沛濤張娜楚曉文白向陽楊冰吳旭東王璽
天津醫藥 2016年10期
趙秀娟,陳萬標,張沛濤,張娜,楚曉文,白向陽,楊冰,吳旭東,王璽△
應用CRISPR/Cas9系統在G401細胞株中敲除p21基因
趙秀娟1,陳萬標2,張沛濤1,張娜2,楚曉文3,白向陽4,楊冰1,吳旭東1,王璽1△
目的運用CRISPR/Cas9基因編輯技術,在人惡性橫紋肌樣瘤細胞株G401中敲除p21基因。方法通過反轉錄定量PCR(RT-qPCR)及Western blot檢測各瘤細胞株中p21的表達,針對p21基因作用的功能域,設計了靶向人p21基因第3個外顯子的向導RNA(sgRNA),克隆入lentiCRISPR v2載體。將測序及酶切鑒定正確的重組質粒在293T工具細胞中制備慢病毒顆粒并感染G401細胞,使用嘌呤霉素進行陽性細胞篩選,顯微鏡下挑取單克隆細胞團并繼續培養獲得G401單克隆細胞株。提取單克隆細胞株RNA及蛋白,利用RT-qPCR及Western blot方法檢測細胞株中p21的敲除效果。結果p21在人橫紋肌樣瘤細胞中高表達。成功構建靶向p21基因的lentiCRISPR v2-sgRNA重組慢病毒質粒。與對照組相比,篩選得到的G401亞克隆細胞系中p21蛋白表達缺失。結論針對難轉染的G401細胞,應用CRISPR/Cas9系統成功構建了p21基因敲除的穩定株,為后續深入研究p21在人惡性橫紋肌樣瘤中的作用機制奠定了基礎。
橫紋肌瘤;p21;基因敲除;CRISPR/Cas9;慢病毒
腫瘤的發生發展是由多基因共同參與的復雜過程,涉及多條信號通路。p21蛋白作為熟知的抑癌因子,通常以p53調控依賴的方式使細胞周期發生阻滯,屬于細胞周期蛋白依賴性激酶抑制因子[1]。但后續有研究表明p21也可促進細胞增殖,發揮癌基因作用[2]。因此,p21在腫瘤中的作用及機制仍需進一步研究。CRISPR/Cas9(Clustered Regularly InterspacedShortPalindromicRepeats/CRISPR-associated 9)是基于細菌保護自身免受病毒感染的系統衍生而來,在細菌[3]、斑馬魚[4]、大鼠[5]以及人類細胞[6]中均表現出較強的基因組編輯活性。本實驗是基于在人惡性橫紋肌樣瘤(malignant rhabdoid tumor,MRT)細胞中觀察到的p21的表達,利用CRISPR/Cas9基因編輯系統中適用于難轉染的細胞的慢病毒載體質粒,在感染靶細胞的同時產生向導RNA(sgRNA)靶向p21基因,并表達招募Cas9核酸酶對其進行切割,導致p21基因的敲除。G401細胞p21基因敲除穩定株的構建,為更加深入地研究p21在相關腫瘤中的作用機制奠定了基礎。
1 材料與方法
1.1 實驗材料
1.1.1 質粒、菌株及細胞質粒lentiCRISPR v2購自addgene公司。質粒圖譜可于http://www.addgene.org網站查詢。感受態細菌Stbl3購自北京全式金生物技術有限公司。人胚腎細胞株293T及人惡性橫紋肌樣瘤細胞株A204、G401、TTC642及BT16為本實驗室存留。
1.1.2 酶類及主要試劑Fast Digest BsmBI、FastAP、10× FastDigest Buffer(Green)購自Fermentas公司;T4 PNK、T4 ligase、10×T4 Ligation Buffer、10 mmol/L ATP購自NEB公司;Plasmid Safe exonuclease、10×Plasmid Safe Buffer購自Epicentre公司;AxyPrep DNA凝膠回收試劑盒購自愛思進生物技術(杭州)有限公司;質粒大量提取試劑盒購自QIAGEN公司;DMEM高糖培養基、血清及雙抗購自Gibco公司;兔源p21抗體,鼠源β-actin抗體及辣根過氧化物酶標抗鼠IgG、抗兔IgG購自CST公司;2xSYBR Green qPCR Mix購自Roche公司。
1.1.3 其他基因測序工作由華大科技公司完成。sgRNA寡鏈核苷酸(oligo)DNA序列由蘇州金唯智生物科技有限公司合成。
1.2 方法
1.2.1 sgRNA oligo序列的設計利用http://www.ncbi.nlm. nih.gov/網站確定針對p21 CDs區基因序列,sgRNA序列設計參考http://crispr.mit.edu/網站。設計原則如下:(1)首選第1個外顯子進行序列分析設計靶點,若exon1<100 bp,則順延選擇合適大小的外顯子。(2)根據網站分析反饋的結果,選擇分數較高的一對序列,去掉PAM序列堿基。(3)sgRNA的5′端第1個堿基若不是G,則需要在前面補G。(4)以sgRNA序列為模版,設計出其互補鏈,在其兩端加上酶切位點。即在每條sgRNA序列F鏈的5′端添加CACC,R鏈的5′端添加AAAC。以下將lentiCRISPRv2重組質粒簡稱為v2-p21。
1.2.2 lentiCRISPR v2-p21的構建與鑒定lentiCRISPR v2為含U6啟動子的sgRNA骨架慢病毒表達載體,既表達具有Cas9切口酶,又帶有氨芐青霉素和嘌呤霉素抗性。(1)lentiCRISPR v2載體線性化:用Fast Digest BsmBI對v2進行酶切,DNA凝膠電泳后回收線性化的載體。(2)sgRNA的合成及形成二聚體:用T4 PNK對oligo進行磷酸化并使用梯度退火形成二聚體。(3)sgRNA二聚體與v2線性載體連接:v2載體和退火產物比例為1∶3,室溫(25℃)反應60 min。(4)重組質粒純化:使用質粒保護的核酸外切酶去除非特異連接。(5)轉化感受態細菌Stbl3,在氨芐抗性的LB平板上篩選克隆。挑取陽性克隆搖菌,送測序。(6)酶切驗證:使用Fast Digest BsmBI對v2質粒載體、v2-p21進行酶切。酶切后,使用1%的瓊脂糖凝膠電泳對酶切產物進行鑒定。測序正確的克隆提取重組質粒v2-p21。
1.2.3 病毒包裝、細胞感染及單克隆細胞的獲得培養293T工具細胞,待細胞匯合度達到90%左右進行磷酸鈣轉染,轉染前2 h換新鮮的含10%滅活血清的DMEM高糖培養基,配置磷酸鈣轉染復合物:管A中為CaCl2和DNA的混合液,其中vector∶VSVG∶psPAX2為10∶3.5∶6.5,余體積用水補足;管B中為2×HBS,與管A體積相等。將B中液體逐滴加入A中并不斷混勻,室溫放置10 min后,將轉染復合物緩慢均勻加入293T細胞中。轉染16 h內給293T細胞換新鮮培養基,48 h后開始收集含病毒的培養基,連續收集2次。感染前24 h將G401細胞接種至6 cm培養皿中,感染時將收集的病毒上清(含polybrene 8 mg/L)加入到靶細胞中,感染8~12 h后更換新鮮培養基,感染48 h后加入嘌呤霉素篩選(2 mg/L)。篩選48 h后消化細胞,計數400個單細胞接種于10 cm培養皿中,待生長1周左右獲得單細胞集落,在顯微鏡下挑取細胞團接種至24孔板繼續培養。
1.2.4 Western blot及反轉錄定量PCR(RT-qPCR)檢測p21的表達培養人MRT細胞株及對照細胞293T,收集各株細胞,一部分用RIPA裂解細胞獲得全細胞裂解液,離心取上清作為樣品。測定樣品蛋白濃度,取等量蛋白樣品進行SDSPAGE電泳,經半干轉印至NC膜后,5%脫脂牛奶封閉,一抗、二抗孵育,加入曝光底物進行曝光,以β-actin為內參。收集的另一部分細胞用Trizol法提取細胞總RNA,去除基因組DNA,再逆轉成cDNA。設計擴增p21基因的qPCR引物,進行RT-qPCR擴增檢測p21在細胞中的表達,β-actin作為內參,內參與目的基因的退火溫度均為60℃。同樣,野生型G401細胞和測序正確的單克隆細胞被收集用于Western blot及RT-qPCR擴增,進而檢測p21在細胞中的表達情況,比較野生型和p21基因敲除細胞中p21的表達差異。具體操作同上。
1.3 統計學方法采用SPSS 19.0統計軟件進行分析,計量資料采用均數±標準差表示,多組間均數比較采用方差分析,組間多重比較采用LSD-t法,兩個樣本的組間比較采用獨立樣本的t檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 p21在人MRT細胞中的表達RT-qPCR結果顯示,p21的mRNA表達水平在對照及瘤細胞株中的差異有統計學意義(n=6,F=747.367,P<0.01)。4株人惡性橫紋肌樣瘤細胞中p21的mRNA表達水平(分別為2.776±0.135、5.153±0.126、4.866±0.150、5.966±0.330)均高于對照細胞(0.993±0.015,均P<0.01),見圖1A。Western blot結果顯示,p21蛋白在對照細胞和腫瘤細胞中表達變化與mRNA水平一致,見圖1B。
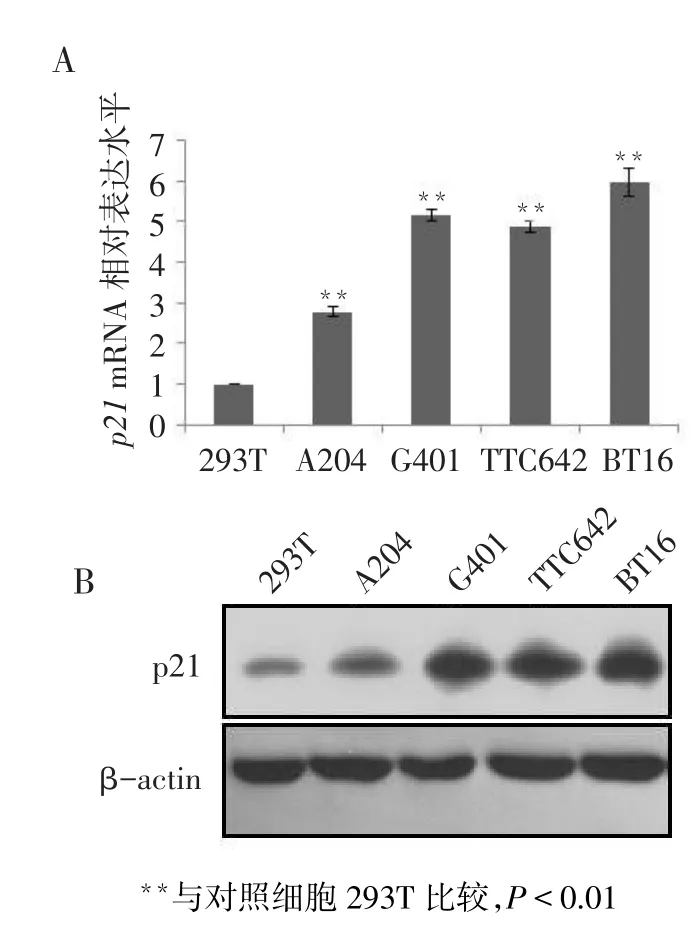
Fig.1The expression of p21 in MRT cell lines圖1 p21在MRT細胞株中的表達
2.2 sgRNA靶點的選擇及寡核苷酸序列人p21基因前2個外顯子區太短,因此選取此基因的第3個外顯子區進行sgRNA設計,挑選脫靶效應最低的sgRNA合成,單鏈DNA模板序列見表1。

Tab.1The sequences of hp21 sgRNA oligo and RT-qPCR primer表1 hp21 sgRNA及RT-qPCR引物序列
2.3 重組質粒lentiCRISPR v2-p21的測序及酶切酶切鑒定結果見圖2A,成功構建的質粒不含有BsmBI酶切位點,因此不能被切割為線性。測序結果見圖2B,插入序列的位置、方向均正確,重組質粒構建成功。
2.4 Western blot及RT-qPCR檢測p21基因敲除的效果同G401野生型細胞相比,G401-p21-KO單克隆細胞中未檢測到p21蛋白表達,表明G401細胞p21基因敲除穩定株構建成功,見圖3A。相比于野生型G401細胞,G401-p21-KO單克隆細胞中p21 mRNA表達降低(n=6,1.000±0.020 vs.0.157± 0.010,t=63.234,P<0.01),見圖3B。
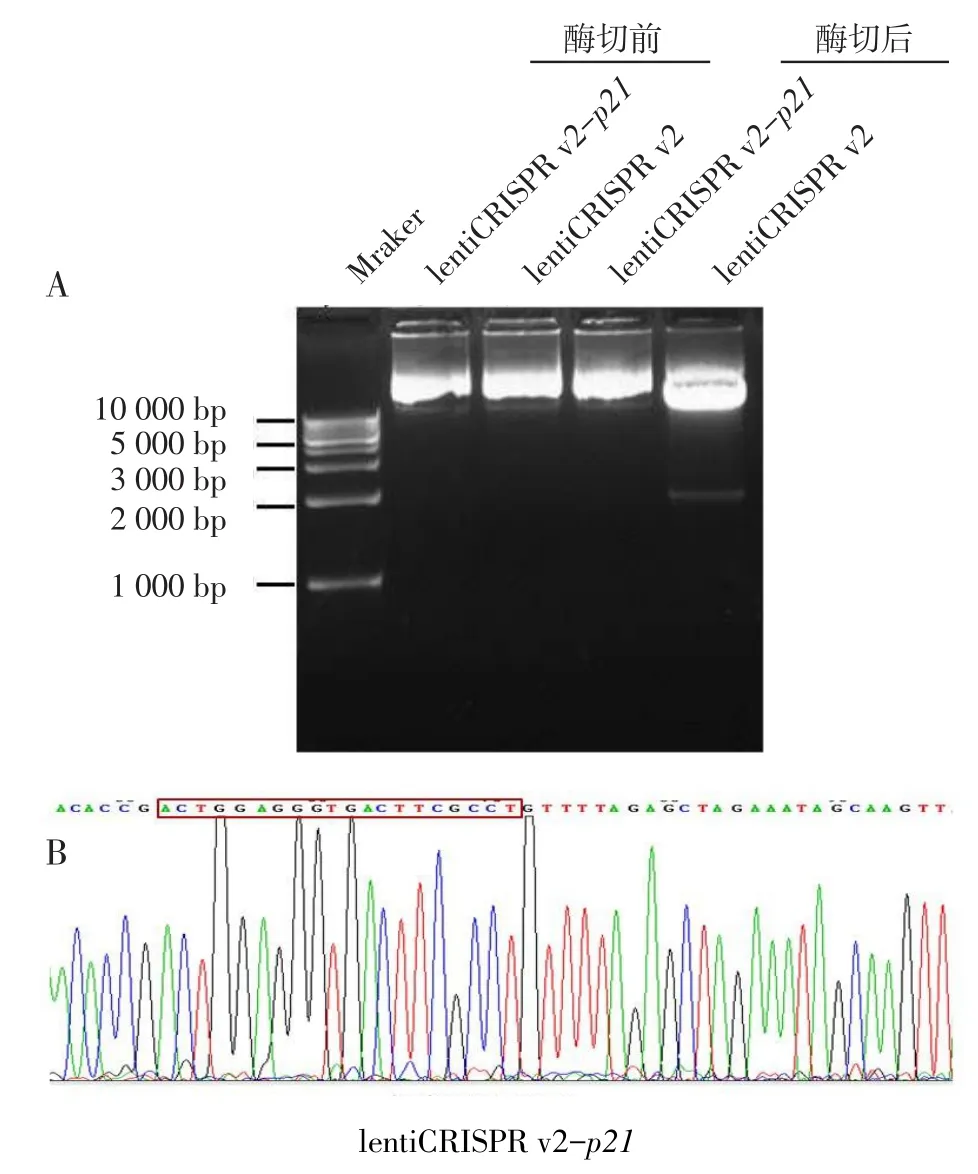
Fig.2The enzyme digestion and sequencing of recombinant eukaryotic expressional plasmid圖2 重組真核表達質粒的酶切和測序結果
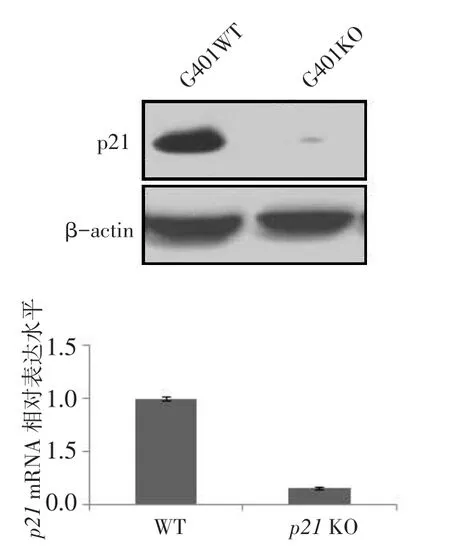
Fig.3Protein and mRNA expression analysis of p21 KO圖3 p21敲除后蛋白及mRNA表達水平結果
3 討論
3.1 p21在腫瘤中的雙重作用自從1993年首次發現p21作為抑癌基因p53的抑癌活性的介導者以來[7],越來越多的文獻報道了p21在腫瘤中的相關作用。首先利用Cdkn1a敲除致瘤的小鼠模型,Martin-Caballero等[8]研究表明細胞周期調控子p21發揮抑癌作用,隨后系列研究報道了多種腫瘤包括結腸癌[9]、宮頸癌[10]、肺癌[11]等均與p21的低表達有關。進一步研究發現在ras基因突變或暴露于致癌物的小鼠聯合p21基因突變會導致腫瘤的發生[12],表明p21可能不是經典的抑癌基因。與之相反,p21被發現在多種情況下還具有癌基因的作用。研究發現p21不僅過表達于各種人類腫瘤如膀胱癌、乳腺癌等,還隨腫瘤的發生發展表達上調,與患者的預后密切相關[13]。具有雙重角色的p21蛋白在腫瘤進展的研究中可能作為新范本,即其作用不是永恒的,而是由其所處的特殊細胞內環境決定的。近年來,有研究者發現p21的這種功能多樣性與其在細胞中的定位相關,即p21表達在胞漿具有抗凋亡作用,而抑制細胞增殖與促進凋亡與其核定位有關[14]。MRT是一種好發于嬰幼兒和兒童的比較少見但高度惡性的腫瘤,大多發生于腎、軟組織及中樞神經系統[15]。本研究顯示p21高表達于MRT多種細胞株中,提示p21在其中可能發揮重要的作用。目前關于p21在MRT中的作用及機制仍不清楚,需進一步研究。
3.2 CRISPR/Cas9慢病毒系統本實驗利用CRISPR/Cas9系統來完成對G401細胞中p21基因的靶向性敲除,包括選擇一個靶區域、構建慢病毒載體、優化轉染等。CRISPR/Cas9系統是新一代的基因定向編輯技術,相比于鋅指核酸內切酶(zinc finger endonuclease,ZFN)[16]和類轉錄激活因子效應物核酸酶(transcription activator-like effector nuclease,TALEN)[17]等經典的基因打靶技術,具有操作簡單、制備周期短、成本低、可同時沉默任意數量的單個基因等優點[18]。此外,CRISPR/Cas9技術與TALEN和ZFN技術相比在降低脫靶效應方面也具有一定的優越性。在CRISPR/Cas9系統中,通過設計及預測出具有最小脫靶率的特異sgRNA,將其導入細胞,對目的基因進行靶標并招募Cas9蛋白進行切割,導致細胞通過非同源性末端接合(nonhomologousendjoining,NHEJ)和同源重組(homology directedrepair,HDR)機制進行DNA修復。在修復過程中斷裂的位點處會發生插入或缺失部分片段,產生框移突變,從而達到敲除目的基因的作用[19]。由于CRISPR/Cas9技術的高效實用性,其在各種細胞模型中被廣泛應用。將目的序列連接到載體并轉染至靶細胞是獲得長期穩定表達細胞株的首要步驟。目前,轉染目的細胞常用的基因導入載體主要包括質粒載體和病毒載體。對于轉染效率較高的細胞通常選擇質粒載體便可達到理想效果[20]。與質粒載體相比,病毒載體具有轉染率高、目的基因長期穩定表達及適用范圍廣等優勢。其中慢病毒載體系統是目前基礎和臨床應用研究中使用最廣泛的一類載體,尤其適用于干細胞、原代細胞及難轉染的細胞株。本實驗使用lentiCRISPR v2質粒作為載體,該質粒不僅可以同時表達Cas9和插入的sgRNA,簡化了表達Cas9質粒和sgRNA質粒共轉染的步驟,而且還含有嘌呤霉素(puromysin)抗性篩選標記,轉染后通過藥物篩選得到陽性克隆。最重要的是其具有慢病毒載體骨架,能夠通過病毒包裝產生慢病毒顆粒導入低轉染率的目的細胞。CRISPR/Cas9介導的基因組編輯技術是目前成功廣泛應用的熱門技術,其在簡易克隆和多重基因組編輯方面有著很大優勢,為利用基因療法治療疾病提供了新方法,在醫學領域具有廣闊的發展前景。
本研究構建的p21基因穩定敲除細胞株,為后續研究p21在人MRT發生發展中的作用及機制提供了工具,也為制定相應的治療策略、提高患者預后奠定了基礎。
[1]Deng C,Zhang P,Harper JW,et al.Mice lacking p21CIP1/WAF1 undergo normal development,but are defective in G1 checkpoint control[J].Cell,1995,82(4):675-684.
[2]Abbas T,Dutta A.p21 in cancer:intricate networks and multiple activities[J].Nat Rev Cancer,2009,9(6):400-414.doi:10.1038/ nrc2657.
[3]Wiedenheft B,Sternberg SH,Doudna JA.RNA-guided genetic silencing systems in bacteria and archaea[J].Nature,2012,482(7385):331-338.doi:10.1038/nature10886.
[4]Chang N,Sun C,Gao L,et al.Genome editing with RNA guided Cas9 nuclease in zebrafish embryos[J].Cell Res,2013,23(4): 465-472.doi:10.1038/cr.2013.45.
[5]Ma Y,Zhang X,Shen B,et al.Generating rats with conditional alleles using CRISPR/Cas9[J].Cell Res,2014,24(1):122-125. doi:10.1038/cr.2013.157.
[6]Ramalingam S,Annaluru N,Chandrasegaran S.A CRISPR way to engineer the human genome[J].Genome Biol,2013,14(2):107. doi:10.1186/gb-2013-14-2-107.
[7]el-Deiry WS,Tokino T,Velculescu VE,et al.WAF1,a potential mediator of p53 tumor suppression[J].Cell,1993,75(4):817-825.
[8]Martin-Caballero J,Flores JM,Garcia-Palencia P,et al.Tumor susceptibility of p21(Waf1/Cip1)-deficient mice[J].Cancer Res,2001,61(16):6234-6238.
[9]Bukholm IK,Nesland JM.Protein expression of p53,p21(WAF1/ CIP1),bcl-2,Bax,cyclin D1 and pRb in human colon carcinomas[J].Virchows Arch,2000,436(3):224-228.
[10]Kim YT,Zhao M.Aberrant cell cycle regulation in cervical carcinoma[J].Yonsei Med J,2005,46(5):597-613.doi:10.3349/ ymj.2005.46.5.597.
[11]Huang CL,Yokomise H,Miyatake A.Clinical significance of the p53 pathway and associated gene therapy in non-small cell lung cancers[J].Future Oncol,2007,3(1):83-89.doi:10.2217/ 14796694.3.1.83.
[12]Gartel AL.Is p21 an oncogene?[J].Mol Cancer Ther,2006,5(6): 1385-1386.doi:10.1158/1535-7163.
[13]Gartel AL.p21WAF1/CIP1and cancer:A shifting paradigm?[J]. BioFactors,2009,35(2):161-164.doi:10.1002/biof.26.
[14]Ohkoshi S,Yano M,Matsuda Y.Oncogenic role of p21 in hepatocarcinogenesis suggests a new treatment strategy[J].World J Gastroenterol,2015,21(42):12150-12156.doi:10.1038/ nrc2657.
[15]Biegel JA,Kalpana G,Knudsen ES,et al.The role of INI1 and the SWI/SNF complex in the development of rhabdoid tumors:meeting summary from the workshop on childhood atypical teratoid/rhabdoid tumors[J].Cancer Res,2002,62(1):323-328.
[16]Sander JD,Dahlborg EJ,Goodwin MJ,et al.Selection-free zincfinger-nucleaseengineeringbycontextdependentassembly(CoDA)[J].Nat Methods,2011,8(1):67-69.doi:10.1038/ nmeth.1542.
[17]Sanjana NE,Cong L,Zhou Y,et al.A transcription activator-like effector toolbox for genome engineering[J].Nat Protoc,2012,7(1): 171-192.doi:10.1038/nprot.2011.431.
[18]Yu Z,Ren M,Wang Z,et al.Highly efficient genome modifications mediated by CRISPR/Cas9 in Drosophila[J].Genetics,2013,195(1):289-291.doi:10.1534/genetics.113.153825.
[19]Nishimasu H,Ran FA,Hsu PD,et al.Crystal structure of Cas9 in complex with guide RNA and target DNA[J].Cell,2014,156(5): 935-949.doi:10.1016/j.cell.2014.02.001.
[20]Fang JP,Zhao XJ,Qi Y,et al.Construction of Asxl2 gene knock out stable NIH3T3 cell line with CRISPR/Cas9n system[J].Tianjin Med J,2015,43(10):1104-1107.[方佳萍,趙秀娟,齊艷,等.利用CRISPR/Cas9n系統構建Asxl2基因敲除的NIH3T3穩定細胞系[J].天津醫藥,2015,43(10):1104-1107].doi:10.11958/j. issn.0253-9896.2015.10.005.
(2016-08-02收稿2016-08-24修回)
(本文編輯李鵬)
Study on p21 gene knock out in G401 cell line by using CRISPR/Cas9 system
ZHAO Xiujuan1,CHEN Wanbiao2,ZHANG Peitao1,ZHANG Na2,CHU Xiaowen3,
BAI Xiangyang4,YANG Bing1,WU Xudong1,WANG Xi1△1 Department of Cell Biology,School of Basic Medical Sciences,2 School of Biomedical Engineering,Tianjin Medical University,Tianjin 300070,China;3 The 66350 Military Medical Team,Lingyun Street,Lianchi District,Baoding City,Hebei Province;4 Dealiner Bioelectronics LLC,Jiangyin△
ObjectiveTo knock out p21 gene in human malignant rhabdoid tumor(MRT)cell line G401 by using CRISPR/Cas9 genome engineering technology.MethodsThe expression of p21 was detected by reverse transcription quantitative PCR(RT-qPCR)and Western blot assay in several MRT cell lines.The guide RNA was designed by targeting the third exon of p21 gene,which encoded its home domains,and then subcloned into lentiCRISPR v2 vector and validated sequencing.The validated plasmids were further used to package and produce the lentivirus in 293T cells,and the G401 cells were infected,then puromycin was used to screen positive cells,and the clusters of G401 monoclonal cells,were obtained by selecting monoclonal cells and culturing under the microscope.The RNA and protein of new clonal cell line were extracted,and RT-qPCR and Western blot assay were applied to confirm whether p21 was successfully knocked out. ResultsThe p21 was highly expressed in MRT tumor cells.The CRISPR/Cas9 lentivirus plasmids,targeted p21 gene were successfully constructed.Compared with negative control group,the expression of p21 was not detected in G401 monoclonal cells,which were successfully screened.ConclusionIn view of the difficult transfection of cells such as G401,p21 knockout stable cell line has been successfully constructed by using CRISPR/Cas9 system,which lays the foundation for further study of the mechanism of p21 in MRT tumors.
rhabdomyoma;p21;gene knockout;CRISPR/Cas9;lentivirus
R73,R349.64
A
10.11958/20160731
天津市高等學校科技發展基金計劃項目(093-201301);天津市自然科學基金青年項目(16JCQNJC10300,16JCQNJC12100);國家自然科學基金資助項目(81500170)
1天津醫科大學基礎醫學院細胞生物學系(郵編300070),2生物醫學工程與技術學院;3河北省保定市蓮池區凌云街66350部隊衛生隊;4江陰迪林生物電子技術有限公司
趙秀娟(1983),女,博士,講師,主要從事腫瘤的表觀遺傳學研究
△通訊作者E-mail:wangxi@tmu.edu.cn
