植物多酚通過Nrf2/ARE信號通路抗氧化作用研究進展
2016-11-14 08:39:03鄭少杰盧可可肖星凝吳素蕊
食品科學 2016年7期
廖 霞,鄭少杰,盧可可,肖星凝,吳素蕊,明 建,3,
(1.西南大學食品科學學院,重慶 400715;2.中華全國供銷合作總社昆明食用菌研究所,云南 昆明 650223;3.重慶市特色食品工程技術研究中心,重慶 400715)
植物多酚通過Nrf2/ARE信號通路抗氧化作用研究進展
廖 霞1,鄭少杰1,盧可可1,肖星凝1,吳素蕊2,明 建1,3,*
(1.西南大學食品科學學院,重慶 400715;2.中華全國供銷合作總社昆明食用菌研究所,云南 昆明 650223;3.重慶市特色食品工程技術研究中心,重慶 400715)
植物多酚是植物體內重要的次生代謝產物,是重要的天然抗氧化劑,能夠清除自由基和淬滅活性氧。Nrf2(NF-E2-related factor 2)/抗氧化反應元件(antioxidant response element,ARE)信號通路是增強機體抗氧化功能最重要的保護性信號途徑,在細胞抵御氧化應激機制中有著重要的地位,是抗氧化研究領域的熱點。本文綜述了Nrf2/ARE信號通路的組成及其調節機制,總結植物多酚通過該信號通路增強機體抗氧化應激能力,減少細胞凋亡、參與神經保護、延緩衰老和減少氧化損傷等的能力,以期為植物多酚應用于抗氧化保健食品和藥品的開發提供一定依據。
植物多酚;Nrf2/ARE;信號通路;抗氧化活性
氧化應激產生的活性氧(reactive oxygen species,ROS)直接或間接地損傷細胞內蛋白質、脂質、核酸等物質的生理功能,是眾多疾病發生的病理生理基礎。植物多酚是植物體內重要的次生代謝產物,其酚羥基中鄰位酚羥基極易被氧化,且對活性氧等自由基有較強捕捉能力,使植物多酚能夠清除自由基和淬滅活性氧[1]。許多研究發現植物多酚可用于治療與氧化應激有關的各種慢性疾病,如腫瘤、炎癥、糖尿病、高脂血癥等,是最理想的天然抗氧化劑來源[2]。
Nrf2(NF-E2-related factor 2)/抗氧化反應元件(antioxidant response element,ARE)信號通路是機體抵抗外界氧化和化學等刺激的防御性轉導通路[3]。Nrf2是細胞氧化應激反應的關鍵因子,受Keap1(Kelch-like ECH-associated protein-1)調控,Nrf2通過與ARE相互作用調節抗氧化蛋白和Ⅱ相解毒酶的表達,Nrf2/ARE被認為是機體最重要的內源性抗氧化信號通路[4]。許多天然植物多酚可以通過誘導抗氧化酶、蛋白表達或激活Nrf2/ARE信號通路,減輕細胞氧化損傷[5]。
1 Nrf2/ARE 信號通路
Nrf2/ARE信號通路就是Nrf2增強機體抗氧化功能的信號途徑,在細胞抵御外源性或內源性氧化應激的機制中具有重要地位。Nrf2在應答氧化應激中,是調節抗氧化和細胞保護基因的關鍵因子,其活性主要受Keapl調控,在抗氧化物質的誘導下,通路激活,Keap1構象改變,蛋白酶體降解,Nrf2釋放,發生核轉位,與抗氧化反應元件ARE結合,促進編碼細胞Ⅱ相解毒酶基因,從而保護細胞[6-7]。
1.1 Nrf2/ARE信號通路的結構和功能
Nrf2作為抗氧化劑轉錄的主要調節者[8],屬于CNC(cap’-n’-collar)轉錄因子家族成員,是一個由2.2 kb堿基對編碼的蛋白(66 kD),是活力最強的轉錄因子[9]。Nrf2含有7 個功能領域(Neh1~Neh7),Nehl區含有1 個C端亮氨酸拉鏈結構——bZip,與細胞核內小Maf蛋白形成異二聚體,使Nrf2能夠識別、結合ARE,從而啟動目標基因轉錄。Neh2區是Nrf2與胞漿蛋白Keap1結合區,含有ETGE基序、DLG基序2 個結合位點,對Nrf2活性進行負調節;Neh3是Nrf2的轉錄活性必不可少的;Neh4和Neh5是參與啟動下游基因轉錄的結構域,需與共激活因子cAMP應答元件結合蛋白(cAMP responsive element binding protein,CREB)轉錄激活劑結合后,才能啟動轉錄過程;Neh6與Nrf2的負調節有關;第3個負調節領域是Neh7,通過與RXRα相互作用進行調節[10-11]。
Keap1是氧化應激調節靶基因表達的感受器,是控制Nrf2信號通路的分子開關。Keap1屬于Kelch家族多區域阻遏蛋白(69 kD)[12]。在胞漿中錨定于肌動蛋白細胞骨架上,是Nrf2的胞漿蛋白伴侶分子,與Nrf2結合并且抑制其活性。Keap1包含5 個區:N端結構域(N-terminal rerminal,NTR)、大復合體形成序列區(broadcomplex, tramtrack,and bric-a-brac,BTB)、插入區(intervening region,IVR)、雙甘氨酸重復區(bouble glycine repeat,DGR)、C端結構域(the C-terminal region,CTR)。BTB區介導Keap1二聚體化并與Nrf2-Keap1結合力有關,是阻止Ⅱ相解毒酶基因轉錄所必需的,與Nrf2的激活有關;IVR區富含半胱氨酸殘基,其對活性親電物質敏感[13],是整個蛋白的功能調節區,與Nrf2的穩定性有關;DGR區是Keap1與Nrf2的結合區,也是Keap1與胞漿內肌動蛋白結合的位點,是Keap1發揮抑制作用的關鍵區域[14]。Keap1與Nrf2結構圖見圖1。
ARE是機體內重要的保護性順式應答元件,ARE的核心序列是5’-TGAC nnnGC-3’[15],是抗氧化蛋白和許多Ⅱ相解毒酶基因上游的一段轉錄調控因子,存在于大部分細胞保護基因的近端啟動子區域[16],Nrf2通過ARE來調節體內多種細胞防御因子,包括細胞內調節氧化還原平衡的蛋白、Ⅱ相解毒酶、轉運蛋白等,在抗氧化作用中扮演重要角色。

圖1 Keap1(a)和Nrf2(b)結構示意圖[10-14]Fig.1 Structure diagram of Keap1 (a) and Nrf2 (b)[10-14]
1.2 Nrf2/ARE信號通路抗氧化調節機制
在正常生理狀態下,絕大部分細胞核轉錄因子Nrf2以非活性狀態儲存于細胞質中,Keapl的DGR與Nrf2的Neh2部位(包括DLG和ETGE序列[17])相互作用結合成二聚體,與胞漿肌動蛋白結合被錨定在胞漿中,是E3泛素連接酶的適配底物,促進Nrf2泛素結合和隨后的蛋白酶體降解,使Nrf2的含量維持在基礎水平[18];少部分Nrf2以活性狀態存在于細胞核中介導相關基因的基本轉錄。
2.風險評估。風險評估是內部控制的首要前提,是在風險識別和預測的基礎上,采用定性或定量方法,對風險發生可能性和影響程度進行預計和估算,最終確定風險評級的過程。具體內容包括風險管理組織體系、風險識別和分類、風險評估、風險應對及風險監控與報告等。企業應全面梳理、識別和評估影響目標實現的相關風險,建立本企業風險清單或風險庫。針對不同風險,結合本企業的風險偏好和承受度,權衡利弊,按照成本效益原則制定完善相應的規避、降低、分擔或承受措施。通常情況下,內部風險可以進行有效防范和控制,外部風險主要采取規避。
現今比較公認的Keap1-Nrf2蛋白相互作用模型為“hinge and latch”模型[19]:發生氧化應激時,Keap1的半胱氨酸殘基被修飾,Keap1構象發生改變,“門閂”DLG序列松了。雖然“鉸鏈”ETGE序列仍然連接著Keap1,但是門閂連接的破壞足夠阻止Nrf2的泛素化,Nrf2發生磷酸化,26S蛋白酶體對Nrf2識別減弱,導致Nrf2釋放出來,Nrf2累積[20]。Nrf2被活化后轉移入核,與小分子蛋白Maf(如MafG、MafT、MafF、MafK[21])、Jun、ATF(activating transcription factor)[22]、Bach1[23]等結合形成異二聚體,識別ARE上的DNA序列并與之結合,激活ARE調節的靶基因啟動Ⅱ相解毒酶和抗氧化應激蛋白基因(醌氧化還原酶(nicotinamide quinone oxidoreductase 1,NQO1)、谷胱甘肽硫轉移酶(glutathione S-transferase,GSTs))的轉錄,編碼抗氧化蛋白,提高細胞抗氧化應激能力。另外,多種蛋白激酶,例如絲裂原活化蛋白激酶(mitogen activated protein kinases,MAPKs)、蛋白激酶C(protein kinase C,PKC)、磷脂酰肌醇-3激酶(phosphatidyl inositol 3-kinase,PI3K)、細胞外信號調節激酶(extracellular signal-regulated kinase,ERK)、蛋白激酶R樣內質網激酶(protein kinase R-like endoplasmic reticulum kinase,PERK)等,可通過誘導Nrf2的磷酸化參與對Nrf2轉錄活性的調節。
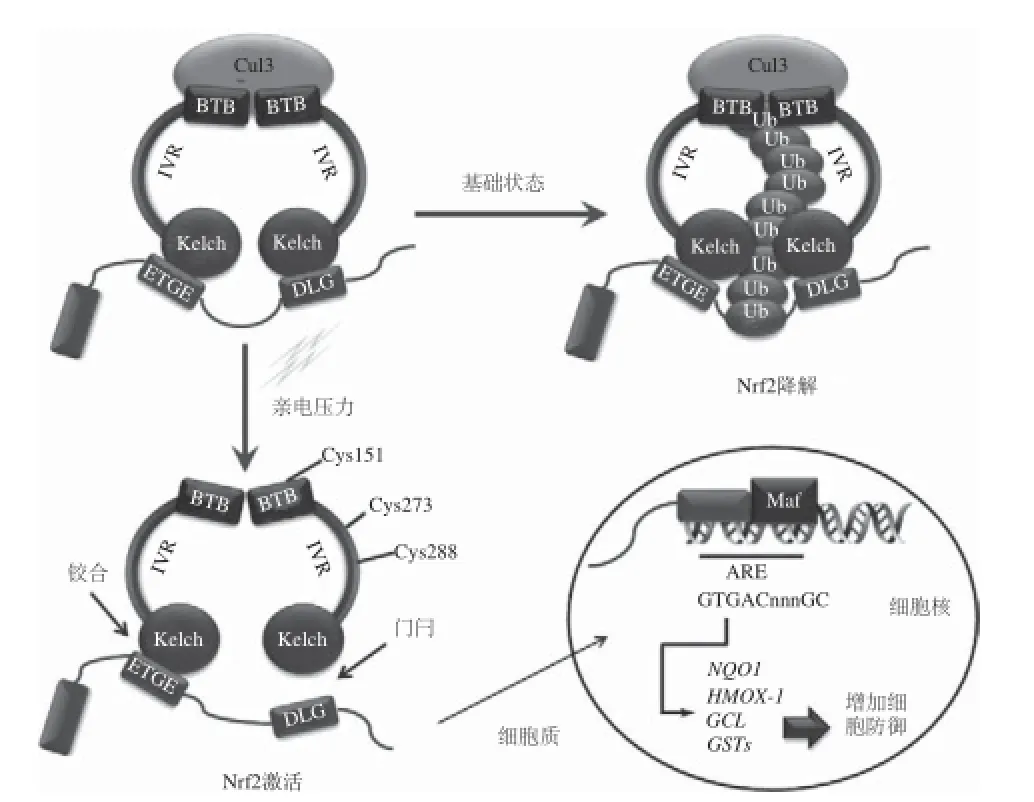
圖2 Nrf2/ARE信號通路抗氧化調節機制[6,24]Fig.2 Regulating mechanism of oxidation resistance by Nrf2/ARE signaling pathway[6,24]
當氧化還原平衡恢復,Nrf2與ARE序列解離,輸出到胞漿,通過支架蛋白Cullin3依賴的E3泛素連接酶機制進行泛素化后降解,關閉Nrf2通路,重新維持低水平的Nrf2[25]。解離后的Nrf2要與其他蛋白(染色質重構蛋白CHD6、CREB結合蛋白/CBP,等)結合才能最終完成對靶基因轉錄的調控。
1.3 Nrf2/ARE信號通路與抗氧化酶關聯
Nrf2是調節細胞抗氧化應激的重要轉錄因子,在生理情況下,胞漿蛋白伴侶分子Keap1與Nrf2結合使其處于抑制狀態,在外界氧化應激源影響下,Nrf2與Keap1解偶聯后轉移入核內,與抗氧化反應元件結合,啟動其調控的Ⅱ相解毒酶(超氧化物岐化酶(superoxide dismutase,SOD)、一氧化氮合成酶(nitric oxide synthase,NOS)、過氧化氫酶(catalase,CAT)、谷胱甘肽過氧化物酶(glutathione peroxidase,GSH-Px)等)[26]和抗氧化酶的基因(HO-1、γ-GCS、SOD、NQO1等)[27]、抗氧化蛋白(bcl-2、Ferritin等[28])的表達,增加細胞對氧化應激的抵抗性從而發揮保護作用。
血紅素氧化合酶1(heme oxygenase-1,HO-1)是血紅素降解的限速酶,在氧化應激條件下,通過激活Nrf2通路,調控HO-1基因表達,可以起到保護器官、神經、人黑色素細胞等的作用[29],Qaisiya等[30]對神經母細胞瘤SH-SY5Y細胞系的實驗表明,Nrf2會誘導膽紅素表達,介導抗氧化應激反應。谷氨酰半胱氨酸合成酶(γ-glutamylcysteine synthetase,γ-GCS)是還原型谷胱甘肽合成的限速酶,是肺內重要的抗氧化酶,通過Nrf2通路,增加γ-GCS的含量和活性,促進谷胱甘肽(glutathione,GSH)的合成,增強組織細胞的抗氧化能力[31]。SOD是抗氧化酶家族重要成員之一,通過Nrf2通路可以上調MnSOD及HO-1蛋白表達,在心肌延遲保護等方面發揮重要作用[32]。NQO1是一種調節細胞內物質處于氧化還原狀態的黃素酶,Cheng Zhenguo等[33]對Nrf2的表達在腦損傷抗氧化保護的研究發現HO-1、NQO1水平在抗氧化應激中發揮著重要作用。Nrf2通路可以通過調控抗氧化蛋白或酶的表達,維持機體氧化還原平衡,從而保護機體或降低損傷。
2 植物多酚通過Nrf2/ARE通路的抗氧化作用
植物多酚作為天然抗氧化物質,這些抗氧化物質會進入細胞膜內,保護機體組織抵御活性氧的攻擊,其抗氧化機制比較復雜。大量研究表明,植物多酚如茶多酚、白藜蘆醇、姜黃素、原花青素、槲皮素等可以通過Nrf2/ARE通路,調節該細胞內信號轉導分子的活性,增強抗氧化基因或蛋白的表達,提高機體抗氧化應激能力,發揮抗氧化作用[34]。
2.1 茶多酚
2.2 白藜蘆醇
白藜蘆醇(resveratrol)是一種多酚類植物抗毒素,具有較強的抗氧化活性,對肝臟、神經系統、血管和細胞損傷有較好的保護作用。白藜蘆醇能夠促進Keap1蛋白降解,激活Keap1-Nrf2-ARE信號通路,啟動抗氧化應激一系列蛋白和酶的表達,增強細胞抗氧化應激的能力[39]。通過Nrf2/keap1調節其下游調控蛋白(SOD、CAT、GSH-Px),可以有效保護腎臟高血糖介導的氧化損傷[40]。白藜蘆醇能降低組織中MDA含量,增加T-SOD活力,上調Nrf2、HO-1基因和其蛋白表達,改善糖尿病小鼠腎臟的氧化應激狀態[41],可以通過調節肝熱休克蛋白和核轉錄因子,如SOD、CAT、GSH Px活性以及Nrf2的表達,減少鵪鶉肝細胞對熱的氧化應激損傷[42]。楊竟[43]研究發現白藜蘆醇可通過抑制小鼠左心室PKC-β2、TGF-βl/Smad2、核因子κB(nuclear factor kappa B,NF-κB)、Nrf2 mRNA表達和激活Nrf2信號通路抗糖尿病和心肌病。
2.3 姜黃素
姜黃素(curcumin)是一種從姜黃屬植物根莖中提取的多酚類物質,能夠有效清除自由基,從而減輕氧化應激過程中產生的損害,姜黃素通過多種機制發揮作用,其中之一是促進細胞的Nrf2信號通路活化。姜黃素能誘導亞慢性鎘暴露大鼠腎臟Nrf2蛋白活化,并進一步誘導Nrf2下游γ-GCS、HO-1及GPx-1等基因表達,拮抗亞慢性鎘染毒所致的大鼠腎臟氧化應激[44]。王欣等[45]研究發現,與單純砷染毒組相比,姜黃素干預組的肝臟Nrf2、HO-1和NQO1蛋白表達均顯著增高,同時肝臟細胞的胞漿和胞核中棕褐色陽性顆粒均顯著增多,且Nrf2入核明顯增多,表明姜黃素能夠促進Nrf2核轉位和活化,并進一步激活Nrf2下游信號通路。此外,姜黃素能抑制MCF-7細胞增殖和誘導Nrf2蛋白的表達,從而顯著抑制FEN1基因的表達,這可能是姜黃素抑制乳腺癌細胞增殖的新機制[46]。另有研究發現,姜黃素能夠增強Nrf2蛋白表達,促進Nrf2核轉位,并激活其下游抗氧化酶基因HO-1和Ⅱ相代謝酶基因γ-GCS的mRNA表達,以及上調HO-1和γ-GCS蛋白表達水平,顯著地提高星形膠質細胞的細胞活力,抑制丙烯腈誘導的ROS生成,作用機制可能與激活Nrf2-ARE通路相關[47]。
2.4 槲皮素
槲皮素(quercetin)及其衍生物是植物界分布最廣的黃酮類化合物,具有良好的抗氧化及清除自由基的作用。槲皮素對H2O2損傷的HepG2細胞具有保護作用,其機制可能與槲皮素通過激活HepG2細胞的轉錄因子Nrf2,上調Ⅱ相酶(SOD、CAT和NQO1)活性和蛋白表達有關[48]。鄭靜等[49]研究發現槲皮素能上調Nrf2、HO-1基因表達,以及上調腦源性神經營養因子(brain derived neurotrophic factor,BDNF)基因表達,減輕腦部氧化應激,改善學習記憶能力。Ramyaa等[50]研究發現槲皮素可以上調Nrf2表達及下調NF-κB和COX-2表達,減輕ROS產生和鈣釋放,調節赭曲霉毒素A(ochratoxin A,OTA)在HepG2細胞中誘導的氧化應激和氧化還原信號。還有研究表明,槲皮素可通過激活c-Jun氨基末端激酶(c-Jun NH2-terminal Kinase,JNK)、p38和PI3K/Akt信號通路發生磷酸化,增強Nrf2與DNA結合活性,誘導金屬硫蛋白的表達,預防肝損傷[51]。
2.5 原花青素
原花青素(grape seed procyanidine,OPC)是由不同數量的兒茶素或表兒茶素縮合而成的一類多酚化合物總稱,已被證實具有抗氧化功能活性。有研究表明,在氧化應激狀態下,葡萄籽原花青素(grape seed procyanidine extract,GSP)通過降低Nrf2、HO-1、NQO1 mRNA的表達量,提高Keapl mRNA表達量以減輕ROS對仔豬肝臟Nrf2信號通路的影響;同時,GSP通過減緩內質網應激相關分子葡萄糖調節蛋白78(glucose-regulated protein 78,GRP78)mRNA表達量,抑制氧化應激誘導的XBP-1、ATF6、PDIA4和CHOP mRNA表達上調,同時降低肝臟caspase-12的含量,減輕內質網氧化損傷[52]。肉桂原花青素(proanthocyanidins of cinnamon,CPAs)通過抑制Nrf2及其調節酶NQO1的表達,可以增強肺癌A549細胞對細胞毒性抗腫瘤藥物的敏感性[53]。蓮子皮低聚原花青素(lotus seeds skin oligomeric proanthocyanidins,PMR)可以有效地激活Nrf2/ARE信號通道,促進Nrf2解離,從而加強具有抗氧化保護基因如HO-1的轉錄[54]。
2.6 其他
除上述植物多酚能通過Nrf2通路調控抗氧化作用外,還存在很多其他植物多酚有類似的作用。比如,染料木黃酮能拮抗Aβ25-35介導的星形膠質細胞氧化損傷作用,主要通過調控Nrf2/ARE通路,上調Nrf2、HO-1、GCLC及MnSOD基因及其蛋白表達水平,從而達到抗氧化保護作用[55]。金絲桃苷可減弱缺氧或H2O2引起的LO2細胞氧化應激損傷,對LO2細胞進行預保護,其機制可能是通過促進Nrf2向細胞核內轉移,與Bach1競爭細胞保護蛋白啟動子區域中的ARE,從而上調細胞保護蛋白的水平來發揮其細胞保護作用[56]。丹皮酚[57]、丹酚酸[58]、安石榴苷[59]、黃芩素[60]等也可通過激活Nrf2/ARE信號通路,而起到抗氧化、抗癌、保護神經作用。
3 結 語
機體內氧化還原水平的失衡是眾多疾病的病理基礎,通過激活Nrf2/ARE信號通路,可以調節體內氧化還原平衡,從而保護機體功能正常,Nrf2/ARE信號通路現已成為疾病預防控制以及治療策略的新靶點,當然,過度的免疫應答和腫瘤細胞需要進一步探討。越來越多的研究表明植物多酚可作用于Nrf2/ARE通路,誘導抗氧化蛋白和Ⅱ相代謝酶的表達,減輕氧化應激損傷,無疑為抗氧化治療提供了新思路。但是由Nrf2/ARE信號通路介導的抗氧化機制還需要更深入的研究,為拓展植物多酚新的研究領域和應用范圍具有重要意義。
[1] FRAGA C G. Plant polyphenols: how to translate their in vitro antioxidant actions to in vivo conditions[J]. IUBMB Life, 2007, 59(4/5): 308-315. DOI:10.1080/15216540701230529.
[2] YOSHIDA T, HATANO T, ITO H. High molecular weight plant poplyphenols (tannins): prospective functions[J]. Recent Advances in Phytochemistry, 2005, 39: 163-190. DOI:10.1016/S0079-9920(05)80008-5.
[3] 蔡維霞, 張軍, 胡大海. 氧化和化學應激的防御性轉導通路: Nrf2/ ARE[J]. 中國生物化學與分子生物學報, 2009, 25(4): 297-303. DOI:10.13865/j.cnki.cjbmb.2009.04.010.
[4] WANG Yingzhen, ZHANG Youcheng, CHENG Junsheng, et al. Protective effects of BML-111 on cerulein-induced acute pancreatitisassociated lung injury via activation of Nrf2/ARE signaling pathway[J]. Inflammation, 2014, 37(4): 1120-1133. DOI:10.1007/ s10753-014-9836-y.
[5] 杜玉. 天然抗氧化劑抗氧化損傷作用及機制[D]. 濟南: 山東大學, 2009: 1-6. DOI:10.7666/d.y1566103.
[6] KANSANEN E, JYRKK?NEN H K, LEVONEN A L. Activation of stress signaling pathways by electrophilic oxidized and nitrated lipids[J]. Free Radical Biology and Medicine, 2012, 52(6): 973-982. DOI:10.1016/j.freeradbiomed.2011.11.038.
[7] LI Li, YANG Na, NIN Ling, et al. Chinese herbal medicine formula Tao Hong Si Wu decoction protects against cerebral ischemiareperfusion injury via PI3K/Akt and the Nrf2 signaling pathway[J]. Journal of Natural Medicines, 2015, 69(1): 76-85. DOI:10.1007/ s11418-014-0865-5.
[8] MILLER C J, GOUNDER S S, KANNAN S, et al. Disruption of Nrf2/ARE signaling impairs antioxidant mechanisms and promotes cell degradation pathways in aged skeletal muscle[J]. Biochimica et Biophysica Acta (Molecular Basis of Disease), 2012(6): 1038-1050. DOI:10.1016/j.bbadis.2012.02.007.
[9] 何金蓮. 五味子經Nrf2信號通路調節肝臟藥物代謝酶及轉運體的研究[D]. 廣州: 廣州中醫藥大學, 2014: 1-12.
[10] 胡敏. Keap1/Nrf2信號通路對再生肝細胞周期的調控[D]. 合肥: 安徽醫科大學, 2014: 9-10.
[11] BAIRD L, SWIFT S, LLèRES D, et al. Monitoring Keap1-Nrf2 interactions in single live cells[J]. Biotechnology Advances, 2014, 32(6): 1133-1144. DOI:10.1016/j.biotechadv.2014.03.004.
[12] 王茜. Nrf2/ARE信號通路在納米二氧化鈦誘導HepG2細胞氧化損傷中的作用研究[D]. 石家莊: 河北醫科大學, 2014: 48-53.
[13] NORDGREN K S, WALLACE K B. Keap1 redox-dependent regulation of doxorubicin-induced oxidative stress response in cardiac myoblasts[J]. Toxicology and Applied Pharmacology, 2013, 274: 107-116. DOI:10.1016/j.taap.2013.10.023.
[14] 劉芳, 杜志銀, 王應雄. Keap1在氧化應激方面的研究進展[J]. 中國臨床藥理學與治療學, 2010, 15(5): 596-600.
[15] 宋亞頎. 轉錄因子NF-E2相關因子2-抗氧化轉錄元件信號路徑細胞保護作用的研究進展[J]. 醫學研究生學報, 2009, 22(4): 431-433. DOI:10.3969/j.issn.1008-8199.2009.04.026.
[16] OHNUMA T, NAKAYAMA S, ANAN E, et al. Activation of the Nrf2/ARE pathway via S-alkylation of cysteine 151 in the chemopreventive agent-sensor Keap1 protein by falcarindiol, a conjugated diacetylene compound[J]. Toxicology and Applied Pharmacology, 2010, 244(1): 27-36. DOI:10.1016/j.taap.2009.12.012.
[17] 王秀君, 吳加國, 唐修文. Nrf2-ARE通路抑制作用研究展望[J]. 浙江大學學報(醫學版), 2010, 39(1): 1-5. DOI:10.3785/j.issn. 1008-9292.2010.01.001.
[18] YANG Ping, KONG Xiangzhe, CHENG Changmei, et al. Synthesis and biological evaluation of 8-substituted and deglucuronidated scutellarin and baicalin analogues as antioxidant responsive element activators[J]. Science China Chemistry, 2011, 54(10): 1565-1575. DOI:10.1007/s11426-011-4361-4.
[19] 姚佳維. PMID激活Nrf2/ARE信號通路的機制及其抗氧化作用研究[D].天津: 天津大學, 2012: 2-10.
[20] STEPKOWSKI T M, KRUSZEWSKI M K. Molecular cross-talk between the NRF2/KEAP1 signaling pathway, autophagy, and apoptosis[J]. Free Radical Biology & Medicine, 2011, 50: 1186-1195. DOI:10.1016/j.freeradbiomed.2011.01.033.
[21] 運晨霞, 吳光, 王坤. Maf家族蛋白的組成及功能[J]. 醫學分子生物學雜志, 2008, 5(1): 82-86. DOI:10.3870/j.issn.1672-8009. 2008.01.019.
[22] YU Miao, LI Hui, LIU Qiongming, et al. Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway[J]. Cellular Signalling, 2011, 23(5): 883-892. DOI:10.1016/j.cellsig.2011.01.014.
[23] 王梅芳, 戴愛國, 胡瑞成. 轉錄因子Nrf2/Bach1及MAPK激酶調控γ-GCS在大鼠慢性阻塞性肺疾病中的作用[J]. 中國病理生理雜志, 2009, 25(5): 959-964. DOI:10.3321/j.issn:1000-4718.2009.05.026.
[24] KASPAR J W, NITURE S K, JAISWAL A K. Nrf2: INrf2 (Keap1) signaling in oxidative stress[J]. Free Radical Biology and Medicine, 2009, 47: 1304-1309. DOI:10.1016/j.freeradbiomed.2009.07.035.
[25] 何茂群, 曹智華, 苗得足. Nrf2信號通路及其在肝組織中的作用[J].藥學進展, 2014, 38(4): 257-263.
[26] 童海達, 王佳茗, 宋英. Keap1-Nrf2-ARE在機體氧化應激損傷中的防御作用[J]. 癌變·畸變·突變, 2013, 25(1): 71-75. DOI:10.3969/ j.issn.1004-616x.2013.01.017.
[27] LI Xianhui, LI Chunyan, XIANG Zhigang, et al. Allicin ameliorates cardiac hypertrophy and fibrosis through enhancing of Nrf2 antioxidant signaling pathways[J]. Cardiovascular Drugs and Therapy, 2012, 26(6): 457-465. DOI:10.1007/s10557-012-6415-z.
[28] 崔俁, 馬海英, 孔力. Nrf2/ARE通路與機體抗氧化機制的研究進展[J].吉林大學學報(醫學版), 2011, 37(1): 187-190. DOI:10.13481/j.1671-587x.2011.01.001.
[29] 喬麗杰, 王延讓, 張明. Nrf2/HO-1通路在氧化損傷保護機制中研究進展[J]. 中國職業醫學, 2012, 40(1): 82-84. DOI:10.11763/ j.issn.2095-2619.2013.01.033.
[30] QAISIYA M, ZABETTA C D C, BELLAROSA C, et al. Bilirubin mediated oxidative stress involves antioxidant response activation via Nrf2 pathway[J]. Cellular Signalling, 2014, 26(3): 512-520. DOI:10.1016/j.cellsig.2013.11.029.
[31] 王梅芳. Nrf2調控抗氧化酶與慢性阻塞性肺疾病[J]. 國際內科學雜志, 2008, 35(2): 120-124.
[32] 黃曉珊. 缺氧預適應通過Nrf2通路上調抗氧化酶介導心肌細胞延遲保護的實驗研究[D]. 南昌: 南昌大學, 2014: 21-27.
[33] CHENG Zhenguo, ZHANG Guodong, SHI Pengqiang, et al. Expression and antioxidation of Nrf2/ARE pathway in traumatic brain injury[J]. Asian Pacific Journal of Tropical Medicine, 2013, 6(4): 305-310. DOI:10.1016/S1995-7645(13)60061-9.
[34] PALL M L, LEVINE S. Nrf2, a master regulator of detoxification and also antioxidant, anti-inflammatory and other cytoprotective mechanisms, is raised by health promoting factors[J]. Acta Physiologica Sinica, 2015, 67(1): 1-18. DOI:10.13294/ j.aps.2015.0001.
[35] 蔣潔琳, 溫旭燁, 胡雅瓊. 表沒食子兒茶素沒食子酸酯(EGCG)對癌癥細胞信號傳導鏈的影響[J]. 食品科學, 2012, 33(9): 319-325.
[36] NA H K, KIM E H, JUNG J H, et al. Epigallocatechin gallate induces Nrf2-mediated antioxidant enzyme expression via activation of PI3K and ERK in human mammary epithelial cells[J]. Archives of Biochemistry and Biophysics, 2008, 476(2): 171-177. DOI:10.1016/ j.abb.2008.04.003.
[37] 孫萬良, 魏麗, 張晶. EGCG對大鼠放射性肺損傷的防治作用及機制探索[J]. 生物技術通訊, 2013, 24(4): 504-509. DOI:10.3969/ j.issn.1009-0002.2013.04.014.
[38] 楊曉云. 茶多酚EGCG在大腸癌預防中的作用及其機制研究[D]. 濟南: 山東大學, 2006: 110-122.
[39] 王前, 閏繼紅, 吳波. 白藜蘆醇促進keap1蛋白降解的可能機制研究[J].中華臨床醫師雜志, 2013, 7(7): 2991-2994. DOI:10.3877/cma. J.issn.1674-0785.2013.07.102.
[40] PALSAMY P, SUBRAMANIAN S. Resveratrol protects diabetic kidney by attenuating hyperglycemia-mediated oxidative stress and renal inflammatory cytokines via Nrf2-Keap1 signaling[J]. Biochimica et Biophysica Acta, 2011, 1812(7): 719-731. DOI:10.1016/ j.bbadis.2011.03.008.
[41] 高絲娜. 白藜蘆醇對糖尿病小鼠腎臟氧化應激損傷的保護作用[D].石家莊: 河北醫科大學, 2013: 34-36.
[42] SAHIN K, ORHAN C, AKDEMIR F, et al. Resveratrol protects quail hepatocytes against heat stress: modulation of the Nrf2 transcription factor and heat shock proteinms[J]. Journal of Animal Physiology and Animal Nutrition, 2012, 96(1): 66-74. DOI:10.1111/j.1439-0396.2010.01123.x.
[43] 楊竟. 白藜蘆醇抗糖尿病心肌病心肌纖維化作用及機制研究[D]. 遵義: 遵義醫學院, 2013: 22-39.
[44] 黎珊珊, 徐兆發, 李靜慧. 姜黃素對鎘致大鼠腎Nrf2及HO-1、γ-GCS、Gpx-1 mRNA和蛋白表達的影響[J]. 毒理學雜志, 2015, 1(29): 31-35. DOI:10.16421/j.cnki.1002-3127.2015.01.006.
[45] 王欣, 董丹丹, 邢曉越. 姜黃素干預對慢性飲水砷暴露小鼠肝臟Nrf2信號通路的影響[J]. 中國地方病防治雜志, 2012, 6(27): 418-421.
[46] CHEN Bin, ZHANG Youzhi, WANG Yang, et al. Curcumin inhibits proliferation of breast cancer cells through Nrf2-mediated downregulation of Fen1 expression[J]. The Journal of Steroid Biochemistry and Molecular Biology, 2014, 143(9): 11-18. DOI:10.1016/ j.jsbmb.2014.01.009.
[47] 趙小武. 姜黃素激活Nrf2-ARE通路保護丙烯睛對大鼠星形膠質細胞毒性的研究[D]. 鎮江: 江蘇大學, 2010: 1-4.
[48] 初亮. 槲皮素對HepG2細胞二相酶的誘導作用[D]. 大連: 大連醫科大學, 2009: 19-30. DOI:10.7666/d.y1557258
[49] 鄭靜, 田英杰, 陳立立. 槲皮素對高脂日糧小鼠學習記憶能力的影響研究[J]. 食品工業科技, 2013, 34(19): 332-336. DOI:10.13386/ j.issn1002-0306.2013.19.003.
[50] RAMYAA P, KRISHNASWAMY R, PADMA V V. Quercetin modulates OTA-induced oxidative stress and redox signaling in HepG2 cells-up regulation of Nrf2 expression and down regulation of NF-κB and COX-2[J]. Biochimica et Biophysica Acta, 2014(1): 681-692. DOI:10.1016/j.bbagen.2013.10.024.
[51] WENG Chiajui, CHEN Meujung, YEH Chitai, et al. Hepatoprotection of quercetin against oxidative stress by induction of metallothionein expression through activating MAPK and PI3K pathways and enhancing Nrf2 DNA-binding activity[J]. New Biotechnology, 2011, 28(6): 767-777. DOI:10.1016/j.nbt.2011.05.003.
[52] 趙嬌. 葡萄籽原花青素緩解氧化應激仔豬肝臟損傷及可能機制研究[D]. 雅安: 四川農業大學, 2013: 20-42.
[53] OHNUMA T, MATSUMOTO T, ITOI A, et al. Enhanced sensitivity of A549 cells to the cytotoxic action of anticancer drugs via suppression of Nrf2 by procyanidins from Cinnamomi Cortex extract[J]. Biochemical and Biophysical Research Communications, 2011, 413(4): 623-629. DOI:10.1016/j.bbrc.2011.09.014.
[54] 李綺麗. 蓮子皮低聚原花青素分級分離、組分鑒定與抗氧化機理研究[D]. 長沙: 湖南農業大學, 2013: 72-84.
[55] 侯成成, 麻微微, 肖榮. 染料木黃酮對星形膠質細胞氧化損傷中Nrf2/ARE通路相關因子表達的調節作用[J]. 中國食品衛生雜志, 2013, 25(1): 1-5. DOI:10.13590/j.cjfh.2013.01.023.
[56] 張倩. 金絲桃苷對Nrf2-ARE途徑的影響及其對肝細胞氧化應激損傷的保護作用[D]. 重慶: 西南大學, 2014: 52-58.
[57] 趙媛. 丹皮酚對局灶性腦缺血的保護作用及對pAkt、Nrf2和HO-1調節作用的實驗研究[D]. 石家莊: 河北醫科大學, 2013: 19-21. DOI:10.7666/d.Y233660.
[58] ZHANG Hui, LIU Yuanyuan, JIANG Qin, et al. Salvianolic acid A protects RPE cells against oxidative stress through activation of Nrf2/HO-1 signaling[J]. Free Radical Biology and Medicine, 2014, 69: 219-228. DOI:10.1016/j.freeradbiomed.2014.01.025.
[59] LEE T M, LIN S Z, CHANG N C. Antiarrhythmic effect of lithium in rats after myocardial infarction by activation of Nrf2/HO-1 signaling[J]. Free Radical Biology and Medicine, 2014, 77: 71-81. DOI:10.1016/j.freeradbiomed.2014.08.022.
[60] QIN Si, DENG Fangming, WU weiguo, et al. Baicalein modulates Nrf2/Keap1 system in both Keap1-dependentand Keap1-independent mechanisms[J]. Archives of Biochemistry and Biophysics, 2014, 559: 53-61. DOI:10.1016/j.abb.2014.03.011.
Plant Polyphenols Exert Antioxidant Activity of by Nrf2/ARE Signaling Pathway: A Review
LIAO Xia1, ZHENG Shaojie1, LU Keke1, XIAO Xingning1, WU Surui2, MING Jian1,3,*
(1. College of Food Science, Southwest University, Chongqing 400715, China; 2. Kunming Edible Fungi Institute of All China Federation of Supply and Marketing Cooperatives, Kunming 650223, China; 3. Chongqing Engineering Research Center of Regional Food, Chongqing 400715, China)
Polyphenols are important secondary metabolites widely distributed in the plant kingdom. They are well known as antioxidants which reduce ROS-induced damage and scavenge free radicals. The Nrf2/ARE signaling pathway is one of the most important pathways involved in preservation of antioxidant activity and defense against oxidative stress release in cells. This review outlines the composition and modulatory mechanism of the Nrf2/ARE signaling pathway. Plant polyphenols enhance oxidative stress resistance, reduce apoptosis, provide nerve protection, resist aging and mitigate oxidative damage through the Nrf2/ARE signaling pathway. All these evidence suggests that plant polyphenols have great potentials to be utilized in foods and drugs.
plant polyphenols; Nrf2/ARE; signaling pathway; antioxidant activity
10.7506/spkx1002-6630-201607041
Q964.8;R151.2
A
1002-6630(2016)07-0227-06
廖霞, 鄭少杰, 盧可可, 等. 植物多酚通過Nrf2/ARE信號通路抗氧化作用研究進展[J]. 食品科學, 2016, 37(7): 227-232. DOI:10.7506/spkx1002-6630-201607041. http://www.spkx.net.cn
LIAO Xia, ZHENG Shaojie, LU Keke, et al. Plant polyphenols exert antioxidant activity of by Nrf2/ARE signaling pathway: a review[J]. Food Science, 2016, 37(7): 227-232. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201607041. http://www.spkx.net.cn
2015-07-22
國家自然科學基金面上項目(31471576);“十二五”國家科技支撐計劃項目(2013BAD16B01);中央高校基本科研業務費專項資金項目(XDJK2015D035)
廖霞(1992—),女,碩士研究生,研究方向為食品化學與營養學。E-mail:994671521@qq.com
*通信作者:明建(1972—),男,教授,博士,研究方向為食品化學與營養學。E-mail:mingjian1972@163.com
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中國生殖健康(2019年3期)2019-02-01 06:12:26
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
西南軍醫(2015年2期)2015-01-22 09:09:37
