香蕉枯萎病菌SIX2基因序列及特異性分析
2016-11-24 07:25:04楊臘英郭立佳梁昌聰王國(guó)芬陳平亞黃俊生
植物保護(hù) 2016年5期
楊臘英, 郭立佳, 汪 軍, 劉 磊,梁昌聰, 王國(guó)芬, 陳平亞, 黃俊生
(中國(guó)熱帶農(nóng)業(yè)科學(xué)院環(huán)境與植物保護(hù)研究所, 農(nóng)業(yè)部熱帶農(nóng)林有害生物入侵檢測(cè)與控制重點(diǎn)實(shí)驗(yàn)室, 海口 571101)
?
香蕉枯萎病菌SIX2基因序列及特異性分析
楊臘英, 郭立佳, 汪 軍, 劉 磊,梁昌聰, 王國(guó)芬, 陳平亞, 黃俊生*
(中國(guó)熱帶農(nóng)業(yè)科學(xué)院環(huán)境與植物保護(hù)研究所, 農(nóng)業(yè)部熱帶農(nóng)林有害生物入侵檢測(cè)與控制重點(diǎn)實(shí)驗(yàn)室, 海口 571101)
依據(jù)侵染的香蕉品種范圍不同,引起香蕉枯萎病的尖孢鐮刀菌古巴專化型(Fusariumoxysporumf.sp.cubense,Foc)分為4個(gè)生理小種。Foc在寄主植物木質(zhì)部分泌的SIX(secreted in xylem)蛋白可能與不同生理小種侵染的香蕉品種范圍不同存在密切關(guān)系,找到 Foc 4號(hào)生理小種(Foc4)特有的SIX蛋白編碼基因?qū)⒂欣谶M(jìn)一步分析Foc4寄主范圍更廣的原因,從而開展抗病育種工作。采用PCR方法比較分析了國(guó)內(nèi)不同地理區(qū)域及來源于澳大利亞與南非的Foc1、Foc2、Foc3、Foc4共56株菌株中的SIX2基因,并以8個(gè)以上其他專化型或其他種或?qū)俚臒釒ё魑锊≡?9株菌株分析了Foc4SIX2基因序列的特異性,分析了SIX2基因的靈敏度及利用其檢測(cè)感病植株。僅從供試的Foc4菌株基因組DNA中擴(kuò)增出SIX2基因序列,檢測(cè)的DNA靈敏度達(dá)5 pg/25 μL,并可用于檢測(cè)感病的球莖組織。Foc4中特有的SIX2基因序列為特異性鑒定感病植株病原菌種類提供了快速分子檢測(cè)技術(shù),為明確該基因是否決定性影響Foc4對(duì)寄主差異性的選擇研究提供了基礎(chǔ)。
尖孢鐮刀菌古巴專化型;SIX2; 寄主選擇性; 分子檢測(cè)
香蕉枯萎病在世界主要香蕉產(chǎn)區(qū)先后發(fā)生并造成了極為嚴(yán)重的危害,是目前困擾世界香蕉產(chǎn)業(yè)發(fā)展的一大難題。該病由土壤真菌尖孢鐮刀菌古巴專化型(Fusariumoxysporumf.sp.cubense,簡(jiǎn)稱Foc)引起。依據(jù)Foc對(duì)不同品種香蕉的侵染特性將其劃分為4個(gè)生理小種(race):其中1號(hào)生理小種(Foc 1)危害基因型為AAB的‘Gros Michel’及栽培種;2號(hào)生理小種危害‘Bluggoe’蕉及基因型為ABB 的相關(guān)煮食蕉類;3號(hào)生理小種危害不是香蕉類的‘Heliconia’種類,因而被Ploetz等認(rèn)定其不為Foc[1-2];4號(hào)生理小種(Foc 4)則危害‘Cavendish’及其他對(duì)1號(hào)與2號(hào)生理小種敏感的蕉類。Foc4號(hào)生理小種更進(jìn)一步被分為熱帶4號(hào)生理小種(TR4)與亞熱帶4號(hào)生理小種(ST4),其中,TR4在熱帶與亞熱帶區(qū)域均能侵染‘Cavendish’蕉類,而ST4則僅能在亞熱帶區(qū)域侵染‘Cavendish’蕉類[3-4],且主要在植物暴露于非生物脅迫下發(fā)生危害,例如低溫和水澇。在我國(guó)廣東、廣西、云南、海南、福建局部地區(qū)有危害粉蕉(MusaABB ‘Pisang Awak’)的1號(hào)生理小種(Foc1)和危害巴西蕉(MusaAAA ‘Giant Cavendish cv. Bax’)和粉蕉的4號(hào)生理小種(Foc4)。國(guó)內(nèi)外尚無有效的控制該病的手段,使用抗病品種是控制香蕉枯萎病最根本的出路[5],而對(duì)相關(guān)致病基因的認(rèn)識(shí)最終可能有助于發(fā)展抗香蕉枯萎病的香蕉品種[6]。
多種植物病原菌,包括細(xì)菌、真菌、卵菌、線蟲,分泌一些蛋白質(zhì)和其他分子到其寄主植物的不同細(xì)胞區(qū)間以調(diào)節(jié)植物的防御體系及使其寄生性定殖[7-9]。在許多情況下,真菌感染特定植物種類的能力取決于其基因組中編碼寄主選擇性的“毒力因子”,這些因子可用于區(qū)分菌株的致病與不致病能力[10]。尖孢鐮刀菌在與寄主的相互作用中會(huì)分泌幾個(gè)特定的富含半胱氨酸的小分子量蛋白(15.8 ~29.9 kD)進(jìn)入木質(zhì)部中啟動(dòng)致病力,被稱為SIX(secreted in xylem)蛋白。近來更多的對(duì)尖孢鐮刀菌番茄專化型(F.oxysporumf.sp.lycopersici,簡(jiǎn)稱Fol)的相關(guān)研究,揭示了SIX基因在Fol與番茄互作系統(tǒng)中的致病作用,到目前為止,通過反向遺傳學(xué)等方法鑒定了Fol中的SIX1~SIX14共14個(gè)SIX基因[11-18]。
Foc侵入植株后,需定殖在寄主植物的木質(zhì)部,不同生理小種侵染的香蕉品種存在的差異性與Foc在寄主植物木質(zhì)部的成功定殖與否可能存在密切關(guān)系。因此Foc菌株亦可能啟動(dòng)相似基因表達(dá)分泌相關(guān)蛋白來決定Foc在寄主木質(zhì)部的定殖。Meldrum等[19]從不同地理來源的Foc 4小種中分別克隆鑒定了SIX1、SIX7、SIX8基因,而測(cè)試的Foc1與Foc2中僅發(fā)現(xiàn)SIX1基因。郭立佳等[20]研究發(fā)現(xiàn)香蕉枯萎病菌4號(hào)生理小種(Foc4)中存在SIX1、SIX2、SIX6、SIX8基因,亞熱帶4號(hào)生理小種中存在SIX7基因。筆者所在實(shí)驗(yàn)室通過比較完成的Foc1菌株N2與Foc4菌株B2的全基因組測(cè)序發(fā)現(xiàn)Foc4菌株中存在特異性的SIX2基因[21],為證實(shí)SIX2基因?yàn)镕oc4中特有,繼而明確該基因是否決定Foc4對(duì)寄主的差異性選擇,采用PCR方法比較分析了來自國(guó)內(nèi)不同地理區(qū)域及來源于澳大利亞與南非的不同F(xiàn)oc1與Foc4菌株中的SIX2基因,并以多個(gè)其他專化型或其他病原菌屬的菌株驗(yàn)證了該基因的序列特異性,同時(shí)為分析應(yīng)用該基因開展田間病株的快速檢測(cè)的可行性,分析了設(shè)計(jì)的基因引物序列的靈敏度,對(duì)病株不同染病部位開展了檢測(cè)分析。
1 材料與方法
1.1 材料
1.1.1 試驗(yàn)材料
共93株測(cè)試菌株,其中來源于澳大利亞的BW1(Foc1)、Race2(Foc2)、Race3(Foc3)、BW4(ST4)以及2株非病原尖孢鐮刀菌AB-11-5-2、AB-2-8的基因組DNA由廣東省東莞市香蕉蔬菜研究所呂順研究員贈(zèng)送;來源于南非的race1(Foc1)、TR4(Foc4)、 STR4(Foc ST4)共3株由廣東省農(nóng)業(yè)科學(xué)院果樹研究所李春雨研究員贈(zèng)送;國(guó)內(nèi)不同區(qū)域的2株Foc1(菌株編號(hào):Foc1 c2、Foc1 e2)和6株Foc4(菌株編號(hào):YF-10-12、YF-10-43、HBW 12、12、NX-27、PB3-15)菌株及5株尖孢鐮刀菌其他專化型菌株苦瓜專化型F.oxysporumf.sp.momordicae(Fom)菌株FO kugua、冬瓜專化型F.oxysporumf.sp.benincasae(Fob)菌株FO donggua、黃瓜專化型F.oxysporumf.sp.cucumerinum(Fou)菌株FO huanggua、節(jié)瓜專化型F.oxysporumf.sp.chiehqua(Foh)菌株FO jiegua、甜瓜專化型F.oxysporumf.sp.melonis(Foe)菌株FO tiangua由華南農(nóng)業(yè)大學(xué)王振中教授贈(zèng)送;甘藍(lán)專化型F.oxysporumf.sp.conglutinans(Foco)菌株Fo、FGL-12-48、FGL-13-1、FGL-13-8、FCH-12-9共5株由中國(guó)農(nóng)業(yè)科學(xué)院蔬菜花卉研究所楊宇紅副研究員惠贈(zèng);西瓜專化型F.oxysporumf.sp.niveum(Fon) 菌株22、75、624、Fon-01共4株由河北省農(nóng)林科學(xué)院孔令曉研究員惠贈(zèng);番茄專化型F.oxysporumf.sp.lycopersici(Fol) oxy-1及3株未知F.oxysporum專化型菌株oxy-2、oxy-3、oxy-4由北京市農(nóng)林科學(xué)院蔬菜研究中心贈(zèng)送;2株串珠鐮刀菌F.moniliforme(菌株編號(hào):FJAT-829、FJAT-866)、2株茄病鐮刀菌F.solani(菌株編號(hào):FJAT-3752、FJAT-3756)及1株桃褐腐病菌Monilinialaxa(菌株編號(hào):FJAT-772)由福建省農(nóng)業(yè)科學(xué)院劉波研究員贈(zèng)送;1株橡膠樹流膠病病原菌F.solani(菌株編號(hào):XJLJ4)及13株非鐮刀菌菌株ND-12、ND-120(劍麻斑馬紋病病菌Phytophthoranicotianae)、菌株P(guān)N006(橡膠樹褐根病病菌Phellinusnoxius)、菌株CH008(柱花草炭疽病病菌Colletotrichumgloeosporioides)、菌株RC178(橡膠樹炭疽病病菌C.acutatum)、菌株HJ-2、HJ-10(劍麻莖腐病病菌Aspergillusniger)、菌株Baifen(橡膠樹白粉病病菌Oidiumheveae)、菌株HHNBT01、HHNTC01(橡膠棒孢霉落葉病病菌Corynesporacassiicola)、菌株mFQ-2(香蕉黑條葉斑病病菌Mycosphaerellafijiensis)、菌株Wei 1、MYD3(香蕉葉斑病菌Cercosporasp.)的基因組DNA分別由筆者所在單位的特色蔬菜花卉病害與線蟲課題組、特色熱作病害課題組、橡膠樹病害課題組、熱帶果樹病害課題組贈(zèng)送;其他7株Foc1(菌株編號(hào):N2、BXFZ-1 b-1、BXFZ-2 a-2、40#①、40#②、42#①、11#②)與32株Foc4菌株(菌株編號(hào):B2、3#①、3#②、5#①、 5#②、6#①、6#②、7#①、7#②、9#①、9#②、34#①、 34#②、36#①、36#②、37#①、37#②、QWXZ b-2、NXXZ、NBXZ a、DTXZ-1 a-1、DTXZ-2W、DTXZ-3W、LDXZ a、LGXD-5W、LGXD-6W、ZJXD C、ZJB1 b1、ZJB2 b2、XNB2 a、XNB5 a、XNB6 a)由筆者所在實(shí)驗(yàn)室分離鑒定并保存。
1.1.2 試劑
rTaq酶、dNTPs、pMD18-T 載體購(gòu)自TaKaRa公司,PCR Product Purification Kit購(gòu)自上海博彩生物工程公司,引物由生工生物工程(上海)股份有限公司合成,其他常規(guī)化學(xué)生物試劑購(gòu)自試劑銷售公司。
1.2 方法
1.2.1 供試菌株的培養(yǎng)
分別將冷藏保存的供試菌種接種于PDA平板上活化,5 d后用打孔器在平板上打取菌餅(直徑0.5 cm),挑取10塊菌餅接入裝有500 mL馬鈴薯葡萄糖培養(yǎng)液的1 L三角瓶中,于28℃、180 r/min振蕩培養(yǎng)5 d,離心收集菌體用于提取DNA。
1.2.2 引物序列的設(shè)計(jì)
利用DNAStar version 5.0.221.0中的MegAlign比較分析測(cè)序的供試菌株的部分基因組序列和已在NCBI中登錄的F.oxysporumf.sp.lycopersici(Fol)(AJ608703.3和GQ268949.1)與F.fujikuroi(HF679028.1)的SIX2基因序列,設(shè)計(jì)SIX2基因引物Foc-six2-f/Foc-six2-r。
各菌株內(nèi)參基因的擴(kuò)增應(yīng)用rDNA核苷酸序列引物ITS1、ITS4;Foc4特異性引物Foc-1/Foc-2參照Lin等[22]的設(shè)計(jì)合成;香蕉組織內(nèi)參基因Actin的擴(kuò)增引物Actin2-F/Actin2-R參照Dita等[23]的設(shè)計(jì)合成。各個(gè)基因引物序列見表1。

表1 本研究涉及的引物序列信息Table 1 Primer sequences used for PCR analysis in this study
1.2.3 菌株基因組DNA、RNA與感病植株基因組DNA的提取
以改進(jìn)的SDS法提取各菌株基因組DNA[24];以接種B2菌株搖培過濾菌液后發(fā)病的香蕉植株為樣本,分別抽提球莖組織與感病葉片葉柄總DNA,感病植株基因組DNA參照劉金福等[21]的方法提取。抽提所得DNA均用紫外分光光度計(jì)檢測(cè)其純度及濃度,以A260/A280的比值為1.8~2.0時(shí)最佳,未達(dá)到要求的均以酚氯仿再次抽提DNA,直至符合試驗(yàn)要求。調(diào)整DNA的濃度至約20 ng/μL,置于-20℃冰箱中保存?zhèn)溆谩?/p>
菌體總RNA的提取采用異硫氰酸胍法。將經(jīng)檢測(cè)為完整的RNA反轉(zhuǎn)錄成完整的cDNA,具體步驟參照TaKaRa RNA PCR Kit說明書中反轉(zhuǎn)錄步驟進(jìn)行。
1.2.4 SIX2基因?qū)闎W4基因組DNA擴(kuò)增的靈敏度分析
用無菌水對(duì)提取的BW4菌株的總DNA進(jìn)行梯度稀釋,得到濃度分別為50 ng/μL、5 ng/μL、500 pg/μL、50 pg/μL、5 pg/μL、500 fg/μL、50 fg/μL和5 fg/μL的BW4總DNA溶液。分別以各濃度的BW4總DNA溶液中的核酸為模板,以無菌水為對(duì)照,進(jìn)行PCR擴(kuò)增反應(yīng)。
1.2.5 PCR反應(yīng)體系及產(chǎn)物檢測(cè)方法
反應(yīng)體系為:TaKaRaTaqDNA聚合酶(5 U/μL) 0.2 μL,10×PCR buffer (Mg2+Plus) 2.5 μL,dNTPs (2.5 mmol/L) 2 μL,模板1 μL,20 μmol/L上下游引物各0.5 μL,加水補(bǔ)足至25 μL。
反應(yīng)條件為: 94℃預(yù)變性5 min;94℃變性30 s,各引物序列對(duì)應(yīng)退火溫度退火30 s,72℃延伸45 s,35個(gè)循環(huán);最后72℃延伸10 min。
反應(yīng)產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳后用凝膠成像系統(tǒng)(BIO-BAD Gel Doc XR)觀察并拍照。
1.2.6 SIX2基因測(cè)序及序列分析
將PCR產(chǎn)物經(jīng)1%的瓊脂糖電泳檢測(cè)后用PCR回收試劑盒回收,與pMD18-T載體連接,轉(zhuǎn)化XL-1菌株挑選陽(yáng)性克隆提取質(zhì)粒酶切,送由生工生物工程(上海)股份有限公司測(cè)序。利用DNAStar v. 5.0.221.0預(yù)測(cè)SIX2蛋白序列,利用MegAlign比較分析本研究獲得的B2菌株的SIX2基因序列與GenBank中登錄的Fol分離物的SIX2基因序列(AJ608703.3和GQ268949.1)及F.fujikuroi(HF679028.1中3707948~3708646間堿基序列)所預(yù)測(cè)的SIX2蛋白序列差異性。
2 結(jié)果與分析
2.1 香蕉枯萎病菌SIX2基因擴(kuò)增與序列比較分析
設(shè)計(jì)合成的Foc-six2-f/Foc-six2-r引物對(duì)僅能從測(cè)試的3株Foc4菌株基因組DNA與cDNA中PCR擴(kuò)增出699 bp的目的條帶,以Foc1 N2菌株基因組DNA與cDNA為模板則均不能擴(kuò)增出目的條帶(圖1)。分別回收DNA與cDNA的PCR產(chǎn)物后連接至pMD18-T載體,比較分析SIX2基因的DNA與cDNA擴(kuò)增序列,發(fā)現(xiàn)Foc4小種的SIX2基因無內(nèi)含子。
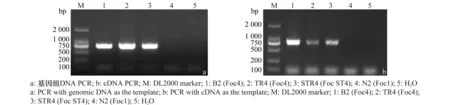
圖1 以不同F(xiàn)oc生理小種的基因組DNA和cDNA擴(kuò)增SIX2Fig.1 PCR products of SIX2 gene using genomic DNA and cDNA from four Foc isolates
DNAStar軟件預(yù)測(cè)獲得的B2SIX2基因編碼的蛋白質(zhì)序列由233個(gè)氨基酸組成,分子量約為25.5 kD,等電點(diǎn)為6.5左右。對(duì)該序列的信號(hào)肽預(yù)測(cè)結(jié)果表明Foc4菌株的SIX2蛋白質(zhì)序列存在信號(hào)肽區(qū)域(信號(hào)肽預(yù)測(cè)網(wǎng)址:http:∥www.cbs.dtu.dk/services/SignalP/),剪切位置位于第20號(hào)與21號(hào)氨基酸之間。將B2菌株的 SIX2蛋白質(zhì)序列與Fol及F.fujikuroi的SIX2蛋白質(zhì)序列進(jìn)行MegAlign比較分析,結(jié)果顯示B2菌株的 SIX2與Fol及F.fujikuroi的SIX2氨基酸序列存在較大差別,但在第100、111、116、171、199、202、209、232這8個(gè)位置均存在保守的半胱氨酸殘基。
2.2 尖孢鐮刀菌古巴專化型不同生理小種及尖孢鐮刀菌其他專化型SIX2基因擴(kuò)增結(jié)果
PCR擴(kuò)增結(jié)果表明(圖2),供試菌株中,共40個(gè)Foc4菌株的基因組DNA中可擴(kuò)增出目的條帶,而從國(guó)內(nèi)采集的粉蕉枯萎病病樣上分離的Foc1菌株及來源于南非的Foc1菌株、澳大利亞的Foc1菌株、Foc2菌株、Foc3菌株共12個(gè)菌株的基因組DNA中無目的條帶擴(kuò)增出來,同時(shí)試驗(yàn)中供試的18株尖孢鐮刀菌其他專化型菌株(苦瓜專化型、冬瓜專化型、黃瓜專化型、節(jié)瓜專化型、甜瓜專化型、番茄專化型、甘藍(lán)專化型、西瓜專化型、3株未知專化型菌)的基因組DNA中亦沒有擴(kuò)增出目的條帶。結(jié)果表明設(shè)計(jì)的引物對(duì)尖孢鐮刀菌古巴專化型4號(hào)生理小種具有非常強(qiáng)的特異性,同時(shí)亦再次證實(shí)尖孢鐮刀菌古巴專化型4號(hào)小種(Foc)中的SIX2基因序列與番茄專化型(Fol)中的SIX2基因序列存在較大的差異性。
利用Foc4的特異引物Foc-1/Foc-2對(duì)供試的71個(gè)菌株進(jìn)行驗(yàn)證,結(jié)果表明,該引物對(duì)僅能從各Foc4菌株基因組DNA中擴(kuò)增出217 bp的目的條帶(圖3),與SIX2基因引物對(duì)各菌株的擴(kuò)增結(jié)果一致。

圖2 尖孢鐮刀菌古巴專化型不同生理小種及尖孢鐮刀菌其他專化型SIX2基因產(chǎn)物Fig.2 Amplification products of SIX2 gene from Foc isolates belonging to different races and other formae specialis of Fusarium oxysporum

圖3 引物Foc-1/Foc-2的特異性驗(yàn)證Fig.3 Specificity verification for primer Foc-1/Foc-2
2.3 鐮刀菌屬其他種及其他屬菌株中SIX2基因擴(kuò)增結(jié)果
驗(yàn)證分析了設(shè)計(jì)的Foc4SIX2基因引物Foc-1/Foc-2的特異性,結(jié)果(圖4)表明,除陽(yáng)性對(duì)照,屬于Foc4小種的B2菌株基因組中擴(kuò)增出目的條帶外,陰性對(duì)照屬于Foc1小種的N2菌株及供試的其他21個(gè)菌株基因組DNA中均沒有擴(kuò)增出目的條帶。
2.4 SIX2基因引物Foc-six2-f/ Foc-six2-r的靈敏度
以菌株BW4基因組DNA 為模板測(cè)定SIX2基因引物Foc-six2-f/ Foc-six2-r的靈敏度,結(jié)果(圖5)顯示,50 ng/25μL(泳道1)、5 ng/25μL(泳道2)、500 pg/25μL(泳道3)、50 pg/25μL(泳道4)、5 pg/25μL(泳道5)均可以擴(kuò)增到699 bp的目的條帶,500 fg/25μL(泳道6)、50 fg/25μL(泳道7)、5 fg/25μL(泳道8)和陰性對(duì)照(泳道9)均不能得到任何條帶。

圖4 鐮刀菌屬不同種及其他屬菌株的SIX2基因PCR擴(kuò)增結(jié)果Fig.4 Amplification products of SIX2 gene from different species of Fusarium and other genus isolates
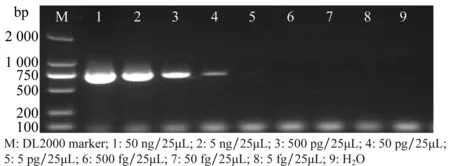
圖5 SIX2基因引物Foc-six2-f/ Foc-six2-r的靈敏度Fig.5 Sensitivity of specific primers Foc-six2-f/ Foc-six2-r for Foc4 SIX2 gene
2.5 從發(fā)病植株檢測(cè)香蕉枯萎病菌
分別以發(fā)病香蕉植株球莖與黃化葉片的葉柄總DNA為模板,以清水和4號(hào)生理小種菌株DNA做對(duì)照,利用設(shè)計(jì)的Foc4SIX2基因引物對(duì)香蕉組織內(nèi)參引物Actin2-F/Actin2-R[20]進(jìn)行多重PCR檢測(cè)。結(jié)果(圖6)顯示,提取的香蕉球莖組織與黃化葉片葉柄的DNA均能擴(kuò)增出217 bp的Actin2目的條帶,香蕉感染B2的球莖組織DNA及4號(hào)生理小種陽(yáng)性對(duì)照得到699 bp的SIX2基因條帶,而黃化的香蕉葉片的葉柄DNA無SIX2基因條帶擴(kuò)增產(chǎn)物。由此可判斷,Foc4SIX2基因引物對(duì)可用于田間所采集的感病香蕉的快速鑒定,也表明所建立的多重PCR檢測(cè)體系適用于對(duì)罹病的香蕉球莖組織進(jìn)行檢測(cè)。
3 討論與結(jié)論
Meldrum等[21]設(shè)計(jì)的引物不能從香蕉枯萎病菌中擴(kuò)增出目的條帶,筆者通過香蕉枯萎病菌全基因組序列比較分析后設(shè)計(jì)的擴(kuò)增SIX2基因的引物可以從供試的各個(gè)Foc4菌株中擴(kuò)增出699 bp的目條帶,這是由于Meldrum等設(shè)計(jì)的引物序列處于差異性較大的堿基區(qū)域所致。B2 菌株SIX2氨基酸序列與Fol及F.fujikuroi的SIX2氨基酸序列存在較大差別,但在相同位置均存在8個(gè)保守的半胱氨酸殘基, Chakrabarti等[26]推測(cè)尖鐮孢西瓜專化型不同生理小種SIX6 蛋白之間存在的差異氨基酸可能決定了不同生理小種的致病力,筆者推測(cè)B2 SIX2蛋白序列中表現(xiàn)出差異的氨基酸亦有可能有致病因子的功能,待后期通過利用同源重組轉(zhuǎn)化方法獲得SIX2基因敲除的突變菌株后分析其致病力與生理生化指標(biāo)變化情況等證實(shí)。

圖6 利用SIX2基因?qū)Ω胁≈仓隀z測(cè)結(jié)果Fig.6 Detection of SIX2 gene from diseased plants
鑒于尖孢鐮刀菌基因橫向轉(zhuǎn)移的證據(jù)[15],基于關(guān)鍵致病相關(guān)基因的病原菌分子診斷比基于系統(tǒng)發(fā)育相關(guān)基因更可靠。Fraser-Smith等[27]以SIX8基因序列為基礎(chǔ)設(shè)計(jì)了區(qū)分Foc4小種及亞熱帶Foc4小種的特異性引物。本研究中利用SIX2基因序列引物對(duì)僅能從Foc4中擴(kuò)增出特異性的目的條帶,與已有的Lin等[22]的用于區(qū)分Foc4的引物Foc-1/Foc-2的擴(kuò)增結(jié)果一致。同時(shí)本研究中設(shè)計(jì)的FocSIX2基因引物序列的靈敏度可達(dá)5 pg/25μL反應(yīng)體系,可以應(yīng)用于快速檢測(cè)鑒定田間發(fā)病香蕉枯萎病植株病樣及種植地土壤中病原菌的種類,為及時(shí)掌握香蕉枯萎病發(fā)生情況并采取有效的田間防控措施提供有力的技術(shù)支持,尤其可保障各組培苗公司出廠的香蕉種苗健康無菌,從而防止病原菌隨種苗的遠(yuǎn)距離傳播。
[1] Ploetz R C, Thomas J E, Slabaugh W R.Diseases of banana and plantain [M]∥Ploetz R C,ed. Diseases of tropical fruit crops. CABI Publishing, Wallingford, 2003: 73-134.
[2] Ploetz R C.Panama disease: an old nemesis rears its ugly head. Part 2.The Cavendish era and beyond [J].Plant Health Progress, 2006(206):1-17.
[3] Ploetz R C, Pegg K G.Fusarium wilt [M]∥Jones D R, ed. Diseases of banana, abacá and enset. CABI Publishing, Wallingford, 2000:143-159.
[4] Buddenhagen I W.Understanding strain diversity inFusariumoxysporumf.sp.cubenseand history of introduction of ‘tropical race 4’ to better manage banana production [M]∥Jones D, Van den Bergh I, eds. Proceedings of the International Symposium on Recent Advances in Banana Crop Protection for Sustainable Production and Improved Livelihoods, White River, South Africa. ISHS Acta Horticulturae, 2009, 828:193-204.
[5] Michielse C B, Rep M.Pathogen profile update:Fusariumoxysporum[J]. Molecular Plant Pathology, 2009, 10(3): 311-324.
[6] Fourie G, Steenkamp E T, Ploetz R C, et al. Current status of the taxonomic position ofFusariumoxysporumformaespecialiscubensewithin theFusariumoxysporumcomplex [J]. Infection, Genetics and Evolution, 2011, 11: 533-542.
[7] Block A, Li Guangyong, Fu Zhengqing, et al. Phytopathogen type III effector weaponry and their plant targets [J]. Current Opinion in Plant Biology, 2008, 11(4): 396-403.
[8] Davis E L, Hussey R S, Mitchum M G, et al. Parasitism proteins in nematode-plant interactions[J]. Current Opinion in Plant Biology, 2008, 11(4): 360-366.
[9] Misas-Villamil J C, van Der Hoorn R A.Enzyme-inhibitor interactions at the plant-pathogen interface [J]. Current Opinion in Plant Biology, 2008, 11(4): 380-388.
[10]Lievens B, Rep M, Thomma B P H J.Recent developments in the molecular discrimination of formae speciales ofFusariumoxysporum[J]. Pest Management Science, 2008, 64(8): 781-788.
[11]Rep M, van Der Does H C, Meijer M, et al. A small, cysteine-rich protein secreted byFusariumoxysporumduring colonization of xylem vessels is required for I-3-mediated resistance in tomato [J]. Molecular Microbiology, 2004, 53(5): 1373-1383.
[12]Houterman P M, Speijer D, Dekker H L, et al. The mixed xylem sap proteome ofFusariumoxysporum-infected tomato plants [J]. Molecular Plant Pathology, 2007, 8(2): 215-221.
[13]van Der Does H C, Rep M.Virulence genes and the evolution of host specificity in plant-pathogenic fungi[J]. Molecular Plant-Microbe Interactions, 2007, 20(10): 1175-1182.
[14]Lievens B, Houterman P M, Rep M.Effector gene screening allows unambiguous identification ofFusariumoxysporumf.sp.lycopersiciraces and discrimination from other formae speciales [J]. FEMS Microbiology Letters, 2009, 300(2): 201-215.
[15]Ma Lijun, van Der Does H C, Borkovich K A, et al. Comparative genomics reveals mobile pathogenicity chromosomes inFusarium[J]. Nature, 2010, 464(7287): 367-373.
[16]Rep M, Kistler H C.The genomic organization of plant pathogenicity inFusariumspecies [J]. Current Opinion in Plant Biology, 2010, 13(4): 420-426.
[17]Takken F, Rep M.The arms race between tomato andFusariumoxysporum[J]. Molecular Plant Pathology, 2010, 11(2): 309-314.
[18]Schmidt S M, Houterman P M, Schreiver I, et al. MITEs in the promoters of effector genes allow prediction of novel virulence genes inFusariumoxysporum[J]. BMC Genomics, 2013, 14: 119.
[19]Meldrum R A, Fraser-Smith S, Tran-Nguyen L T T, et al. Presence of putative pathogenicity genes in isolates ofFusariumoxysporumf.sp.cubensefrom Australia[J]. Australasian Plant Pathology, 2012, 41(5): 551-557.
[20]郭立佳, 楊臘英, 彭軍, 等. 香蕉枯萎病病原菌Six同源基因的鑒定[J]. 熱帶作物學(xué)報(bào), 2013, 34(12):2391-2396.
[21]Guo Lijia, Han Lijuan, Yang Laying, et al. Genome and transcriptome analysis of the fungal pathogenFusariumoxysporumf.sp.cubensecausing banana vascular wilt disease [J]. PLoS ONE, 2014, 9(4): e95543.
[22]Lin Yinghong, Chang Jingyi, Liu E T, et al. Development of a molecular marker for specific detection ofFusariumoxysporumf.sp.cubenserace 4 [J]. European Journal of Plant Pathology, 2009, 123(3): 353-365.
[23]Dita M A, Waalwijk C, Buddenhagen I W, et al. A molecular diagnostic for tropical race 4 of the banana fusarium wilt pathogen [J]. Plant Pathology, 2010, 59(2): 348-357.
[24]楊臘英, 黃華平, 唐復(fù)潤(rùn), 等. 香蕉炭疽菌rDNA ITS區(qū)的分子鑒定與檢測(cè)[J]. 植物病理學(xué)報(bào). 2006, 36(3): 219-225.
[25]劉金福, 潘東明, 代立春, 等. 香蕉RAPD反應(yīng)體系的建立[J]. 中國(guó)農(nóng)學(xué)通報(bào), 2009, 25(16): 51-55.
[26]Chakrabarti A, Rep M, Wang B, et al. Variation in potential effector genes distinguishing Australian and non-Australian isolates of the cotton wilt pathogenFusariumoxysporumf.sp.vasinfectum[J]. Plant Pathology, 2011, 60 (2): 232-243.
[27]Fraser-Smith S, Czislowski E, Meldrum R A, et al. Sequence variation in the putative effector geneSIX8 facilitates molecular differentiation ofFusariumoxysporumf.sp.cubense[J]. Plant Pathology, 2014, 63(5): 1044-1052.
(責(zé)任編輯:田 喆)
Sequence and specificity analysis ofSIX2 gene ofFusariumoxysporumf.sp.cubense
Yang Laying, Guo Lijia, Wang Jun, Liu Lei, Liang Changcong, Wang Guofen, Chen Pingya, Huang Junsheng
(Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences,Key Laboratory of Pests Comprehensive Governance for Tropical Crops, Ministry of Agriculture, Haikou 571101,China)
Fusariumoxysporumf.sp.cubense(Foc), causal agent ofFusariumwilt of banana, has been divided into four races according to cultivar specificity. There may be a close relationship between the races infested different banana varieties and the proteins secreted in xylem (SIX) by Foc when it invaded into the host plant xylem. The specificSIXgene will be conductive to further analyzing the reasons for broader host range of Foc4 and helpful for the disease-resistant cultivar breeding. Using PCR and sequencing approach,SIX2 gene were analyzed in 56 Foc isolates collected from different geographic areas of Southern China, Australia and South Africa. The specificity and sensitivity of the primers were determined by using other thirty-nine isolates including eight otherF.oxysporumformae specialis, two non-pathogenF.oxysporumand other genus or species of some tropical plant pathogens. With the designed primers, the target band was detected only in the DNA of Foc4 isolates, and the sensitivity of the primers were 5 pg/25 μL and could be used to detect diseased bulb section. Therefore,SIX2 gene specific in Foc4 could be applied for rapid molecular identification of the pathogens in susceptible plants, which will provide a basis for further research on whether the gene is clearly a decisive influence on the host selection of Foc4.
Fusariumoxysporumf.sp.cubense;SIX2; host selection; molecular diagnosis
2015-09-29
2015-12-18
國(guó)家自然科學(xué)基金(31301629)
S 436.681
A
10.3969/j.issn.0529-1542.2016.05.020
致 謝: 感謝華南農(nóng)業(yè)大學(xué)王振中教授、廣東省農(nóng)業(yè)科學(xué)院果樹研究所李春雨研究員、北京市農(nóng)林科學(xué)院蔬菜研究中心、中國(guó)農(nóng)業(yè)科學(xué)院蔬菜花卉研究所楊宇紅研究員、河北省農(nóng)林科學(xué)院植保所孔令曉研究員、福建省農(nóng)業(yè)科學(xué)院劉波研究員、廣東東莞香蕉研究所呂順研究員及作者所在單位特色熱作病害課題組與橡膠樹病害課題組及熱帶果樹病害課題組同事們贈(zèng)送菌株,基于他們的支持才能較全面地驗(yàn)證本試驗(yàn)的相關(guān)研究結(jié)果,特此表示真誠(chéng)的謝意。
* 通信作者 E-mail:H888111@126.com
