新疆棉花產量和纖維品質性狀SSR關聯分析
2016-11-26 03:55:09艾先濤梁亞軍龔照龍王俊鐸鄭巨云李雪源
新疆農業科學 2016年11期
關鍵詞:新疆
艾先濤,梁亞軍,龔照龍,王俊鐸,鄭巨云,李雪源
(新疆農業科學院經濟作物研究所,烏魯木齊 830091)
?
新疆棉花產量和纖維品質性狀SSR關聯分析
艾先濤,梁亞軍,龔照龍,王俊鐸,鄭巨云,李雪源
(新疆農業科學院經濟作物研究所,烏魯木齊 830091)
【目的】研究新疆陸地棉品種資源目標相關性狀QTL緊密連鎖的分子標記,發掘與目標性狀相關的具有重要利用價值的基因源。【方法】利用54對SSR 標記,分析94 份新疆自育陸地棉品種的基因組。【結果】群體結構分析表明,94個品種總體被劃分為2個類群,同時還出現了1個混合群。關聯分析結果表明,在P值<0.05時,GLM(Q)模型中共有 36個標記位點與纖維品質性狀和農藝產量性狀關聯,其中與比強度、馬克隆值、長度、伸長率、鈴重、生育期和衣分相關聯的位點分別為12、4、13、1、4、1和1個。【結論】主成分分析和群體結構分析從不同角度反映了新疆棉花品種的親緣關系的遠近,自育品種遺傳多樣性不夠豐富,在分子水平上差異并不大,需要努力拓寬品種選育的遺傳基礎。
陸地棉;遺傳多樣性;SSR;關聯分析
0 引 言
【研究意義】新疆是我國最重要的植棉大省,棉花總產已占據全國50%以上,肩負著我國棉花有效供給和安全的重大責任。當前新疆棉花生產面臨的主要問題是棉花單產潛力需要進一步挖掘,棉花纖維品質不能適應現代紡織工業的要求。如何進一步提高產量和改良纖維品質一直是育種的重點。棉花產量和纖維品質性狀屬于多基因控制的數量性狀。隨著分子生物學技術的發展和數量遺傳學理論的創新,分析和發掘控制數量性狀的基因(QTL)并利用分子標記輔助選擇(MAS),對育成棉花品種的產量和品質進行改良有重大意義。【前人研究進展】20世紀90年代以來,棉花的分子遺傳學研究有了較快的發展。通過開發分子標記、構建連鎖圖譜和建立分離群體進行連鎖分析,一系列控制棉花產量和纖維品質性狀的數量性狀基因位點被定位到染色體或連鎖群上。到目前為止,棉花上已經報道的與產量和纖維品質相關的QTL已達上千個,其中50%左右是利用陸地棉的分離群體進行檢測。雖然近10年來已有上千個棉花經濟性狀的QTL定位文獻報道。但在這些已經報道的QTL中,有一部分來自于海陸種間雜交群體分析,難以直接應用于陸地棉的標記輔助育種;另外一部分QTL則來自于陸地棉的兩親本雜交群體,具有較強的“群體專一性”,因為遺傳背景的影響,用于陸地棉標記輔助育種時仍然存在一定的困難。這些已定位的QTL,除少數在標記輔助選擇過程中被驗證之外,大多數QTL需在育種親本和種質資源群體中進一步證實,以有效地用于標記輔助選擇。研究前期已初步開展新疆自育品種農藝性狀和分子標記多樣性的研究[1-3],包括鑒定和評價陸地棉種質多樣性方面。【本研究切入點】新疆自育陸地棉資源遺傳多樣性水平較低,遺傳基礎較為狹窄。形態性狀鑒定和分子標記技術是研究種質資源多樣性和篩選優良種質的方法,二者從性狀表達及基因組水平兩方面反映了種質的多樣性,雖然存在差別,但也存在相互驗證的關系。【擬解決的關鍵問題】研究陸地棉全基因組基因發掘的新方法,篩選新疆品種資源目標相關性狀QTL緊密連鎖的分子標記,發掘出與目標性狀相關的具有重要利用價值的基因源。
1 材料與方法
1.1 材 料
收集新疆陸地棉栽培種數量共計94個(截止到2010年自育審定的品種),品種包括新陸早1~49號、軍棉1號、新陸中1~48號(不包括新陸中25、32、42、45號)。表1
表1 新疆陸地棉自育品種
Table 1 Local upland cotton cultivars released in Xinjiang

編號No.品種名稱Cultivars審定年限Yearauthorized系譜來源Pedigree選育單位BreedingInstitutes1新陸早1號1981(斯字5A×611波)的722選系中系選農八師下野地試驗站2新陸早2號198869-2×中棉所4號石河子棉花研究所3新陸早3號1988(車66-241×愛字無毒棉)×荊無4588農七師農科所4新陸早4號1994(車66-241×澧74-47W)×岱70農七師農科所5新陸早5號1994(科遺181×347-2)F1×(83-2-3+陜1155)石河子棉花研究所6新陸早6號199785-174×貝爾斯諾農七師農科所7新陸早7號1997自育優系347-2×塔什干2號石河子棉花研究所8新陸早8號1997(抗黃系V.Wx×新陸早1號)F1輻射石河子棉花研究所9新陸早9號1999(系5×貝爾斯諾)×中棉所17農七師農科所10新陸早10號1999(黑山棉1號×02Ⅱ)×中棉所12石河子棉花研究所11新陸早11號1999豫422×陜3215博州引進12新陸早12號2000從遼寧經作所引入遼棉95-25引進13新陸早13號2002自育83-14×(中無5601+1693)農七師農科所14新陸早14號2002新陸早7號×ZK90石河子棉花研究所15新陸早15號2002雞腳JW×中棉所12農七師農科所16新陸早16號2003早熟雞腳棉×貝爾斯諾農七師農科所17新陸早17號2004內地引進9908系選新疆農科院經作所18新陸早18號2004內地已近高代材料69118系選新疆農科院經作所19新陸早19號200491-2×自育品系900石河子棉花研究所20新陸早20號200597-185(新陸早16號)系選農八師150團21新陸早21號20051034系選石河子富依德公司22新陸早22號2005本所材料45-1×新陸早6號農墾科學院棉花所23新陸早23號2005中棉所27系選奎屯萬氏種業錦棉公司24新陸早24號2005中長絨品系7074×司6524康地種業25新陸早25號2006[(系5×貝爾斯諾)×晉14]×中棉所17農七師農科所26新陸早26號2006新陸早8號系選天合種業27新陸早27號20067147×貝爾斯諾康地種業28新陸早28號2006新陸早4號×(貝爾斯諾+西農大材料)惠遠公司29新陸早29號2006新陸早16號系選金博種業30新陸早30號2006新陸早16號系選金博種業31新陸早31號2006(新陸早6號×貝爾斯諾)×愛字棉奎屯萬氏種業32新陸早32號2006拉馬干77系選農墾科學院棉花所33新陸早33號2007石選87農墾科學院棉花所34新陸早34號200797-65×7003康地種業35新陸早35號2007[(新陸早3號×中2621)×抗35)]×新陸早16號農七師農科所36新陸早36號20071304×BD103石河子棉花研究所37新陸早37號2008(遼83421×系5)×(遼9001+系5+90-2)農五師農科所38新陸早38號2008(92-226×新陸早9號)×(中6331×中17)農七師農科所39新陸早39號2008(新陸早4號×貝爾斯諾)×愛字棉奎屯萬氏種業40新陸早40號2009新陸早16號×(D256×SW2)F2農墾科學院棉花所41新陸早41號200917-79巴州富全新科種業42新陸早42號2009新陸早10號×97-6-9農墾科學院棉花所43新陸早43號200941-4×H2石河子棉花所44新陸早44號2009MP1×FP1農墾科學院棉花所45新陸早45號2010新陸早13號×9941農墾科學院棉花所
續表1
表1 新疆陸地棉自育品種
Table 1 Local upland cotton cultivars released in Xinjiang

編號No.品種名稱Cultivars審定年限Yearauthorized系譜來源Pedigree選育單位BreedingInstitutes46新陸早46號2010系9×822石河子棉花研究所47新陸早47號2010(中17+9901)×新陸早16號農七師農科所48新陸早48號2010石選87×優系604惠遠公司49新陸早49號20109765×新陸早16號農七師農科所50軍棉1號1979[司1470×(五一大鈴+147大+司1470+早落葉棉+司3521+新海棉+2依3)]農二師34團良種繁育站51新陸中1號1988[(巴州6017×上海無毒棉)×巴6017]巴州農業科學研究所52新陸中2號1988麥克奈210×新陸201新疆農科院經作所53新陸中3號1989{【(108夫×C1470)×108夫】×137夫}陜401新疆農科院莎車試驗站54新陸中4號1992岱字棉45選×新陸202新疆農科院經作所55新陸中5號1994陜721×108夫新疆農科院經作所56新陸中6號1997巴州6017×上海無毒棉巴州農業科學研究所57新陸中7號199985-113×中棉所12兵團農一師農科所58新陸中8號1999[{C-8017×〈寧細6133-3+(910依×陸地棉)F4〉×11818×198}F8×(永年小雙桃×594依)F5]×海南野生棉}]新疆種子站引進59新陸中9號2000新陸中4號中系選新疆農科院經作所60新陸中10號2000從烏茲別克斯坦雜交組合高代材料(95-10)系選新疆農科院經作所61新陸中11號2000巴州7648×K-202巴州農業科學研究所62新陸中12號2000108夫系選108夫系選63新陸中13號2002(ND45×3287)×ND45(回交4次)新疆農業大學農學院64新陸中14號2002司6524×中棉所19兵團農一師農科所65新陸中15號2002(ND25×3287)×ND25(回交4次)新疆農業大學農學院66新陸中16號2003新陸中5號×(中棉所17+中棉所12+中棉所19)阿拉爾良繁場67新陸中17號2003協作92-36×中棉所17阿拉爾良繁場68新陸中18號2003冀9119×遼棉10號天然彩色棉有限公司69新陸中19號2004新900品系×貝爾斯諾新疆農業大學農學院70新陸中20號200489-19×中長絨棉材料33兵團種子站和天合種業71新陸中21號200492D×引進的豐產抗病品系96-07新疆農科院經作所72新陸中22號2005新陸中8號×抗病品系9658紅太陽種業公司73新陸中23號2005從獨聯體引進陸地棉高代材料(陸-4)系選吐魯番長絨棉研究所74新陸中24號2006胞質不育系H-1038A×海島棉恢復系1304R兵團農一師農科所75新陸中26號2006從引進17-79中系選出巴棉3號再由巴棉3號選育出6603品系系選而成巴州富全新科種業76新陸中27號2006自育032品系×品系048天合種業公司77新陸中28號2006中9409(中棉所35)×父本邯109個人選育78新路中29號2006J95-8×11-6新疆優質雜交棉公司79新陸中30號2006不祥新疆優質雜交棉公司80新陸中31號2007不育系KA×恢復系01588康地種業有限公司81新陸中33號2007036×渝棉1號巴州富全新科種業82新陸中34號2007品系8316×中99+系選巴州農業科學研究所83新陸中35號2007不詳巴州富全新科種業84新陸中36號20089119×155石大85新陸中37號2008B23×渝棉1號塔河種業86新陸中38號200999-26康地種業87新陸中39號2009KA×H3康地種業88新陸中40號2009新陸早16號×(D256×SW2)F2庫爾勒種子公司89新陸中41號2009巴州6807×Acala1517巴州農科所90新陸中43號200941-4×H2農一師農科所塔河種業91新陸中44號2010石遠321×中棉所19號巴州富全新科種業92新陸中46號2010中棉所12號×(新植1號+15號)河南正林,禾春州種業93新陸中47號2010Ji98-72×1099巴州農科所94新陸中48號2010[(99-708(C6524×中棉所19號)]×99-425農一師農科所塔河種業
1.2 方 法
SSR標記分析部分實驗,棉花總DNA提取與純化采用CTAB法提取棉花基因組DNA,即小量法DNA提取。棉花總DNA純度和濃度的測定由基因公司生產的Nanodrop-ND-1000全波長紫外/可見光掃描分光光度計檢測DNA純度和濃度。讀取A260和A280吸光值。DNA濃度(ng/pl)=稀釋倍數×50×A260,可根據A260/A280來判斷DNA純度 ,比值若在1.6~2.0,證明DNA純度較好。大于2.0,證明RNA污染;小于1.6,證明蛋白質、酚等污染。根據前人(Xiao,2009)文獻資料,篩選均勻分布棉花26條染色體的SSR標記,遺傳圖譜大小約4140cM,以每20cM大小為間距選擇已定位于染色體的SSR標記共計206對,引物包括DPL、CGR、CER、DC、BNL和JESPR等類型。引物序列等信息均來自(http://www.ncbi.nlm.nih.gov/和http://www.cottondb.org)。引物由上海生工生物工程有限公司合成。實驗電泳所用凝膠為8%非變性聚丙烯酰胺凝膠。
2 結果與分析
2.1 基于SSR標記的主成分(PCA)
主成分分析使品種地域性和遺傳背景關系表現得更為明顯, 為品種分析提供了重要的信息。基于遺傳相似系數,在NTSYS2.1軟件上對SSR多態性位點作了主成分分析,得到第1、2主成分的貢獻率分別達到了20.11%和6.40%,前5主成分累計貢獻率達到了39.98%。對94 份品種,以第一主成分為x軸,第二主成分為y軸作三維散點圖,形成的各品種位置分布,位置相近表示關系密切,遠離者表示關系疏遠。將位置靠近的品種劃歸在一起,研究表明,主成分分析結果與聚類分析結果基本一致,主成分分析結果更直觀地表明了不同品種之間的親緣關系的遠近。
從主坐標分析所得的三維散點圖看出,兩大群體的分布具有規律性,第一主坐標的右半部分主要為第一類群品種,左半部分則主要為第二類群品種。表2,圖1
表2 前5主成分貢獻率及累計貢獻率
Table 2 Characteristic values and their contributions of the prior five principal components
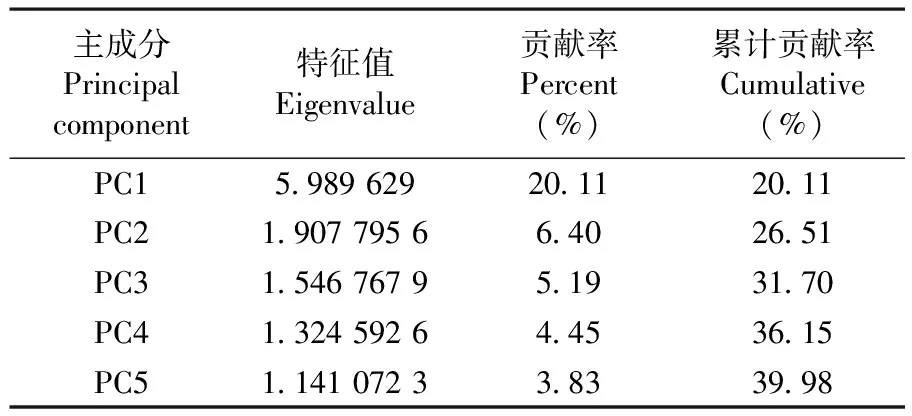
主成分Principalcomponent特征值Eigenvalue貢獻率Percent(%)累計貢獻率Cumulative(%)PC1598962920112011PC2190779566402651PC3154676795193170PC4132459264453615PC5114107233833998
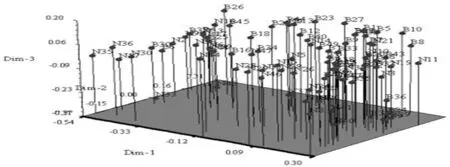
圖1 SSR數據 PCA聚類結果
Fig. 1 The cluster analysis results of PCA based on SSR data
2.2 基于SSR標記的群體結構
為進一步明確94份新疆陸地棉品種間的遺傳組成并將遺傳相似性高的材料劃分在同一類群中,用Structure2.3軟件對品種進行群體結構分析。設置K值參數為1~10,即將94個新疆自育品種預先劃分為1~10個亞群,每一個K值獨立運行3次。根據遺傳相似性比例及群體結構圖將94份品種劃分到相應亞群中,其余劃分為混合群。
用LnP(D)對K作圖,橫坐標為K值,相應的LnP(D)值為縱坐標做曲線圖,當出現明顯的拐點時,此處對應的橫坐標即為所需要的K值,如果拐點不明顯,再對LnP(D)求二階導數,得到△k,然后用△k對K作圖,當出現明顯拐點時最終確定所需要的K值,此時的K值即為群體數目。對于94份品種來說,通過structure軟件運算結果,可以看出,當K=2時曲線出現了一個明顯的拐點(圖2a),然后求得LnP(D)的二階導數△K,以△K作曲線圖(圖2b),同時也發現K=2時拐點明顯;且K=2時,按遺傳相似性比例為0.7進行劃分,將94份品種大致劃分2個群體,分別記為pop1和pop2。pop1群體有23份品種,pop2群體有58份品種,mixed群體有13份品種。說明當K=2時,類群劃分能得到理想的結果。所以將K值確定為2,即將94份棉花核心種質材料劃分為兩個群體圖由也可以看出K=2時,可以比較清晰的將新疆自育陸地棉品種劃分為兩大類,結果與聚類結果大致相似。群體結構將94個品種劃分為兩個類群的同時,又出現了一個混合群,這一點與聚類結果有所不同。分析兩者差異,可能是由于兩種方法分析的角度不同造成的,因為群體結構主要是基于模型角度進行研究的。表3~4,圖2
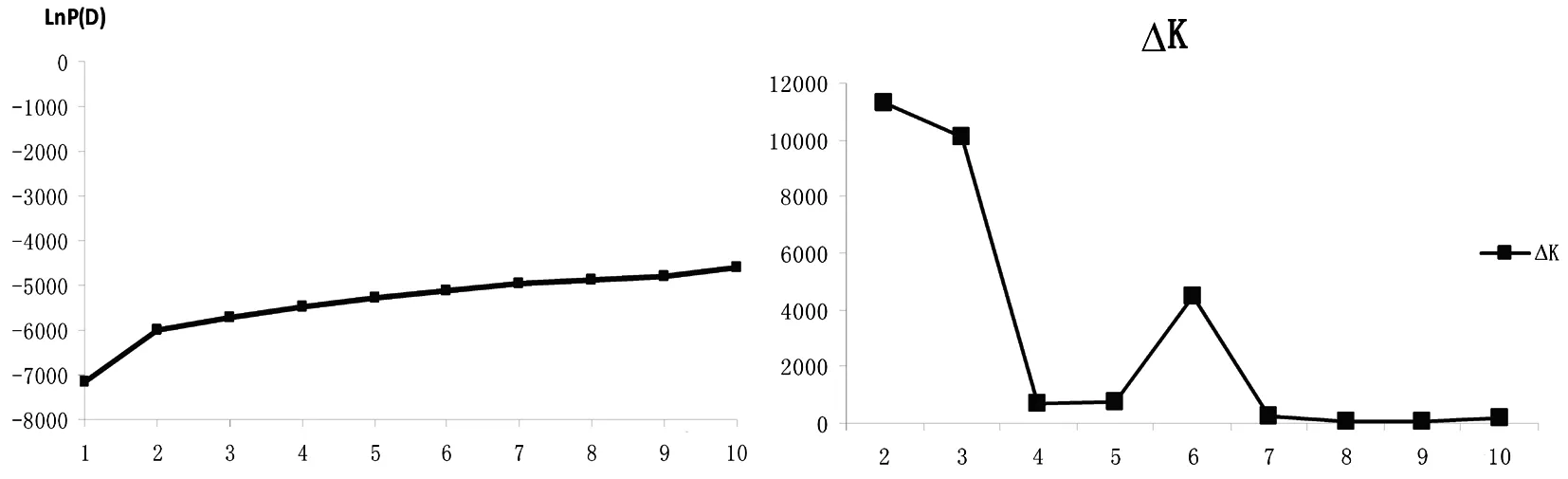
圖2 94份材料K值(a)及△K(b)曲線
Fig.2 Curve diagram of K(a) and △K (b) value of 94 cultivars
表3 94份新疆自育品種遺傳相似性比例
Table 3 The propotion of genetic similarity of local upland cotton cultivars in Xinjiang
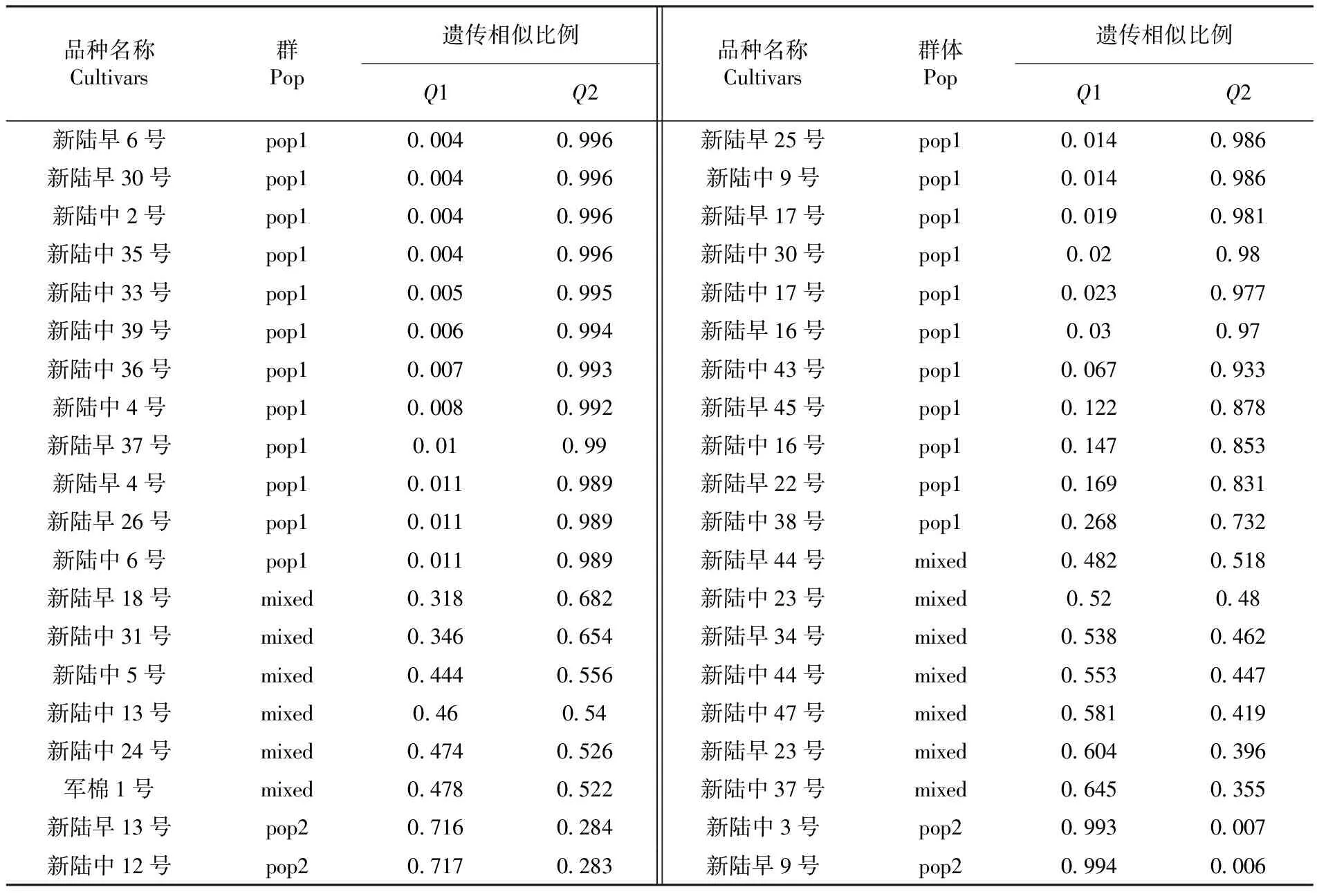
品種名稱Cultivars群Pop遺傳相似比例Q1Q2品種名稱Cultivars群體Pop遺傳相似比例Q1Q2新陸早6號pop100040996新陸早25號pop100140986新陸早30號pop100040996新陸中9號pop100140986新陸中2號pop100040996新陸早17號pop100190981新陸中35號pop100040996新陸中30號pop1002098新陸中33號pop100050995新陸中17號pop100230977新陸中39號pop100060994新陸早16號pop1003097新陸中36號pop100070993新陸中43號pop100670933新陸中4號pop100080992新陸早45號pop101220878新陸早37號pop1001099新陸中16號pop101470853新陸早4號pop100110989新陸早22號pop101690831新陸早26號pop100110989新陸中38號pop102680732新陸中6號pop100110989新陸早44號mixed04820518新陸早18號mixed03180682新陸中23號mixed052048新陸中31號mixed03460654新陸早34號mixed05380462新陸中5號mixed04440556新陸中44號mixed05530447新陸中13號mixed046054新陸中47號mixed05810419新陸中24號mixed04740526新陸早23號mixed06040396軍棉1號mixed04780522新陸中37號mixed06450355新陸早13號pop207160284新陸中3號pop209930007新陸中12號pop207170283新陸早9號pop209940006
續表3
表3 94份新疆自育品種遺傳相似性比例
Table 3 The propotion of genetic similarity of local upland cotton cultivars in Xinjiang
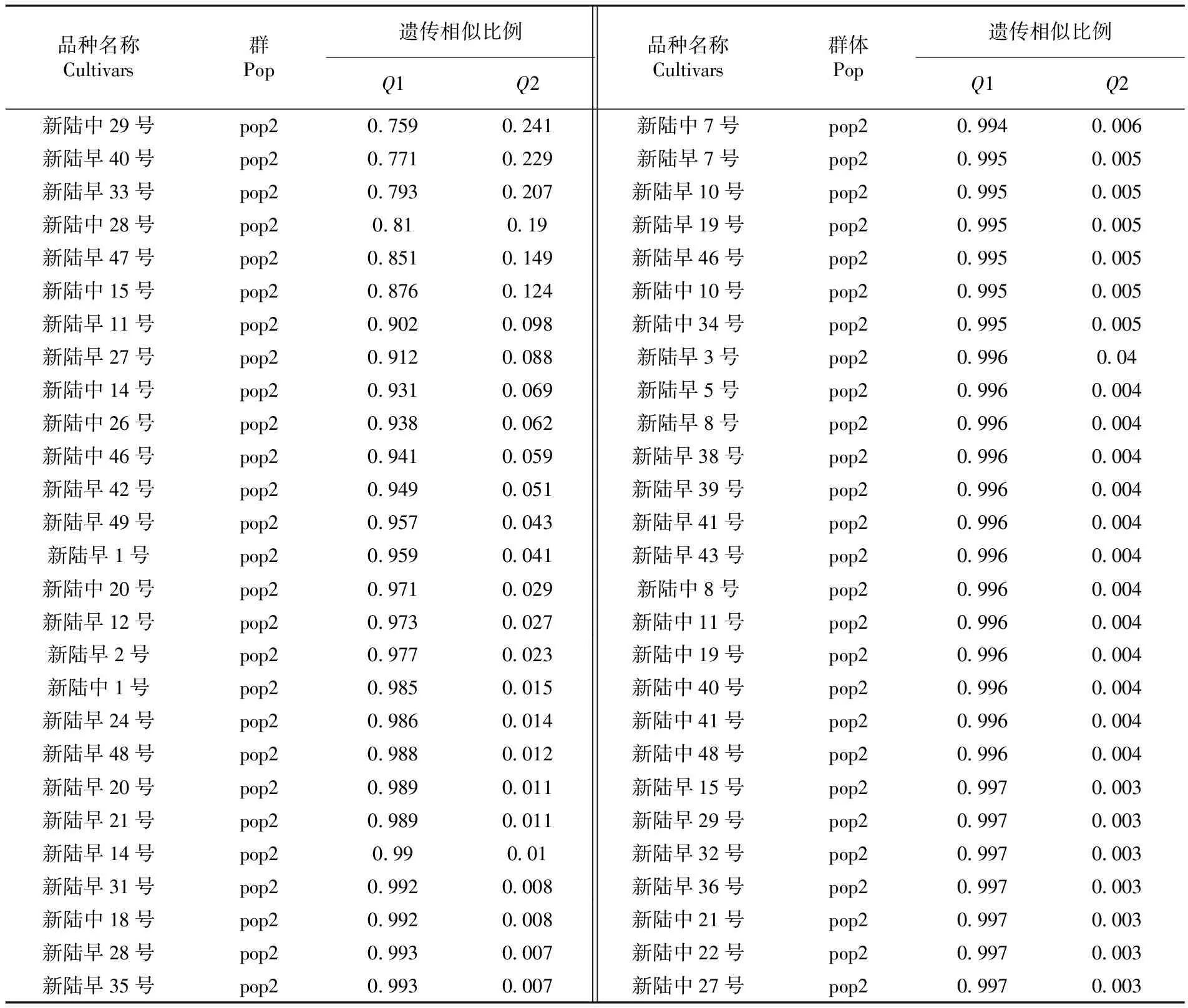
品種名稱Cultivars群Pop遺傳相似比例Q1Q2品種名稱Cultivars群體Pop遺傳相似比例Q1Q2新陸中29號pop207590241新陸中7號pop209940006新陸早40號pop207710229新陸早7號pop209950005新陸早33號pop207930207新陸早10號pop209950005新陸中28號pop2081019新陸早19號pop209950005新陸早47號pop208510149新陸早46號pop209950005新陸中15號pop208760124新陸中10號pop209950005新陸早11號pop209020098新陸中34號pop209950005新陸早27號pop209120088新陸早3號pop20996004新陸中14號pop209310069新陸早5號pop209960004新陸中26號pop209380062新陸早8號pop209960004新陸中46號pop209410059新陸早38號pop209960004新陸早42號pop209490051新陸早39號pop209960004新陸早49號pop209570043新陸早41號pop209960004新陸早1號pop209590041新陸早43號pop209960004新陸中20號pop209710029新陸中8號pop209960004新陸早12號pop209730027新陸中11號pop209960004新陸早2號pop209770023新陸中19號pop209960004新陸中1號pop209850015新陸中40號pop209960004新陸早24號pop209860014新陸中41號pop209960004新陸早48號pop209880012新陸中48號pop209960004新陸早20號pop209890011新陸早15號pop209970003新陸早21號pop209890011新陸早29號pop209970003新陸早14號pop2099001新陸早32號pop209970003新陸早31號pop209920008新陸早36號pop209970003新陸中18號pop209920008新陸中21號pop209970003新陸早28號pop209930007新陸中22號pop209970003新陸早35號pop209930007新陸中27號pop209970003
表4 新疆自育陸地棉群體劃分
Table 4 The population division of local upland cotton cultivars in Xinjiang

群Groups數目品種名稱Cultivarspop123新陸早4、6、16、17、22、25、26、30、37、45號新陸中2、4、6、9、16、17、30、33、35、36、38、39、43號mixed13新陸早18、23、34、44號新陸中5、13、23、24、31、37、44、47號、軍棉1號pop258新陸早1、2、3、5、7、8、9、10、11、12、13、14、15、19、20、21、24、27、28、29、31、32、33、35、36、38、39、40、41、42、43、46、47、48、49號新陸中1、3、7、8、10、11、12、14、15、18、19、20、21、22、26、27、28、29、34、40、41、46、48號
用distruct軟件包對structure軟件運行結果進行柱形圖繪制。圖3~5
2.3 衰減圖
因為 LD 衰減所延伸的距離決定著關聯分析的精度和所需標記的多寡,所以評價 LD隨物理或遺傳距離的下降速率很重要。以分布標記較多的 6、8、12、20 號染色體上的 SSR 標記位點的 D′值做其隨遺傳距離(cM)變化的衰減圖,通過對陸地棉的 D′與遺傳距離的衰減圖其衰減速率相對較慢。因此就這 4 條染色體來說做關聯分析時要達到作圖精度,陸地棉需要更多的標記。圖6
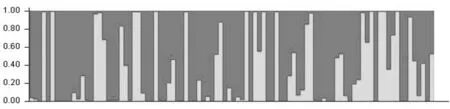
圖3 新疆自育陸地棉品種群體結構
Fig.3 The population structure of local upland cotton cultivars in Xinjiang(K=2)

圖4 經distruct軟件處理的包含2個群的群體結構
Fig.4 Model-based cluster membership for 94 cultivars in two groups
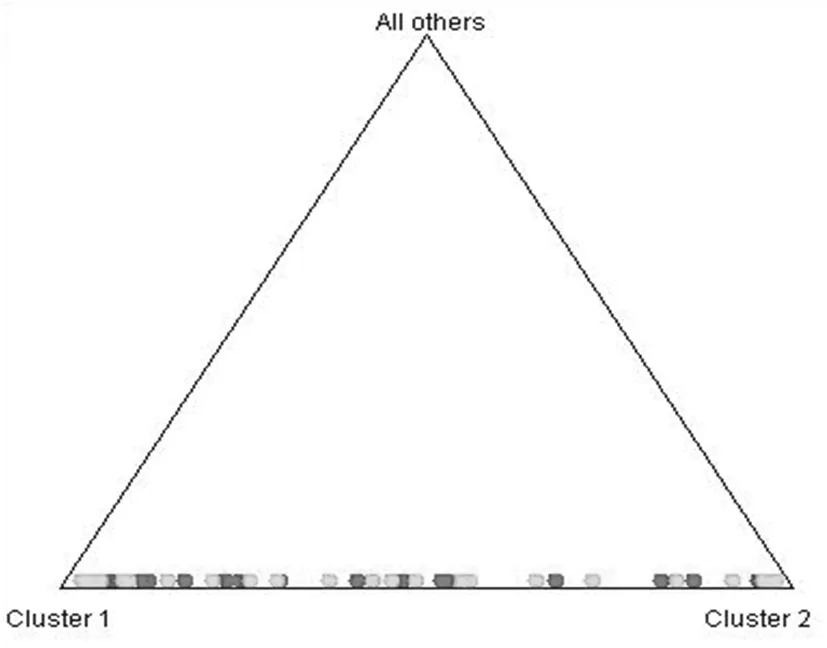
圖5 新疆自育品種群聚類
Fig.5 Clustering of local upland cotton cultivars in Xinjiang
2.4 關聯分析
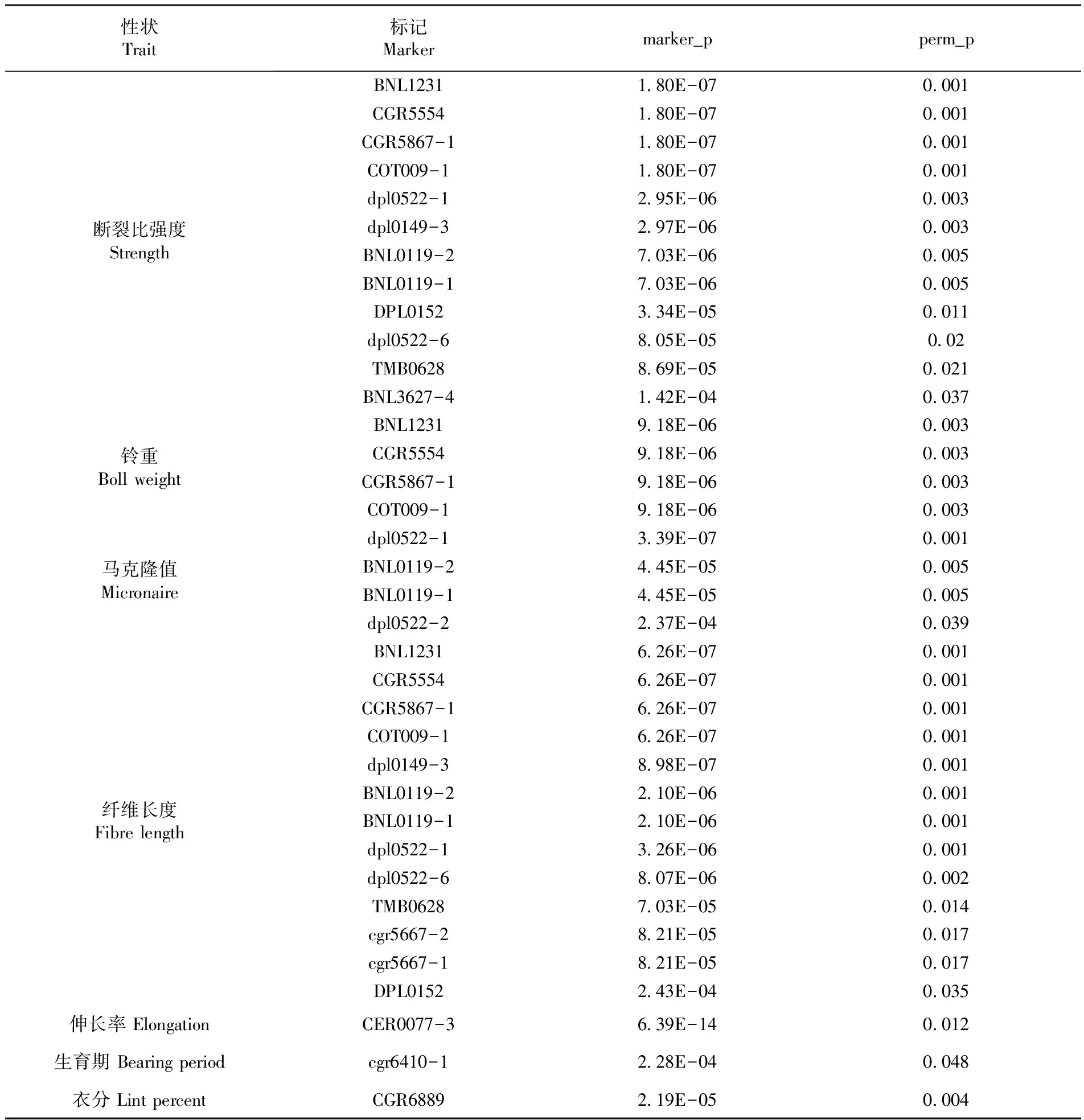
用TASSEL(http://www.maizegenetics.net)的 GLM(Mixed Linear Model)模型進行關聯分析。利用54對引物與兩個環境的四個重復所獲得纖維品質的長度,比強度,伸長率,馬克隆值、整齊度、生育期、衣分、鈴重、株高、子葉節高、籽指、單株鈴數、果枝臺數等 13個品質、產量和農藝性狀指標進行關聯分析,GLM 回歸方程是:Yj=α+βIPj+β1X1j+β2X2j+ … +βkXkj+ε公式中Yj為第j個材料數量性狀測定值;β是群體各位點各等位變異的平均效應;β1~βk是亞群體各位點各等位變異的平均效應;Ipj是第j材料第p等位變異出現的指示變量;X1j~Xkj是第j材料基因組變異源于1-k群體的概率值(Q值);ε是殘差。考慮群體結構對關聯分析的影響,采用 tassel 軟件的一般線性模型(GLM,未考慮親緣關系的Q模型)對標記位點和性狀進行關聯分析, 94份陸地棉關聯分析結果表明,在P值<0.05時,GLM(Q)模型中共有 36個標記位點與纖維品質性狀和農藝產量性狀關聯,其中與比強度、馬克隆值、纖維長度、伸長率、鈴重、生育期和衣分相關聯的位點分別為12、4、13、1、4、1和1個。表5
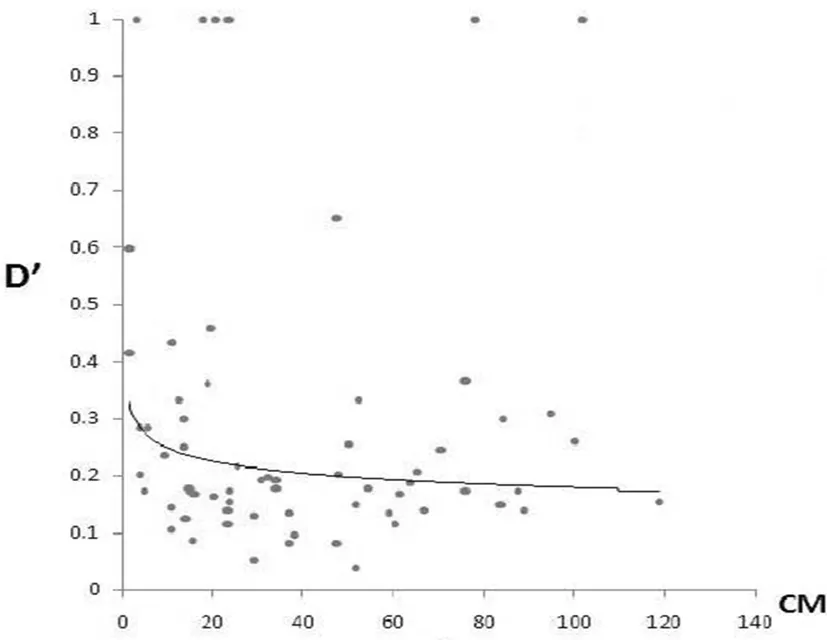
圖6 陸地棉 5、6、8、20 號染色體上的SSR位點隨圖距(CM)衰減散點圖
Fig.6 distribution of D′valus observed between marker pairs against genetic distance (CM)in chromsome5、Chromsome6、chromsome8 and chromsome20 of Gossypium hirsutum(A)
表5 顯著關聯標記信息匯總
Table 5 Summary of significant marker information associated
性狀Trait標記Markermarker_pperm_p斷裂比強度StrengthBNL1231180E-070001CGR5554180E-070001CGR5867-1180E-070001COT009-1180E-070001dpl0522-1295E-060003dpl0149-3297E-060003BNL0119-2703E-060005BNL0119-1703E-060005DPL0152334E-050011dpl0522-6805E-05002TMB0628869E-050021BNL3627-4142E-040037鈴重BollweightBNL1231918E-060003CGR5554918E-060003CGR5867-1918E-060003COT009-1918E-060003馬克隆值Micronairedpl0522-1339E-070001BNL0119-2445E-050005BNL0119-1445E-050005dpl0522-2237E-040039纖維長度FibrelengthBNL1231626E-070001CGR5554626E-070001CGR5867-1626E-070001COT009-1626E-070001dpl0149-3898E-070001BNL0119-2210E-060001BNL0119-1210E-060001dpl0522-1326E-060001dpl0522-6807E-060002TMB0628703E-050014cgr5667-2821E-050017cgr5667-1821E-050017DPL0152243E-040035伸長率ElongationCER0077-3639E-140012生育期Bearingperiodcgr6410-1228E-040048衣分LintpercentCGR6889219E-050004
3 討 論
艾先濤等[1]分子聚類將94份自育品種劃分為2個類群,反映了自育品種在基因組水平上差異,聚類結果整體反映出新陸早和新陸中品種間遺傳關系相對簡單,品種的遺傳基礎相對狹窄。主成分分析使品種地域性和遺傳背景關系表現得更為明顯和直觀,通過位置遠近表示關系疏密,使很清楚看到94個品種分布可以劃分為兩大群體,并表現出一定規律性。將位置靠近的品種劃歸在一起,結果表明主成分分析結果與聚類分析[1]結果基本一致,主成分分析結果更直觀地表明了不同品種之間的親緣關系的遠近。在群體結構分析中,94個品種盡管被劃分為兩個類群,但出現了一個混合群,這與分子聚類結果有所不同。這可能是由于兩種分析方法不同所造成的,因為群體結構分析主要是基于模型角度進行研究的。
利用SSR分子標記對新疆自育陸地棉品種的遺傳改良進行研究,明晰了新疆自育陸地棉品種間的親緣關系和群體結構,對于指導新疆棉花育種具有重要意義。研究結果反映新疆陸地棉品種間的遺傳相似性相對較高,總體上遺傳多樣性不夠豐富。自育品種在分子水平上差異并不大,遺傳基礎狹窄的現狀并未徹底改變,這一結論與前人的研究結果一致。別墅等[4]對我國三大主產棉區只選取了30個有代表性的品種作了品種遺傳多樣性作了研究,得出我國棉花大多數品種的遺傳基礎比較狹窄。刁明等[5]利用RAPD指紋圖譜研究了50 a來北疆棉花品種演變過程的遺傳多樣性,認為隨著品種的演變,品種群體間遺傳關系趨于復雜,遺傳基礎逐漸豐富,品種適應性、豐產性都得到加強。劉文欣等[6]對建國以來有代表性的166 個棉花主栽品種的遺傳多樣性研究表明:在我國主栽棉花品種中,海島棉品種遺傳基礎窄于陸地棉品種;我國自育陸地棉品種的遺傳基礎窄于國外引進品種;雜交陸地棉品種的遺傳基礎窄于常規品種;20世紀80 年代以后陸地棉品種遺傳基礎窄于70年代品種;長江棉區品種遺傳基礎窄于黃淮棉區品種,西北內陸棉區品種窄于長江棉區品種。陳光[7]對我國陸地棉基礎種質和衍生系的遺傳多樣性及親緣關系進行了分析,結果表明:第一、二、三期基礎種質分子標記平均遺傳相似系數分別是0.587、0.630、0.630,基礎種質基因組水平的差異呈下降的趨勢,棉花的育種背景逐漸變窄。黃河棉區基礎種質綜合性狀較好,長江棉區基礎種質產量性狀較高,北部特早熟棉區基礎種質早熟性好,美國品種在我國形成的基礎種質抗黃萎病性較強。長江、黃河流域兩大棉區的基礎種質在基因組水平上的差異變化明顯。杜雄明等[8]利用SSR引物對不同親本來源、不同選育時期、不同種植生態區的陸地棉基礎種質進行了遺傳多樣性分析,結果表明現代基礎種質比早期基礎種質在基因組水平的差異呈下降的趨勢。薛艷等[9,10]利用SSR對北疆42個新陸早聚類分析發現,無論是過去自育的主栽品種還是近年自育品種以及從外地引進的品種,在DNA水平上彼此間的差異并不是很大,絕大多數供試品種的親緣關系比較近,遺傳基礎比較狹窄。
由于實驗材料數量有限(94個),SSR標記數量少(54對),無法進一步對新疆自育品種進行全基因組關聯分析,因為關聯分析方法可以更有效的發掘自然群體中的優良基因。今后可以通過擴大群體數目和增加標記數,發掘有利基因,進一步利用分子手段更好的為育種服務。
4 結 論
4.1 對94 份品種進行主成分分析,分析結果更直觀地表明了不同品種之間的親緣關系的遠近。第一主坐標的右半部分主要為第一類群品種,左半部分則主要為第二類群品種。
4.2 群體結構將94個品種劃分為兩個類群的同時,又出現了一個混合群,這一點與聚類結果有所不同。分析兩者差異,這可能是由于兩種方法分析的角度不同造成的,因為群體結構主要是基于模型角度進行研究的。
4.3 94份陸地棉關聯分析結果表明,在P值<0.05時,GLM(Q)模型中共有 36個標記位點與纖維品質性狀和農藝產量性狀關聯,其中與比強度、馬克隆值、長度、伸長率、鈴重、生育期和衣分相關聯的位點分別為12、4、13、1、4、1和1個。
References)
[1] 艾先濤,梁亞軍,沙紅,等. 新疆自育陸地棉品種SSR 遺傳多樣性分析[J].作物學報,2014,40(2):369-379.
AI Xian-tao, LIANG Ya-jun, SHA Hong, et al. (2014). Genetic Diversity Analysis on Local Upland Cotton Cultivars in Xinjiang Based on SSR Markers [J].ActaAgronomicaSinica, 40(2):369-379. (in Chinese)
[2]鄭巨云,王俊鐸,艾先濤,等.陸地棉產量與纖維品質性狀的遺傳相關分析[J].新疆農業科學,2013,50(6):995-1002.
ZHENG Ju-yun, WANG Jun-duo,AI Xian-tao,et al.Inheritance Analysis of Upland Cotton Yield and Fiber Quality Characters[J]. Xinjiang Agricultural Sciences,2013,50(6):995-1002.(in Chinese)
[3]鄭巨云,龔照龍,王俊鐸,等.新疆陸地棉遺傳連鎖圖譜構建及葉綠素含量和光合速率的QTL定位[J].新疆農業科學,2014,51(9):1577-1582.
ZHENG Ju-yun,GONG Zhao-long,WANG Jun-duo,et al.Construction of Linkage Genetic Map and QTL Mapping for the Chlorophyll Content and Photosynthetic Rates in Upland Cotton[J]. Xinjiang Agricultural Sciences,2014,51(9):1577-1582. (in Chinese)
[4]別墅,孔繁玲,周有耀,等.中國3大主產棉區棉花品種遺傳多樣性的RAPD及其與農藝性狀關系的研究.中國農業科學,2001,34(6):597-603.
BIE Shu, KONG Fan-ling, ZHOU You-yao, et al. (2001). Genetic diversity analysis representative elite cotton varieties in three main cotton regions in China by RAPD and its relation with agronomic characteristics [J].ScientiaAgriculturaSinica, (34):597-603,(in Chinese)
[5] 刁明,褚貴新,李少昆,等.北疆50年來主栽棉花品種親緣關系的研究[J].中國農業科學,2002,35(12):1 456-1 460.
DIAO Ming, CHU Gui-xin, LI Shao-kun, et al. (2002). Studies on the Relationship of Upland Cotton Varieties in the Course of Replacement of Varieties in North Xinjiang over the Past Fifty Years [J].ScientiaAgriculturaSinica, 35(12):1,456-1,460. (in Chinese)
[6] 劉文欣,孔繁玲,郭志麗等.建國以來我國棉花品種遺傳基礎的分子標記分析[J]遺傳學報,2003,30(6):560-570.
LIU Wen-xin, KONG Fan-ling, GUO Zhi-li, et al. (2003). Analysis about genetic basis of cotton cultivars in China since 1949 with molecular markers [J].ActaGenetSin, 30(6):560-570. (in Chinese)
[7]陳光.我國陸地棉基礎種質及其衍生品種的遺傳多樣性[D].北京:中國農業科學院碩士研究生學位論文,2005.
CHEN Guang. (2005).GeneticDiversityofBasicGermplasmandTheirOffspringofUplandCottoninChina[D]. Master Dissertation. Graduate School of Chinese Academy of Agricultural Sciences, Beijing. (in Chinese)
[8]杜雄明,劉國強, 陳光.論我國棉花育種的基礎種質[J].植物遺傳資源學報,2004,(1):69-74.
DU Xiong-ming, LIU GUO-qiang, CHEN Guang. (2004).Basic germplasm for cotton breeding in China[J].JournalofPlantGeneticResources, (5):69-74. (in Chinese)
[9] 薛艷,張新宇,沙紅,等.新疆早熟棉品種SSR指紋圖譜構建與品種鑒別.棉花學報, 2010 ,22(4):360-366.
XUE Yan, ZHANG Xin-yu, SHA Hong, et al. (2010). Construction of Fingerprinting Map Based on SSR and Identification of Cultivars for Earliness Cultivars in Upland Cotton in Xinjiang [J].ActaGossypiiSin, 22(4):360-366. (in Chinese)
[10] Xiao, J., Wu, K., Fang, D., Davidm, S., Yu, J., & Royg, C. (2009). New ssr markers for use in cotton (gossypium spp.) improvement.JournalofCottonScience, 13(2):75-157.
Fund project:Xinjiang Natural Science Foundation(2012211B47)
Analysis of Xinjiang Cotton Yield and Fiber Quality Traits Associated with SSR
AI Xian-tao1,2, LIANG Ya-jun1, GONG Zhao-long1, WANG Jun-duo1,ZHENG Ju-yun1, LI Xue-yuan1
(1.Research Institute of Economic Crops, Xinjiang Academy of Agricultural Sciences, Urumqi, 830091,China; 2.CollegeofAgronomy,XinjiangAgriculturalUniversity,Urumqi,Xinjiang830052,China)
【Objective】 Screening closely related traits of Xinjiang upland cotton cultivars resources goals and objectives of the molecular markers,excavated chain has important value in use of genes related to the source traits.【Method】The genome of 94 self-fertile cotton varieties in Xinjiang was analyzed by 54 SSR markers.【Result】Population structure analysis showed that 94 cotton materials could be divided into two groups and one mixed group. The association analysis showed that 36 SSR loci were (P<0.05) associated with fiber quality and agronomic phenotype in GLM(Q) model, 12 loci were associated with strength, 4 loci were associated with fiber micronaire, 13 loci were associated with fiber length, 1 locus was associated with Elongation, 4 loci were associated with boll weight, 1loci was associated with growth process, 1 loci was associated with lint percentage.【Conclusion】Principal component analysis and population structure analysis reflect the proximity of the phylogenetic relationship of cotton varieties in Xinjiang from different angles. On the whole, the genetic diversity of self-fertile variety is not rich enough, and at the molecular level the differences are not great, which demonstrates that great effort should be made to broaden the genetic basis of breeding.
upland cotton; genetic diversity; SSR; association analysis
10.6048/j.issn.1001-4330.2016.11.002
2016-06-28
新疆維吾爾自治區自然科學基金項目(2012211B47)
艾先濤(1979-),男,陜西人,副研究員,研究方向為棉花遺傳育種, (E-mail)yixiantao@sina.com
李雪源(1964-),男,遼寧人,研究員,研究方向為棉花遺傳育種,(E-mail)xjmh2338@163.com
S562
A
1001-4330(2016)11-1969-11
猜你喜歡
國畫家(2022年2期)2022-04-13 09:07:46
四川文學(2021年4期)2021-07-22 07:11:54
新作文·高中版(2021年4期)2021-07-14 11:55:06
環球人物(2021年8期)2021-05-06 08:00:43
音樂天地(音樂創作版)(2020年12期)2021-01-11 01:13:40
音樂天地(音樂創作版)(2020年7期)2020-09-11 06:17:18
華人時刊(2018年15期)2018-11-10 03:25:32
人民中國(日文版)(2015年10期)2015-04-16 03:53:48
絲綢之路(2014年9期)2015-01-22 04:24:46
新疆人文地理(2009年7期)2009-09-29 09:56:14
