穩定表達NLRC5基因肝癌細胞株的建立
2016-11-28 03:48:12何鶯華倪明明孟曉明
安徽醫科大學學報 2016年1期
關鍵詞:肝癌
何鶯華,徐 濤,倪明明,黃 成,李 娟,孟曉明,李 俊
?
穩定表達NLRC5基因肝癌細胞株的建立
何鶯華1,徐 濤1,倪明明1,黃 成1,李 娟2,孟曉明1,李 俊1
目的 建立起能夠穩定表達NLRC5基因的肝癌HepG2細胞株。方法 設計遺傳霉素(G418)的濃度梯度,通過對HepG2 細胞篩選最終確定篩選藥物濃度。將構建好的人源pEGFP-C2-NLRC5重組質粒利用脂質體介導法轉染至HepG2肝癌細胞株中,用G418篩選出耐藥陽性克隆。通過免疫熒光技術觀察綠色熒光蛋白穩定表達情況,Western blot法驗證NLRC5蛋白表達情況。結果 G418在14 d內使HepG2細胞全部死亡的最小濃度是300 ng/ml。用G418篩選瞬轉pEGFP-C2-NLRC5 14 d在熒光顯微鏡下可見耐藥克隆的形成。Western blot法檢測顯示,穩定過表達組NLRC5的蛋白水平比空質粒轉染組高(t=15.356,P<0.05)。結論 成功構建穩定表達NLRC5基因的肝癌細胞株,為進一步研究NLRC5基因在肝癌的體內實驗奠定了基礎。
NLRC5;肝癌;穩定細胞株;HepG2
肝細胞肝癌約占原發性肝癌的90%,已成為全世界最常見的惡性腫瘤之一,其死亡率在腫瘤相關死因中位列第三[1]。而肝癌的發生、發展主要與肝臟慢性炎癥等免疫反應所導致的肝細胞反復損傷與增生密切相關[2-3],深入了解和掌握免疫反應的分子機制將為今后治療與炎癥相關的癌癥提供幫助。天然免疫系統中的核苷酸結合寡聚結構域樣受體(nucleotide-binding domain, leucine-rich repeat containing receptors,NLRs)家族成員之一,NLR家族含半胱天冬酶激活招募結構域5(NOD-like receptor family CARD domain containing 5, NLRC5)蛋白分子,其在抗病毒免疫過程、炎癥小體形成以及主要組織相容性復合體-1(major histocompatibility complex- 1, MHC-I)分子基因轉錄過程中均有參與,但其具體的機制尚不能確定[4]。研究[5]顯示NLRC5在人肝星狀細胞(LX-2)中能正向調節核轉錄因子κB(nuclear factor kappa B, NF-κB)/Smad3通路介導白介素-6(interleukin-6, IL-6)和白介素-1β(interleukin-1 beta, IL-1β)的分泌,提示NLRC5在肝癌的發生發展中可能有所參與。該研究建立能夠穩定過表達NLRC5的HepG2細胞株,為研究NLRC5在肝癌中的生物學功能奠定基礎。
1 材料與方法
1.1 材料
1.1.1 細胞系 人肝癌細胞株HepG2由本實驗室保存。
1.1.2 主要試劑 無內毒素質粒大提試劑盒(北京康為世紀生物有限公司);質粒小抽試劑盒(美國AxyGen公司);LB-Kana液體培養基由本實驗室配制;G418、LiprofectamineTM2000、Opti-MEM(美國Invitrogen公司);PCR擴增試劑盒、引物(上海生物工程有限公司);一抗(英國Abcam公司)。
1.2 方法
1.2.1 人源重組真核表達載體pEGFP-C2-NLRC5質粒抽提及鑒定 取微量pEGFP-C2和pEGFP-C2-NLRC5質粒轉化DH5α 感受態細菌,將其涂布于LB-Kana瓊脂培養基上,37 ℃倒置過夜,第2天挑取單個菌落接種于8 ml LB-Kana液體培養基,37 ℃、150 r/min搖菌14 h。20%甘油保留1 ml菌液,其余菌液用小抽試劑盒抽提質粒,瓊脂糖凝膠電泳鑒定陽性重組質粒菌液。將陽性重組質粒菌液擴大搖菌后使用去內毒素質粒大提試劑盒抽提質粒,瓊脂糖凝膠電泳鑒定并測定其濃度,-20 ℃保存。
1.2.2 確定G418篩選濃度 取對數生長期的HepG2細胞,胰酶消化,計數板計數,12孔板中每孔接種1 000個細胞。待細胞貼壁后,梯度加入G418,其濃度分別是100、200、300、400、500、600、700、800、900、1 000 ng/ml, 37 ℃、5% CO2培養箱培養,每隔1 d更換1640完全培養基1次,對每孔細胞生長情況多進行觀察。篩選14 d后,觀察每孔細胞死亡情況,將全部殺死細胞的最小濃度確定為最終藥物篩選濃度。
1.2.3 人源pEGFP-C2-NLRC5重組質粒轉染人肝癌細胞系HepG2 將HepG2細胞消化重懸接種至6孔板中,置37 ℃、5% CO2培養,待細胞融合生長至70%~80%進行轉染。將1 μg質粒和5 μl LiprofectamineTM2000分別溶于250 μl Opti-MEM培養基中,孵育5 min后將兩者混合,室溫靜置20 min。無菌PBS洗滌細胞2次后每孔加入1.5 ml Opti-MEM,再緩慢加入上述混合物,置37 ℃、5% CO2培養6 h后更換含1640完全培養液繼續培養,24 h后在熒光顯微鏡下觀察轉染情況并拍照,帶有綠色熒光的細胞即為重組質粒轉染成功的細胞,經G418篩選、單克隆后得到穩定過表達組,實驗中同時設置未轉染組和空質粒轉染組。
1.2.4 篩選穩定轉染細胞株 HepG2細胞經脂質體瞬時轉染24 h后,以300 ng/ml的G418藥物濃度篩選,每隔1~2 d更換含有G418的完全培養基,約6 d細胞出現大量死亡,換用特殊培養基培養。特殊培養基的制備方法:HepG2生長融合約達80 %,換液,培養過夜,收集培養基,濾器消毒,和新鮮培養基1 ∶1混合。待未轉染組細胞全部被殺死后繼續篩選,轉染組可見耐藥克隆形成。將耐藥克隆消化,有限稀釋法稀釋至96孔板中,每孔1個細胞,用特殊培養基培養,繼續加藥篩選,30 d后在熒光顯微鏡下選取陽性孔,接種至新的培養皿中擴大培養,培養期間保持300 ng/ml的G418藥物濃度,1周后進行鑒定。
1.2.5 Western blot法檢測穩定過表達組NLRC5的蛋白表達 取穩定過表達組、空質粒轉染組細胞,兩組細胞數相同,使用RIPA強裂解液(含1%的蛋白酶抑制劑)提取總蛋白,加蛋白上樣緩沖液,100 ℃、10 min后收好蛋白保存至-20 ℃。配8% SDS-PAGE凝膠,加25 μl蛋白,電泳、轉膜、脫脂奶粉封閉,TBST洗膜后,一抗孵育。次日TBST洗膜,在5%脫脂奶粉中加入辣根過氧化物酶標記的二抗室溫孵育1 h,TBST洗膜后,加入ECL發光劑,電腦顯影成像。
2 結果
2.1 質粒小抽后鑒定 將搖菌后的菌液按小抽試劑盒說明書抽提質粒,經1%瓊脂糖凝膠電泳,與DNA Marker比對,發現第5組質粒沒有在大小符合的位置出現條帶,說明這組單克隆菌液中并不含重組載體,應該丟棄。見圖1。
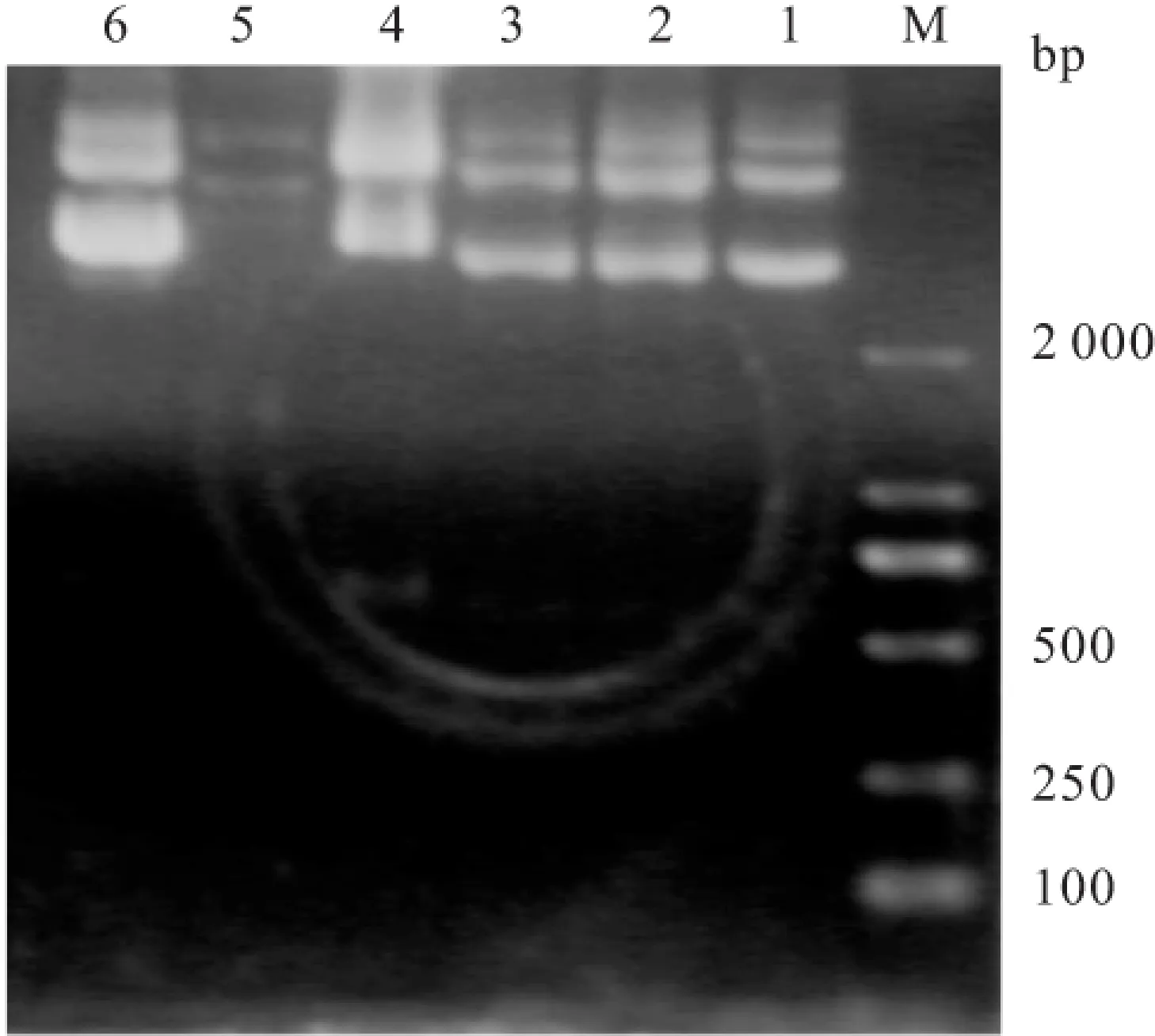
圖1 RT-PCR鑒定質粒小抽
M:Marker; 1:挑取的1號單克隆;2:挑取的2號單克隆;3:挑取的3號單克隆;4:挑取的4號單克隆;5:挑取的5號單克隆;6:挑取的6號單克隆
2.2 顯微鏡下觀察確定G418 篩選藥物濃度 接種HepG2細胞的12孔板,經不同濃度G418篩選14 d后在顯微鏡下觀察細胞的死亡情況,發現300 ng/ml以及比其濃度更高的G418濃度可將細胞全部殺死,最終確定G418篩選濃度為300 ng/ml。見圖2。
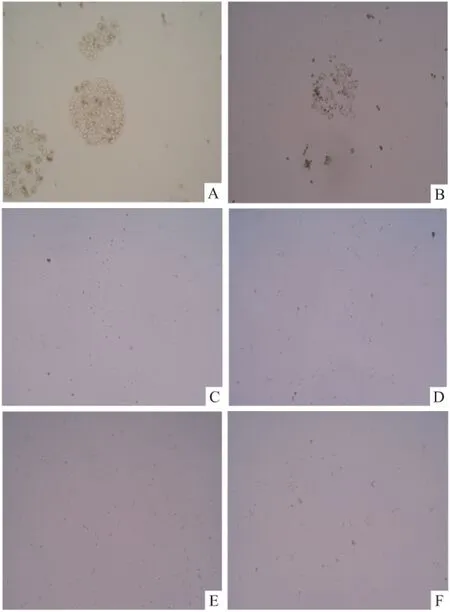
圖2 G418不同濃度篩選14 d后HepG2細胞結果 ×100
A:100 ng/ml;B:200 ng/ml;C:300 ng/ml;D:400 ng/ml;E:500 ng/ml;F:600 ng/ml
2.3 熒光顯微鏡下觀察穩定細胞株的篩選情況 細胞經脂質體轉染后,因重組質粒攜帶GFP綠色熒光基因以及 Neomysin 抗性基因,若質粒基因轉入細胞內,則在熒光倒置顯微鏡中可觀察到綠色熒光并能夠在G418 篩選壓力下存活。結果如圖3所示,轉染后24 h,熒光顯微鏡下可見綠色熒光細胞團;G418 持續篩選14 d后,單克隆化的細胞耐藥群形成。
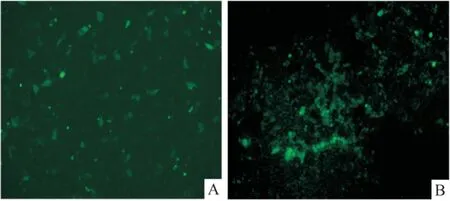
圖3 熒光顯微鏡下觀察NLRC5蛋白表達 ×200
2.4 穩定細胞株中NLRC5蛋白的表達 Western blot法檢測結果顯示,穩定過表達組和空質粒轉染組的細胞內均有NLRC5蛋白表達,穩定過表達組NLRC5蛋白表達明顯高于空質粒轉染組,差異有統計學意義(t=15.356,P<0.05),見圖4。
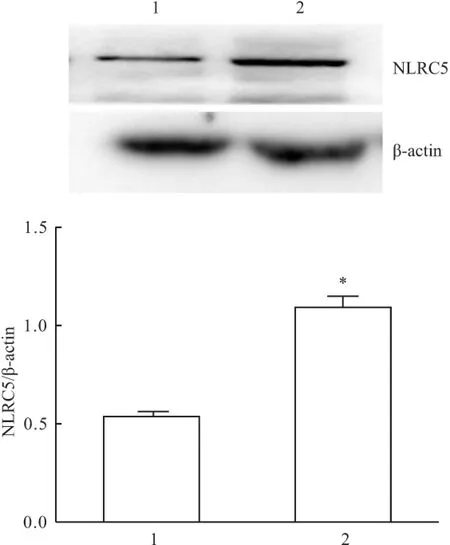
圖4 Western blot法檢測NLRC5蛋白表達
1:空質粒轉染組;2:穩定表達組;與空質粒轉染組比較:*P<0.05
3 討論
NLRs是細胞內模式識別受體家族,該家族包括5個亞家族,其中在人類至少發現了23個成員,在鼠中至少發現了34個成員。 這些成員通常含有3個結構區域: 碳端為亮氨酸重復區域,能夠識別相應的配體;中部區域為中心核酸結合結構域,在NLRs與核苷酸鏈的結合以及自我聚合方面具有重要作用[6]。NLRs的氨基端為半胱天冬酶激活招募結構域、熱蛋白結構域,能夠結合下游效應分子從而激活NLRs,故這個區域稱為功能結構域。近期新發現的NLRC5是NLRs家族中的一員,其基因定位于人類基因組的16q13,基因全長約96 000 bp[7],編碼蛋白由1 866個氨基酸分子組成,是所有NLRs家族中分子量最大的一個成員。研究[8]顯示,NLRC5在人類及小鼠的多種組織中都能表達,造血細胞中具有較高的表達量,人體和小鼠骨髓、淋巴結、胸腺和脾這些免疫組織具有高表達量的mRNA[8-10]。
NLRC5被證明能夠參與到多種免疫反應,但其在免疫調節過程中具體扮演什么角色還具有爭議性。Cui et al[11]研究發現NLRC5負向調節NF-κB途徑以及I型干擾素信號發生途徑。Tong et al[12]發現NLRC5基因缺陷性小鼠在受到水皰性口炎病毒感染后,體內多種細胞內的NF-κB以及I型干擾素信號途徑得到了加強。Neerincx et al[9]發現切除NLRC5基因的人單核細胞(THP-1)以及人真皮成纖維細胞受到Sendai病毒及聚胞嘧啶核苷酸感染后,胞內I型干擾素表達量會降低,認為NLRC5是I型干擾素產生途徑中的一個正向調節子。最近研究[5]顯示NLRC5在LX-2細胞中正向調節NF-κB/Smad3通路介導IL-6和IL-1β的分泌。有關NLRC5在免疫反應中的作用的研究很多,但其在炎癥小體的形成過程中所起的激活作用尚不明確。然而慢性炎癥終歸是導致癌癥的主要原因之一,發展免疫療法也成為治療癌癥的理想靶點分子[13]。
脂質體瞬時轉染中,外源性基因片段整合到基因組的概率非常低,主要以染色體外的形式存在,并且不能隨細胞分裂一同復制,常用于24~72 h內就進行處理分析結果的實驗中。而穩定轉染相對于瞬時轉染而言,其進入細胞的質粒能整合入基因組中,隨細胞的分裂一同復制。但穩定轉染并不是不同于瞬時轉染的方法,只是對瞬時轉染的細胞進行篩選,得到穩定整合的細胞株。通過構建穩定表達NLRC5的細胞株可以大大降低轉染該基因的頻率,降低成本,方便實驗研究。并且在后續動物實驗中,需要向動物體內注射表達有外源片段的細胞,需要構建穩定表達細胞株,以防止注射入動物體內后,外援片段很快丟失。
[1] Kudo M. Molecular targeted therapy for hepatocellular carcinoma:bench to bedside [J].Dig Dis,2011,29(3):273-7.
[2] Zhang D Y,Friedman S L.Fibrosis-dependent mechanisms of hepatocarcinogenesis [J].Hepatology, 2012, 569(2):769-75.
[3] He G,Karin M.NF-κB and STAT3-key players in liver inflammation and cancer [J].Cell Res,2011,21(1):159-68.
[4] 陳亞冰, 蘭道亮, 湯 程, 等.NLRC5:一種新發現的天然免疫調節分子[J].中國獸醫科學,2013,43(4):436-40.
[5] Xu T,Ni M M,Huang C,et al.NLRC5 mediates IL-6 and IL-1beta secretion in LX-2 cells and modulated by the NF-κB/Smad3 pathway [J].Inflammation,2015,38(5):1794-1804.
[6] Magalhaes J G, Sorbara M T, Girardin S E,et al. What is new with Nods? [J].Curr Opin Immunol,2011,23(1):29-34.
[7] Kuenzel S, Till A, Winkler M, et al. The nucleotide-binding oligomerizationdomain-like receptor NLRC5 is involved in IFN-dependent antiviral immune responses [J].J Immunol,2010,184(4):1990-2000.
[8] Benko S, Magalhaes J G, Philpott D J, et al. NLRC5 limits the activation of inflammatory pathways [J].J Immunol, 2010, 185(3):1681-91.
[9] Neerincx A, Lautz K, Menning M, et al. A role for the human nucleotide-binding domain, leucine-rich repeat-containing family member NLRC5 in antiviral responses [J].J Biol Chem,2010,285(34):26223-32.
[10] Davis B K, Roberts R A, Huang M T, et al. Cutting edge:NLRC5-dependent activation of the inflammasome [J].J Immunol,2011,186(3):1333-7.
[11] Cui J, Zhu L,Xia X, et al.NLRC5 negatively regulates the NF-kappaB andtype I interferon signaling pathways [J].Cell, 2010,141(3):483-96.
[12] Tong Y,Cui J,Li Q,et al.Enhanced TLR-induced NF-κB signaling and type I interferon responses in NLRC5 deficient mice [J].Cell Res,2012,22(5):822-35.
[13] Oldstone M B.Virus-lymphoid cell interactions[J].Proc Natl Acad Sci USA, 2012,93(23):12756-8.
Construction of stable a human hepatoma cell line HepG2 with over-expression of NLRC5
He Yinghua, Xu Tao, Ni Mingming, et al
(School of Pharmacy,Anhui Medical University,Hefei 230032)
Objective To establish a human hepatoma cell line HepG2 with over-expression of NLRC5. Methods Identified the suitable G418 concentration in the selection of stable transfection HepG2 cell. The eukaryotic expression plasmid pEGFP-C2-NLRC5 was transfected into the HepG2 cells mediated by lipofectamine and then selected with G418. Immunofluorescence assay and Western blot were used to analyze the expression of NLRC5. Results HepG2 cells were killed by G418 after 14 days with the minimum concentration of 300 ng/μl. Drug-resistant clones were formed after 14 days by selecting with G418. In the protein level, pEGFP-C2-NLRC5 HepG2 cell line was higher expression than pEGFP-C2 HepG2 cell line, the difference was statistically significant. Conclusion A hepatoma stable cell line over-expressing NLRC5 is successfully established, which may provide critical foundation for functional research of NLRC5 in liver cancer.
NLRC5; liver cancer; stable cell line; HepG2
時間:2015-12-30 14:38
http://www.cnki.net/kcms/detail/34.1065.R.20151230.1438.004.html
國家自然科學基金(編號:81473268、81202978);安徽省自然科學基金(編號:1308085MH145)
1安徽醫科大學藥學院,合肥 230032
2合肥瑤海區長淮街道社區衛生服務中心,合肥 230011
何鶯華,女,碩士研究生;
李 俊,男,教授,博士生導師,責任作者,E-mail: lj@ahmu.edu.cn
R 735.7
A
1000-1492(2016)01-0005-04
2015-10-20接收
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
