生長分化因子-9下調對小鼠卵巢顆粒細胞增殖能力的影響
2016-11-28 03:49:32馬會明張永芳王蒙蒙李昀伽王燕蓉裴秀英
安徽醫科大學學報 2016年1期
馬會明,張永芳,王蒙蒙,李昀伽,蒲 靜,于 佳,王燕蓉,裴秀英,徐 仙
?
生長分化因子-9下調對小鼠卵巢顆粒細胞增殖能力的影響
馬會明,張永芳,王蒙蒙,李昀伽,蒲 靜,于 佳,王燕蓉,裴秀英,徐 仙
目的 探討生長分化因子-9(GDF9)對小鼠卵巢顆粒細胞增殖能力基因周期蛋白A(cyclin A)、周期蛋白D1(cyclin D1)和周期蛋白依賴性激酶抑制因子27(p27kip1)表達的影響。方法 利用CCK-8檢測法檢測細胞的增殖能力;使用流式細胞術、Real-time PCR和Western blot法驗證GDF9 siRNA的干擾效率;采用Real-time PCR和Western blot法檢測細胞增殖cyclin A、cyclin D1和p27kip1 mRNA和蛋白表達情況。結果 GDF9 siRNA顯著抑制顆粒細胞增殖活力,其中72 h抑制最明顯(P<0.05);Real-time PCR和Western blot法檢測結果顯示GDF9 siRNA顯著下調小鼠卵巢顆粒細胞中GDF9 mRNA及蛋白的表達(P<0.01);Real-time PCR法檢測結果顯示,細胞cyclin A、cyclin D1 mRNA表達降低,p27kip1 mRNA表達增強;Western blot法結果顯示,GDF9 siRNA可促進細胞周期蛋白依賴性激酶抑制劑p27kip1的表達,同時下調G2/M期調控蛋白cyclin A、cyclin D1的表達。結論 靶向抑制小鼠卵巢顆粒細胞中GDF9的表達,下調細胞cyclin A、cyclin D1的表達,上調P27 kip1的表達,顯著抑制細胞的增殖。
GDF9 siRNA;細胞增殖;細胞周期;卵巢顆粒細胞;小鼠
女性卵巢儲備功能減退原因可能與顆粒細胞一系列重要的信號分子和周期蛋白調控有關[1-2], 細胞周期蛋白和細胞周期蛋白依賴性激酶(cyclin-dependent kinases,CDK)抑制劑在調控細胞周期轉換過程中具有重要作用[3],其中細胞周期D1(cyclin D1)和細胞周期A(cyclin A)的首要功能主要是促進細胞增殖[4]。CDK抑制蛋白p27(p27kip1)是一種CDK抑制劑,通過抑制cyclin D/CDK4、cyclinE/CDK2 的活性,從而抑制細胞增殖活性[5-6],使細胞失去分裂能力。在卵母細胞的成熟過程中GDF9作為一個旁分泌因子參與調節卵丘顆粒細胞擴展的過程[7]。該研究通過GDF9基因沉默對小鼠顆粒細胞中cyclin D1、cyclin A和p27kip1表達的檢測,旨在探討小鼠顆粒細胞細胞周期調控蛋白在細胞增殖分化機制中的作用,為卵母細胞發育機制提供一定理論依據。
1 材料與方法
1.1 主要試劑和儀器 逆轉錄試劑盒和熒光定量PCR(Real-time PCR,RT-PCR)試劑盒均購自日本TaKaRa 公司;實時熒光定量PCR (Real-time PCR) 試劑盒、TRIzol、DMEM/F12(1 ∶1)、Ⅰ型膠原酶、胎牛血清均購自美國Life Tech公司;兔來源cyclin A、cyclin D1及p27kip1單克隆抗體購自英國Abcam公司;β-actin兔單克隆抗體購自美國Santa Cruz公司;羊抗兔二抗及ECL化學發光試劑盒購自中國博士德生物公司;蛋白提取試劑盒購自南京凱基生物試劑公司;重組pcDNA 6.2-GDF9 siRNA(簡稱GDF9 siRNA)質粒、siRNA轉染專用試劑HiPerFect購自美國Qiagen公司; 680型酶標儀購自美國Bio-rad儀器公司;FACSCalibur型流式細胞儀購自美國BD公司;普通PCR儀和GelDoc XR System凝膠成像系統購自美國Bio-rad儀器公司;Real-time PCR儀購自美國ABI公司;BX51熒光倒置顯微鏡購自日本OLYMPUS公司。
1.2 小鼠卵泡顆粒細胞的獲得 取18~22 g 未成熟的雌性Bal小鼠30只(寧夏醫科大學實驗動物中心),按照常規方法[8]每只腹腔注射孕馬血清(PMSG) 5 IU,注射42 h后在無菌條件下迅速剖取卵巢放入無菌的PBS 中,去除卵巢周圍組織及表面包膜,PBS清洗后置于TCM199-Hepes培養基中。用不銹鋼針刺破卵巢表面的3~8 mm卵泡, 使顆粒細胞釋放于TCM199-Hepes培養基中。1 000 r/min 低速離心5 min,洗滌2次。將生長培養基TCM199 (含100 IU/ml雙抗、10% FBS、10 ng/ml EGF) 加入細胞團并制成單細胞懸液。將細胞接種在 6 孔培養板中,在5% CO2、37.5 ℃條件下孵育24 h。鏡下觀察是否污染及細胞的生長情況, 去除未貼壁細胞。此后隔日換液1次。所有動物實驗均遵循實驗動物的倫理要求和實驗動物管理條例。
1.3 顆粒細胞轉染及最適條件確定 小鼠顆粒細胞貼壁24 h后,開始轉染,按照質粒GDF9 siRNA DNA量和HiPerFect量為1.5 μg和12 μg設計實驗;每組設置3個復孔,轉染后48 h換生長培養基TCM199。在熒光顯微鏡下觀察轉染情況,用PBS充分洗滌去除漂浮的死細胞,用流式細胞儀分析轉染率,確定最佳轉染時間。轉染率(%)=綠色熒光細胞/細胞總數×100%。轉染設計分為空質粒siRNA control組和blank control組及GDF9 siRNA組。
1.4 小鼠顆粒細胞增殖情況的確定 按照常規方法[8]將分離純化的小鼠顆粒細胞接種至96孔培養板,使每孔細胞密度為1×104個,按照實驗設計,每組設3個平行孔,并設不含細胞的空白對照,37 ℃培養24 h使細胞貼壁。然后根據上述方法開始轉染GDF9 siRNA及對照siRNA,轉染48 h后,按照CCK-8試劑盒使用說明每孔加入10 μl CCK-8試劑,繼續培養4 h,檢測波長為450 nm下的光密度(optical density,OD)值。
1.5 反轉錄PCR(RT-PCR)和Real-time PCR法檢測轉染細胞中mRNA的表達 細胞轉染后48 h,清洗、消化細胞后按照常規方法(9)分別提取總RNA,并用Oligo(dT)反轉錄得到cDNA。利用GDF9、cyclin A、cyclin D1和p27kip1 mRNA特異性引物(表1),擴增特異性條帶。RT-PCR擴增反應結束后用2%的瓊脂糖凝膠檢測擴增的條帶大小;Real-time PCR反應以β-actin mRNA的表達作為內參,每組平行試驗重復3次,計算基因的相對表達量并用2-ΔΔCt法表示,各組的Ct值均經β-actin校正,以正常組的表達作為100%。
1.6 Western blot 法檢測轉染細胞中蛋白表達 細胞轉染后48 h,清洗、消化細胞并裂解細胞全蛋白,按照常規方法[9]用BCA法測定濃度并定量,取 50 μg 蛋白樣品按照常規方法進行聚丙烯酰胺凝膠電泳, 并濕轉移至PVDF膜上,封閉液封閉后,分別加入一抗cyclinA、cyclinD1和p27kip1 (1 000 ∶1),4 ℃孵育過夜;漂洗后加入相應的辣根過氧化物酶標記的二抗 (2 000 ∶1),37 ℃室溫孵育1 h,增強化學發光顯色系統ECL顯色,曝光。以β-actin作為等量蛋白質上樣對照,每個樣本至少重復3 遍,曝光圖片經GelDoc XR System凝膠成像系統掃描后,計算條帶的吸光度,cyclin A、cyclin D1和p27kip1 蛋白相對表達采用與β-actin吸光度比值來表示。

表1 Real-time PCR相關基因的引物序列及參數
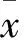
2 結果
2.1 GDF9 siRNA對小鼠卵巢顆粒細胞干擾后干擾效率的檢測 分離純化的小鼠卵巢顆粒細胞用GDF9 siRNA和無關序列siRNA control分別轉染細胞,48 h 后更換為正常培養基TCM199,熒光顯微鏡下觀察(圖1A),經流式細胞儀分析GDF9 siRNA和siRNA control兩組細胞,約有70%以上卵巢顆粒細胞已經表達綠色熒光蛋白(圖1B、1C),能滿足后續表達調控分析的要求。GDF9 siRNA和siRNA control轉染細胞48 h 后,提取總RNA,反轉錄后采用Real-time PCR檢測GDF9 mRNA的表達。結果表明,GDF9 siRNA轉染細胞GDF9 mRNA表達顯著下調(F=33.058,P<0.05),siRNA control轉染細胞GDF9 mRNA表達沒有顯著變化(圖1D)。

圖1 GDF9 siRNA轉染顆粒細胞轉染效率分析
A:顆粒細胞原代轉染×100;B:siRNA control轉染效率;C:GDF9 siRNA轉染效率;D:GDF9轉染后基因沉默檢測;與siRNA control和blank control比較:*P<0.05
2.2 siRNA干擾顆粒細胞GDF9基因表達對細胞增殖的影響 顆粒細胞轉染后,顯微鏡下觀察發現GDF9 siRNA顆粒細胞開始表現為細胞輪廓稍有萎縮,細胞漂浮物變多。CCK-8法檢測結果顯示:siRNA control與blank control相比,siRNA control轉染后48 h的OD值降低,GDF9 siRNA與siRNA control和blank control組之間差異有統計學意義(F=5.321,P<0.05)。見圖2。
2.3 siRNA干擾顆粒細胞GDF9對cyclin A、cyclin D1和p27kip1 mRNA表達的影響 GDF9 siRNA轉染細胞后, RT-PCR分別擴增出特異性cyclin A、cyclin D1和p27kip1片段,瓊脂糖凝膠電泳結果如圖3A,擴增的條帶大小與理論片段大小一致;Real-time PCR分析檢測cyclin A、cyclin D1和p27kip1基因表達情況 (圖3B)。siRNA control與blank control組相比較,cyclin A、cyclin D1和p27kip1基因表達無顯著性差異。干擾GDF9后p27kip1的mRNA表達量顯著上升 (F=259.165,P<0.05);干擾GDF9后cyclin D1的mRNA表達顯著下降(F=17.224,P<0.05);cyclin A的mRNA表達量降低,但沒有顯著性變化。

圖2 CCK-8法檢測GDF9 siRNA 對細胞增殖的影響

圖3 GDF9 siRNA 對p27 kip1、cyclin D1及cyclin A mRNA表達的影響
A:瓊脂糖凝膠電泳擴增結果;B:Real-time PCR分析結果;1: cyclin A; 2: cyclin D1; 3: p27kip1;與blank control組比較:*P<0.05
2.4 siRNA干擾顆粒細胞GDF9對cyclin A、cyclin D1和p27kip1 蛋白表達的影響 Western blot檢測顯示,干擾GDF9后,可顯著增加p27kip1蛋白的表達(F=123.564,P<0.05),cyclin D1蛋白的表達明顯下降(F=11.237,P<0.05)。但干擾GDF9 后,cyclin A蛋白表達下降,但是沒有顯著性改變。見圖4。

圖4 GDF9 siRNA 對p27kip1、cyclin D1及cyclin A 蛋白表達的影響
1:blank control組; 2:siRNA control組; 3:GDF9 siRNA組;與blank control組比較:*P<0.05
3 討論
在正常卵巢的周期性卵泡發育過程中, 卵泡發育與顆粒細胞增殖分化和凋亡密切相關。GDF9對卵泡發育有重要作用的細胞因子,可以通過多個信號通路促進顆粒細胞增殖過程[7]。本研究中利用RNA干擾技術沉默GDF9基因的表達,達到抑制顆粒細胞增殖,首先發現靶向GDF9的siRNA對顆粒細胞干擾效果明顯,GDF9基因表達量明顯降低,能夠通過細胞周期蛋白表達研究卵巢顆粒細胞增殖,為研究PCOS患者卵巢顆粒細胞黃素化和凋亡的機制提供實驗依據,為研究細胞周期調控的相關信號通路奠定基礎。
細胞周期是與細胞生長增殖極其相關的一系列細胞內反應過程,在這一重要過程中有許多關鍵的因子調控細胞的進程,這些分子包括cyclin D1、cyclin A和p27kip1等,正常的細胞周期有序的循環是通過激活和失活CDK來實現的。cyclinD1作為細胞周期進程的啟動子,是調控細胞周期G1期的關鍵蛋白,能夠促進細胞的生長增殖。cyclin A在細胞周期中的G1-S和G2-M時相中均發揮了作用,能夠以有活性的cyclin A-CDK-2復合物的形式促進細胞的增殖[10]。本研究結果顯示,沉默GDF9基因的表達后,cyclin D1的mRNA和蛋白表達顯著下降,導致細胞周期進程減慢,抑制細胞生長增殖。其中,可以與CDK4結合形成復合體,通過磷酸化和失活視網膜母細胞瘤蛋白(RB),促進細胞周期從G1期到S期的進展,并通過這種方式介導DNA的合成,是一個比其他Cyclins更加敏感的指標,對細胞周期調控至關重要[10]。cyclin A可在細胞周期的不同時相分別與 CDK-1、CDK-2結合,并激活其活性[11]。可以磷酸化RB蛋白而使E2F游離,活化、細胞便進入增殖階段[12]。Viglietto et al[13]研究結果顯示,cyclin A的表達量有一定的降低,但是沒有顯著性下降,p27kip1可以與cyclin E-CDK2、cyclin A-CDK2、cyclin D-CDK4復合物結合抑制其對底物的磷酸化作用,使細胞不能發生G1~S期的時相轉換,停滯于G1期,細胞增殖停止。本研究結果顯示,干擾GDF9后, p27kip1的mRNA和蛋白表達量顯著上升,細胞增殖減緩,因此也證實了p27kip1在介導細胞外信號,尤其在細胞增殖及其調控中扮演負性調節的角色[14],可能p27kip1在細胞PI3K 和 Notch通路中參與細胞增殖和細胞周期調控[15], 本研究也證實了p27kip1表達上升促進了對cyclin D1的抑制作用,使cyclin D1表達顯著性下降,從而抑制了細胞的增殖分裂。但是研究中是否存在PI3K/Akt/p27kip1信號轉導通路,顆粒細胞PI3K通路活性是否受到影響,從而影響細胞周期的正常調控,需要進一步對顆粒細胞進行研究。
[1] 曹云霞.高齡女性生育力的變化與評估[J].中國計劃生育和婦產科,2014,6(8):8-12,23.
[2] 陳蓓麗, 曹云霞. 卵巢早衰的病因學研究進展[J].國際生殖健康/計劃生育雜志,2013,32(5):339-43.
[3] Foijer F, te Riele H. Check, double check: the G2 barrier to cancer[J].Cell Cycle,2006,5(8):831-6.
[4] Wang X, Meng X, Sun X, et al. Wnt/beta-catenin signaling pathway may regulate cell cycle and expression of cyclin A and cyclin E protein in hepatocellular carcinoma cells[J]. Cell Cycle, 2009,8(10):1567-70.
[5] Shi Z, Ji D F, Li Y G, et al.Activation of P27kip1-cyclin D1/E-CDK2 pathway by polysaccharide from Phellinus linteus leads to S-phase arrest in HT-29 cells[J]. Chem Biol Interact, 2013, 206(2):222-9.
[6] 黃 濤, 張令達.口腔鱗狀細胞癌組織中p57~(kip2)和Cyclin D1 mRNA的表達及意義[J]. 安徽醫科大學學報,2013, 48(4):398-401.
[7] 常秀峰, 張 敏, 吳小華,等. 卵母細胞分泌的生長分化因子-9和骨形成蛋白-15與卵母細胞發育潛能的關系[J].生殖與避孕,2014, 34(3):200-4.
[8] 馬會明, 黑常春, 張永芳,等. EGF通過PI3K/AKT通路介導小鼠卵巢顆粒細胞增殖[J].寧夏醫科大學學報, 2014,36(5):483-6+90.
[9] 張永芳, 王蒙蒙, 李 昕,等. 二甲雙胍通過抑制核糖體40S小亞基S6K蛋白激酶(P70S6)調節體外人顆粒細胞胰島素受體底物-1(IRS-1)的表達[J].生殖與避孕. 2015,35(3):146-53.
[10] Ji Y R, Kim H J, Park S J, et al. Critical role of Rgs19 in mouse embryonic stem cell proliferation and differentiation[J]. Differentiation,2015,89(1-2):42-50.
[11] Yang T Y, Chang G C, Chen K C, et al. Sustained activation of ERK and Cdk2/cyclin-A signaling pathway by pemetrexed leading to S-phase arrest and apoptosis in human non-small cell lung cancer A549 cells[J].Eur J Pharmacol,663(1-3):17-26.
[12] Nougarede R, Della Seta F, Zarzov P, et al. Hierarchy of S-phase-promoting factors: yeast Dbf4-Cdc7 kinase requires prior S-phase cyclin-dependent kinase activation[J]. Mol Cell Biol,2000,20(11):3795-806.
[13] Viglietto G, Motti M L, Fusco A.Understanding p27(kip1) deregulation in cancer: down-regulation or mislocalization[J]. Cell Cycle,2002,1(6):394-400.
[14] Zhang W, Nie Y, Chong L, et al. PI3K and Notch signal pathways coordinately regulate the activation and proliferation of T lymphocytes in asthma[J].Life Sci,92(17-19):890-5.
[15] Jian Q, Tao Z, Li Y, et al. Acute retinal injury and the relationship between nerve growth factor, Notch1 transcription and short-lived dedifferentiation transient changes of mammalian Muller cells[J]. Vision Res, 2015,110(Pt A):107-17.
Effect of downregulated growth differential factor-9 on cell proliferative activity in mouse ovarian granulose cell
Ma Huiming,Zhang Yongfang,Wang Mengmeng,et al
(Key Laboratory of Fertility Preservation and Maintenance of Ministry of Education,FertilityCenterforReproductiveMedicine,DeptofAnotomyandHistology-Embryology,NingxiaMedicalUniversity,yinchuan750004)
Objective In the present study, we determined whether GDF9 siRNA were involved in the expressions of cyclin A, cyclin D1 and p27kip1 which were key molecules in controlling cell cycling and cell proliferation by regulating cyclin D1 expression in mouse ovary granulose cell. Methods Real-time PCR and Western blot assay were used to investigate the expressions of cyclin D1, cyclin A, and p27kip1.We further observed the effect of GDF9 siRNA on the cell proliferation by regulating the expressions of cyclin D1, cyclin A, and p27kip1 in mouse ovary granulose cell.Results We found that the expressions of cyclin D1 and cyclin A were down-regulated at both protein and mRNA levels in GDF9 siRNA group while p27kip1 was upregulated by real-time PCR and Western blot assay. Furthermore, GDF9 siRNA had additive effect in down-regulation of cyclin D1,cyclin A and upregulation of p27kip1.Compared with the control group,the expression level of cyclin D1 was significantly decreased(P<0.05)and p27kip1 was significantly increased(P<0.05).Conclusion These data suggest that cell cyclin signal pathway may coordinately regulate the cell proliferation and differentiation processes through up-regulating cyclin D1 and down-regulating p27kip1 of mouse ovary granulose cell.
GDF9 siRNA;cell proliferation;ovary granulose cell;mouse
時間:2015-12-30 14:38
http://www.cnki.net/kcms/detail/34.1065.R.20151230.1438.024.html
寧夏自然科學基金(編號:NZ11128)
寧夏醫科大學生育力保持教育部重點實驗室、寧夏回族自治區生殖與遺傳重點實驗室、人體解剖與組織胚胎學系,寧夏醫科大學總醫院生殖醫學中心,銀川 750004
馬會明,男,講師,博士;
裴秀英,女,教授,碩士生導師,責任作者,E-mail:peixiuying@163.com;
R 711.6
A
1000-1492(2016)01-0051-05
2015-09-30接收
徐 仙,男,主任醫師,教授,碩士生導師,責任作者,E-mail:xux36@163.com
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
