江西山藥莖尖包埋玻璃化法超低溫保存及其遺傳穩(wěn)定性檢測(cè)
2016-11-28 03:24:00尹明華徐志堅(jiān)章省琴呂思杰曾艷紅付有章夏瑾華洪森榮
浙江農(nóng)業(yè)學(xué)報(bào) 2016年6期
尹明華,徐志堅(jiān),章省琴,呂思杰,曾艷紅,付有章,夏瑾華,洪森榮
(上饒師范學(xué)院 生命科學(xué)學(xué)院,江西 上饒 334001)
?
江西山藥莖尖包埋玻璃化法超低溫保存及其遺傳穩(wěn)定性檢測(cè)
尹明華,徐志堅(jiān),章省琴,呂思杰,曾艷紅,付有章,夏瑾華,洪森榮*
(上饒師范學(xué)院 生命科學(xué)學(xué)院,江西 上饒 334001)
采用植物組織培養(yǎng)法對(duì)廣豐藥薯莖尖的包埋玻璃化法超低溫保存程序進(jìn)行優(yōu)化,并采用擴(kuò)增片段長(zhǎng)度多態(tài)性(AFLP)分子標(biāo)記法和流式細(xì)胞術(shù)對(duì)其凍后再生苗的遺傳穩(wěn)定性進(jìn)行檢測(cè),同時(shí)將超低溫程序應(yīng)用到江西山藥其他地方品種,旨在為薯蕷屬植物種質(zhì)資源的長(zhǎng)期保存奠定理論基礎(chǔ)。結(jié)果表明,廣豐藥薯莖尖預(yù)培養(yǎng)的較佳時(shí)間為5 d,較佳的蔗糖濃度為0.75 mol·L-1;裝載的較佳時(shí)間為40 min;脫水的較佳溫度為0 ℃,較佳時(shí)間為60 min;凍后黑暗培養(yǎng)7 d可以顯著提高其成活率。用AFLP分子標(biāo)記法和流式細(xì)胞術(shù)對(duì)莖尖凍后再生植株的遺傳穩(wěn)定性進(jìn)行檢測(cè),沒(méi)有發(fā)現(xiàn)異常條帶和染色體倍性變化,氣孔觀察也未發(fā)現(xiàn)葉下表皮氣孔參數(shù)的顯著變異。將這種超低溫保存程序用于江西山藥其他基因型,成活率約為40%~85%。該研究建立的包埋玻璃化法超低溫保存程序能保證江西山藥的遺傳穩(wěn)定性,可為建立江西山藥種質(zhì)資源超低溫保存庫(kù)提供一定的技術(shù)支撐。
江西山藥;莖尖;包埋玻璃化法;超低溫保存;遺傳穩(wěn)定性
山藥是薯蕷科(Dioscoreaceae)薯蕷屬(Dioscorea)一年生或多年生纏繞性藤本植物,能形成肥大的地下肉質(zhì)塊莖供為食用或藥用,營(yíng)養(yǎng)價(jià)值高[1]。江西是中國(guó)正品山藥的地道和主要產(chǎn)區(qū)之一,出產(chǎn)的山藥品質(zhì)優(yōu)良,在國(guó)內(nèi)外享有盛譽(yù)。江西種植的山藥品種繁多,且大多為江西傳統(tǒng)名優(yōu)地方品種,如南城縣的南城淮山、瑞昌的瑞昌山藥、廣豐的千金薯和廣豐藥薯、泰和竹篙薯等[2-9]。這些山藥有些已成為產(chǎn)區(qū)的地理標(biāo)志和品牌性種質(zhì)資源,其品質(zhì)和營(yíng)養(yǎng)成分與懷山藥具有顯著的差異[10]。目前,江西山藥種質(zhì)資源的繁殖和保存主要采取苗床集中悶種、出苗即栽的方法每年進(jìn)行田間種植,但由于江西山藥地方品種因常年在同一地區(qū)甚至同一地塊上連作和農(nóng)民缺乏提純復(fù)壯技術(shù)[2],品種染病和感病害率較高,品種退化嚴(yán)重,病蟲害抗性降低,產(chǎn)量較低,而且傳統(tǒng)搭架栽培模式不僅費(fèi)工、費(fèi)時(shí)、費(fèi)力,而且容易受到自然災(zāi)害(如病蟲害、冰雪、地震等)的影響[9]。因此,研究江西山藥地方品種種質(zhì)資源的超低溫保存具有重要的意義。
超低溫保存(cryopreservation)是指將植物材料活體,采取一定的技術(shù),安全存入-196 ℃液氮中長(zhǎng)期保存,需要時(shí)采取一定的方法使之回到常溫并正常生長(zhǎng)的一整套生物技術(shù)[11]。超低溫保存是目前長(zhǎng)期保存植物種質(zhì)資源最有效的方法,已廣泛應(yīng)用于數(shù)百種植物[12]。包埋玻璃化超低溫保存是20世紀(jì)90年代中期興起的一種新的超低溫保存技術(shù),具有毒害性小、設(shè)備簡(jiǎn)單、易于操作、恢復(fù)生長(zhǎng)快等優(yōu)點(diǎn)。目前,關(guān)于懷山藥超低溫保存的研究在國(guó)內(nèi)外多有報(bào)道[13-17],但關(guān)于江西山藥地方品種種質(zhì)資源的超低溫保存尚未見報(bào)道。本文先以廣豐藥薯為研究對(duì)象,探討和優(yōu)化其包埋玻璃化法超低溫保存程序,然后將這種程序應(yīng)用于江西山藥其他地方品種,以期為建立江西山藥特異種質(zhì)資源超低溫庫(kù)提供科學(xué)依據(jù)和一定的技術(shù)支撐。
1 材料與方法
1.1 材料
廣豐藥薯、廣豐千金薯、瑞昌山藥、南城淮山和泰和竹篙薯試管苗均由上饒師范學(xué)院生命科學(xué)學(xué)院植物組織培養(yǎng)室提供。
1.2 方法
1.2.1 廣豐藥薯莖尖的切取與包埋
將廣豐藥薯試管苗(培養(yǎng)60 d)的腋芽在10~20倍的雙目解剖鏡下切取約1~2 mm莖尖(含有3~4葉原基)。(25±1) ℃下將莖尖移入含3%海藻酸鈉和0.4 mol·L-1蔗糖的無(wú)鈣離子改良MS培養(yǎng)基中,注射器吸取莖尖混合物滴入含0.1 mol·L-1CaCl2和0.4 mol·L-1蔗糖的溶液中停留30 min,以充分形成直徑約4 mm的球形包埋珠,每包埋珠含有1個(gè)莖尖。
1.2.2 廣豐藥薯莖尖包埋珠的預(yù)培養(yǎng)
廣豐藥薯莖尖包埋珠浸泡于附加有0~1 mol·L-1蔗糖的MS+2 mg·L-1KT+0.5 mg·L-1NAA培養(yǎng)基中,14 h (L)/10 h (D)光周期下預(yù)培養(yǎng)5 d;或廣豐藥薯莖尖包埋珠浸泡于附加有0.75 mol·L-1蔗糖的MS+2 mg·L-1KT+0.5 mg·L-1NAA培養(yǎng)基中,置于14 h (L)/10 h (D)光周期下預(yù)培養(yǎng)0~7 d,然后將廣豐藥薯莖尖包埋珠放入含有2 mol·L-1甘油和0.4 mol·L-1蔗糖的MS裝載液,(25±1)℃處理40 min,在0 ℃再用PVS2[18](30%甘油+15%二甲基亞砜+15%乙二醇+0.4 mol·L-1蔗糖)脫水60 min后直接投入液氮保存。保存24 h后,包埋珠在37 ℃水浴中化凍3 min,然后用MS+2 mg·L-1KT+0.5 mg·L-1NAA+1.2 mol·L-1蔗糖的溶液洗滌3次,每次約10 min。去除海藻酸鈣包埋物,將包埋珠的莖尖剝離出來(lái),轉(zhuǎn)入MS+2 mg·L-1KT+0.5 mg·L-1NAA固體培養(yǎng)基上,暗培養(yǎng)7 d后轉(zhuǎn)入14 h (L)/10 h (D)光周期中進(jìn)行恢復(fù)和再生。以莖尖長(zhǎng)出新芽為莖尖成活的標(biāo)志,試驗(yàn)重復(fù)3次,統(tǒng)計(jì)廣豐藥薯莖尖的平均成活率,成活率(%)=(玻璃化超低溫保存后成活的莖尖數(shù)/保存的總莖尖數(shù))×100,篩選出廣豐藥薯莖尖包埋珠預(yù)培養(yǎng)的最佳蔗糖濃度和預(yù)培養(yǎng)時(shí)間。
1.2.3 廣豐藥薯莖尖包埋珠的裝載
廣豐藥薯莖尖包埋珠浸泡于MS+2 mg·L-1KT+0.5 mg·L-1NAA+0.75 mol·L-1蔗糖的培養(yǎng)基中,置于14 h (L)/10 h (D)光周期下預(yù)培養(yǎng)5 d,然后將廣豐藥薯莖尖包埋珠放入含有2 mol·L-1甘油和0.4 mol·L-1蔗糖的MS裝載液,(25±1)℃處理0~50 min。然后包埋珠經(jīng)過(guò)脫水、液氮保存、化凍和洗滌后,將包埋珠的莖尖剝離出來(lái),轉(zhuǎn)入MS+2 mg·L-1KT+0.5 mg·L-1NAA固體培養(yǎng)基上,暗培養(yǎng)7 d后轉(zhuǎn)入14 h (L)/10 h (D)光周期中進(jìn)行恢復(fù)和再生。試驗(yàn)重復(fù)3次,統(tǒng)計(jì)廣豐藥薯莖尖的平均成活率,篩選出廣豐藥薯莖尖包埋珠的最佳裝載時(shí)間。
1.2.4 廣豐藥薯莖尖包埋珠的脫水
廣豐藥薯莖尖包埋珠浸泡于MS+2 mg·L-1KT+0.5 mg·L-1NAA+0.75 mol·L-1蔗糖的培養(yǎng)基中,置于14 h (L)/10 h (D)光周期下預(yù)培養(yǎng)5 d,然后將廣豐藥薯莖尖包埋珠放入含有2 mol·L-1甘油和0.4 mol·L-1蔗糖的MS裝載液,(25±1)℃處理40 min后,0 ℃下用PVS2脫水0~80 min,或25 ℃用PVS2脫水0~60 min后直接投入液氮保存。然后包埋珠經(jīng)過(guò)化凍和洗滌后,將包埋珠的莖尖剝離出來(lái),轉(zhuǎn)入MS+2 mg·L-1KT+0.5 mg·L-1NAA固體培養(yǎng)基上,暗培養(yǎng)7 d后轉(zhuǎn)入14 h (L)/10 h (D)光周期中進(jìn)行恢復(fù)和再生。試驗(yàn)重復(fù)3次,統(tǒng)計(jì)廣豐藥薯莖尖的平均成活率,篩選出廣豐藥薯莖尖包埋珠脫水的最佳溫度和脫水時(shí)間。
1.2.5 廣豐藥薯莖尖包埋珠的凍后培養(yǎng)
廣豐藥薯莖尖包埋珠經(jīng)過(guò)預(yù)培養(yǎng)、裝載和脫水、液氮保存、化凍和洗滌后,將包埋珠的莖尖剝離出來(lái),轉(zhuǎn)入MS+2 mg·L-1KT+0.5 mg·L-1NAA固體培養(yǎng)基上。培養(yǎng)方式為2種:一種是正常的14 h (L)/10 h (D)光周期培養(yǎng),另一種是先暗培養(yǎng)7 d后轉(zhuǎn)入14 h (L)/10 h (D)光周期中培養(yǎng)。試驗(yàn)重復(fù)3次,統(tǒng)計(jì)廣豐藥薯莖尖的平均成活率,篩選出廣豐藥薯莖尖包埋珠的最佳凍后培養(yǎng)方式。
1.2.6 廣豐藥薯莖尖凍后再生植株的氣孔觀察
氣孔觀察參照陳佰鴻等[19]報(bào)道的透明膠帶粘取法。用透明膠帶粘取再生苗葉下表皮并制片,于顯微鏡下觀測(cè)氣孔大小(氣孔長(zhǎng)直徑、氣孔短直徑)、氣孔面積(氣孔面積=圓周率×長(zhǎng)半徑×短半徑)以及1 000 μm2氣孔數(shù)(取50個(gè)視野),拍照。
1.2.7 廣豐藥薯莖尖凍后再生植株的流式細(xì)胞儀檢測(cè)
流式細(xì)胞儀(flow cytometry,F(xiàn)CM)檢測(cè)按照尹明華等[20]建立的方法進(jìn)行。
1.2.8 廣豐藥薯莖尖凍后再生植株的AFLP檢測(cè)
按照郭文等[21]提供的AFLP程序?qū)V豐藥薯再生植株進(jìn)行AFLP分析,步驟如下:DNA樣品的制備、接頭制作、酶切—連接、預(yù)擴(kuò)增、選擇性擴(kuò)增和非變性膠檢測(cè)PCR結(jié)果。
1.2.9 江西山藥基因型對(duì)包埋玻璃化法超低溫保存的影響
將上述建立和優(yōu)化的廣豐藥薯莖尖包埋玻璃化法超低溫保存程序應(yīng)用到江西山藥其他基因型上,比較其超低溫保存效果的差異。廣豐千金薯、瑞昌山藥、南城淮山和泰和竹篙薯試管苗莖尖經(jīng)過(guò)包埋、預(yù)培養(yǎng)、裝載和脫水以及液氮保存后,去除海藻酸鈣包埋物,將莖尖轉(zhuǎn)入MS+2 mg·L-1KT+0.5 mg·L-1NAA固體培養(yǎng)基上,暗培養(yǎng)7 d后轉(zhuǎn)入14 h (L)/10 h (D)光周期中培養(yǎng)。試驗(yàn)重復(fù)3次,統(tǒng)計(jì)江西山藥各基因型莖尖的成活率,比較江西山藥基因型對(duì)超低溫保存的影響。
1.2.10 統(tǒng)計(jì)方法
所有數(shù)據(jù)表示為平均值±標(biāo)準(zhǔn)差,試驗(yàn)數(shù)據(jù)用SPSS 19.0軟件進(jìn)行One-way ANOVA分析,再進(jìn)行LSD法檢驗(yàn),P<0.05表示差異顯著。
2 結(jié)果與分析
2.1 預(yù)培養(yǎng)對(duì)廣豐藥薯莖尖包埋玻璃化法超低溫保存的影響
從圖1-A可知,預(yù)培養(yǎng)5 d時(shí),隨著蔗糖濃度的增大,莖尖的凍后成活率也顯著提高;蔗糖濃度為0.75 mol·L-1時(shí),莖尖的凍后成活率達(dá)到最大值;當(dāng)蔗糖濃度高于達(dá)到1 mol·L-1時(shí),莖尖的凍后成活率開始顯著下降。從圖1-B可知,蔗糖濃度為0.75 mol·L-1時(shí),預(yù)培養(yǎng)1和3 d,莖尖的凍后成活率無(wú)顯著差異,均顯著高于對(duì)照組(預(yù)培養(yǎng)時(shí)間為0 d);預(yù)培養(yǎng)5 d,莖尖的凍后成活率達(dá)到最高值,并與預(yù)培養(yǎng)1和3 d的成活率有極顯著差異;預(yù)培養(yǎng)7 d時(shí),莖尖的凍后成活率開始顯著下降。因此,MS+2 mg·L-1KT+0.5 mg·L-1NAA培養(yǎng)基中添加0.75 mol·L-1蔗糖,預(yù)培養(yǎng)5 d有助于莖尖的凍后成活。
2.2 裝載時(shí)間對(duì)廣豐藥薯莖尖包埋玻璃化法超低溫保存的影響
從圖2可知,含有2 mol·L-1甘油和0.4 mol·L-1蔗糖的MS裝載液對(duì)于莖尖的凍后成活有顯著作用。裝載時(shí)間為10和20 min時(shí),莖尖的凍后成活率均極顯著高于對(duì)照。裝載時(shí)間為30 min時(shí),莖尖的凍后成活率極顯著高于10和20 min。裝載時(shí)間為40 min時(shí),莖尖的凍后成活率達(dá)到最高。但裝載時(shí)間為50 min時(shí),莖尖的凍后成活率極顯著下降,下降至裝載時(shí)間為30 min的水平。
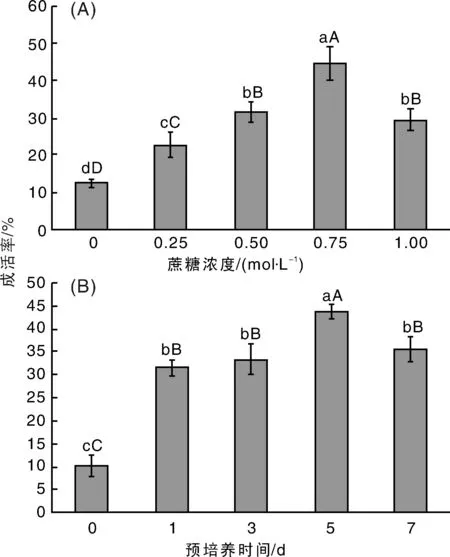
圖中不同柱子上無(wú)相同大小寫字母分別表示在0.01和0.05水平差異顯著。下同。圖1 預(yù)培養(yǎng)基的蔗糖濃度(A)和預(yù)培養(yǎng)時(shí)間(B)對(duì)廣豐藥薯莖尖包埋玻璃化法超低溫保存的影響Fig.1 Effect of sucrose concentration in preculture meidum (A) and preculture time (B) on cryopreservation of Guangfeng medicinal yam shoot-tip by encapsulation-vitrification

圖2 裝載時(shí)間對(duì)廣豐藥薯莖尖包埋玻璃化法超低溫保存的影響Fig.2 Effect of loading time on cryopreservation of Guangfeng medicinal yam shoot-tip by encapsulation-vitrification
2.3 PVS2脫水溫度和脫水時(shí)間對(duì)廣豐藥薯莖尖包埋玻璃化法超低溫保存的影響
從圖3可知,PVS2脫水溫度為0 ℃時(shí),莖尖在20~80 min的凍后成活率為32.8%~43.9%;脫水溫度為25 ℃時(shí),莖尖在10~60 min的凍后成活率為14.9%~37.2%,兩者差異顯著。脫水溫度為0 ℃時(shí),PVS脫水時(shí)間也會(huì)顯著影響莖尖的凍后成活,其中脫水60 min有助于莖尖的凍后成活。

圖3 PVS2脫水溫度和脫水時(shí)間對(duì)廣豐藥薯莖尖包埋玻璃化法超低溫保存的影響Fig.3 Effect of PVS2 dehydrationon temperature and dehydrationon time on cryopreservation of Guangfeng medicinal yam shoot-tip by encapsulation-vitrification
2.4 凍后培養(yǎng)方式對(duì)廣豐藥薯莖尖包埋玻璃化法超低溫保存的影響
14 h (L)/10 h (D)光周期培養(yǎng)和暗培養(yǎng)7 d后轉(zhuǎn)入14 h (L)/10 h (D)光周期培養(yǎng),成活率分別為32.9%±4.6%和43.7%±4.2%,差異極顯著。表明凍后一定時(shí)間的黑暗培養(yǎng)可以提高其凍后成活率,同時(shí)顯著減輕廣豐藥薯莖尖的褐化。
2.5 廣豐藥薯莖尖凍后再生植株的氣孔觀察
在FCM檢測(cè)之前,對(duì)廣豐藥薯常規(guī)繼代植株和莖尖凍后再生植株進(jìn)行氣孔觀察。結(jié)果表明,常規(guī)繼代植株和莖尖凍后再生植株的葉下表皮氣孔參數(shù)之間無(wú)顯著差異(表1,圖4),表明超低溫沒(méi)有造成廣豐藥薯再生植株的氣孔變化,推測(cè)不會(huì)影響到再生植株的光合生理。
表1 廣豐藥薯2種再生植株的葉下表皮氣孔參數(shù)比較
Table 1 Comparison of stomatal parameters of leaf lower epidermis of two kinds of regenerated plantlets of Guangfeng medicinal yam

植株來(lái)源氣孔長(zhǎng)直徑/μm氣孔短直徑/μm氣孔長(zhǎng)短直徑比氣孔面積/μm21000μm2氣孔數(shù)常規(guī)繼代植株3.9±0.22.3±0.31.70±0.3232.46±0.4242.8±3.8莖尖凍后再生植株3.7±0.92.1±0.41.76±0.2431.21±0.6743.9±2.9

A, 正常繼代植株;B, 莖尖凍后再生植株圖4 廣豐藥薯2種再生植株的氣孔Fig.4 Stomatal observation of two kinds of regenerated plantlets of Guangfeng medicinal yam
2.6 廣豐藥薯莖尖凍后再生植株的FCM檢測(cè)
利用FCM對(duì)廣豐藥薯的母株和莖尖凍后再生植株進(jìn)行倍性檢測(cè),DNA含量分布顯示,再生植株的倍性和對(duì)照母株相比,PI熒光值曲線相似,峰值相同,沒(méi)有發(fā)現(xiàn)染色體倍性的變化(圖5和圖6)。因此,廣豐藥薯莖尖凍后再生植株的倍性是穩(wěn)定的。
2.6 廣豐藥薯莖尖凍后再生植株的AFLP檢測(cè)
采用AFLP技術(shù)對(duì)廣豐藥薯常規(guī)繼代植株和莖尖凍后再生植株進(jìn)行檢測(cè)分析,5對(duì)引物(E-AAG/M-CAC,E-ACC/M-CTC,E-AGC/M-CTG,E-AGG/M-CAA,E-AGG/M-CTC)的擴(kuò)增出的總條帶數(shù)為41,平均每個(gè)引物獲得8.2個(gè)位點(diǎn),其中28個(gè)為可重復(fù)的多態(tài)性位點(diǎn),平均每個(gè)引物檢測(cè)到的多態(tài)性位點(diǎn)數(shù)為5.6個(gè),平均多態(tài)性位點(diǎn)百分率為68.29%。其中引物E-AAG/M-CAC的AFLP擴(kuò)增結(jié)果如圖7所示。結(jié)果表明,在廣豐藥薯常規(guī)繼代植株和莖尖凍后再生植株之間沒(méi)有發(fā)現(xiàn)變異位點(diǎn),說(shuō)明廣豐藥薯常規(guī)繼代植株和莖尖凍后再生植株之間基因組序列保持一致,無(wú)可檢測(cè)到的遺傳變異。
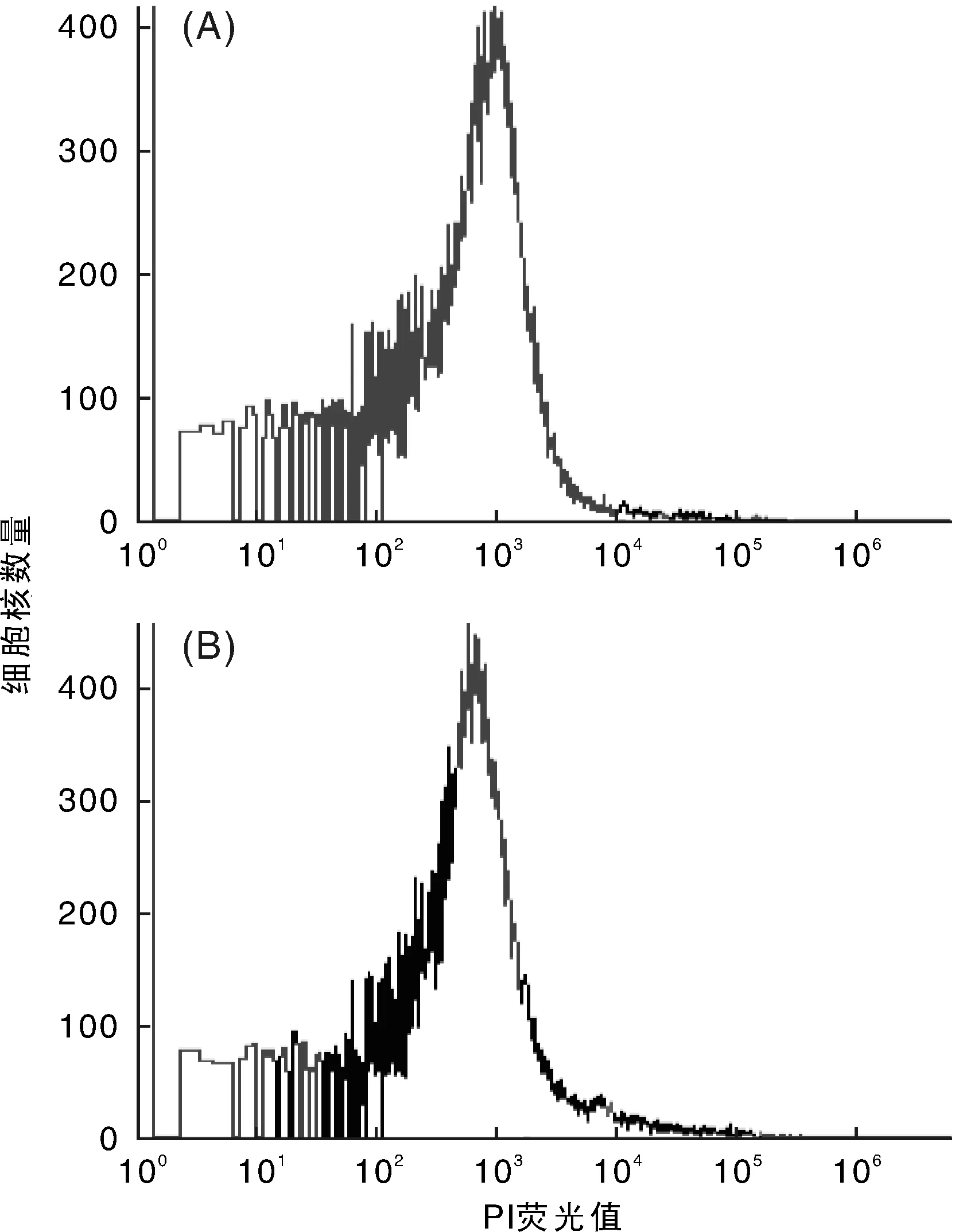
A, 廣豐藥薯常規(guī)繼代植株;B, 莖尖凍后再生植株圖5 廣豐藥薯FCM檢測(cè)的直方圖Fig.5 FCM histogram of Guangfeng medicinal yam
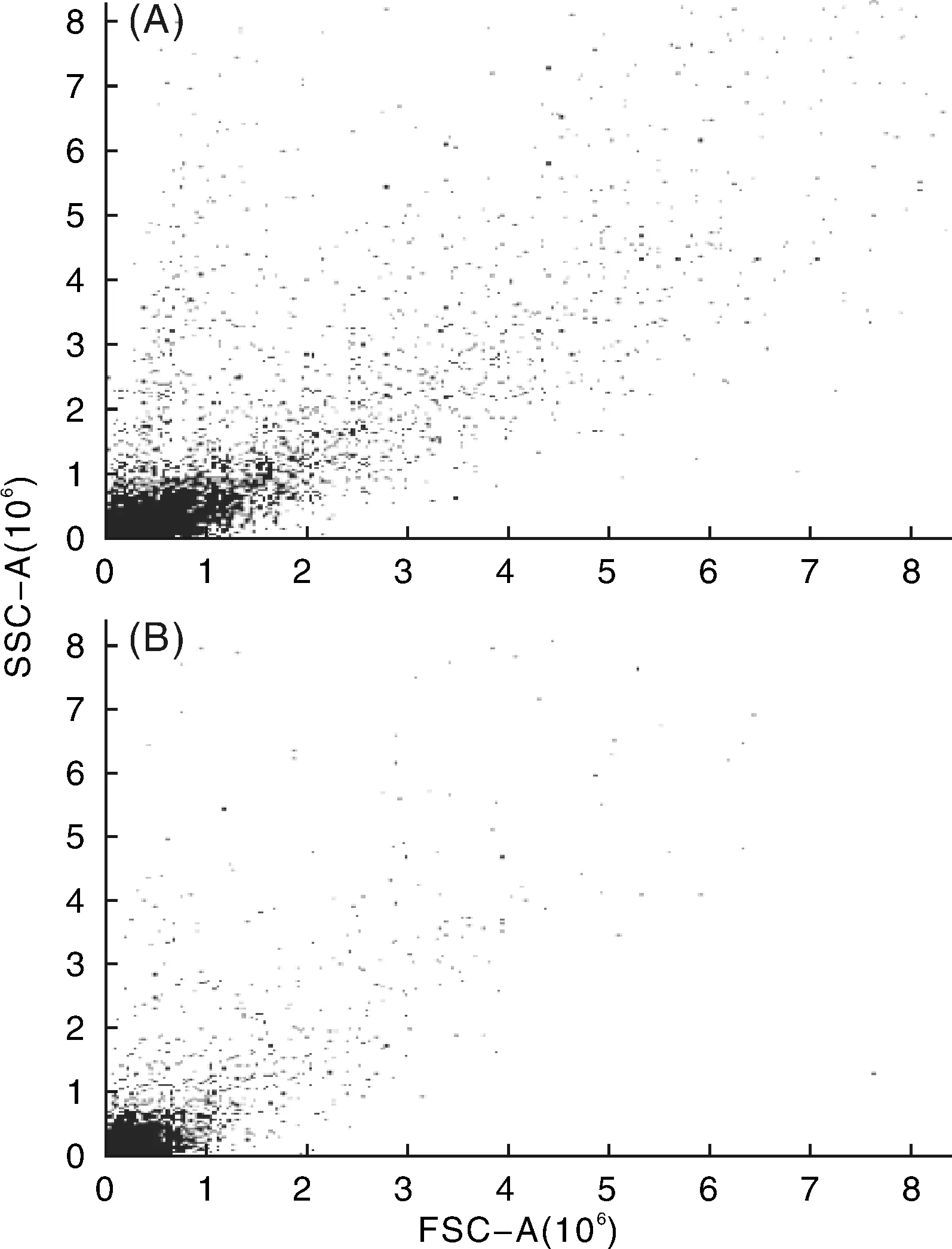
A, 常規(guī)繼代植株;B, 莖尖凍后再生植株圖6 廣豐藥薯FCM檢測(cè)的散點(diǎn)圖Fig.6 FCM scatter diagram of Guangfeng medicinal yam
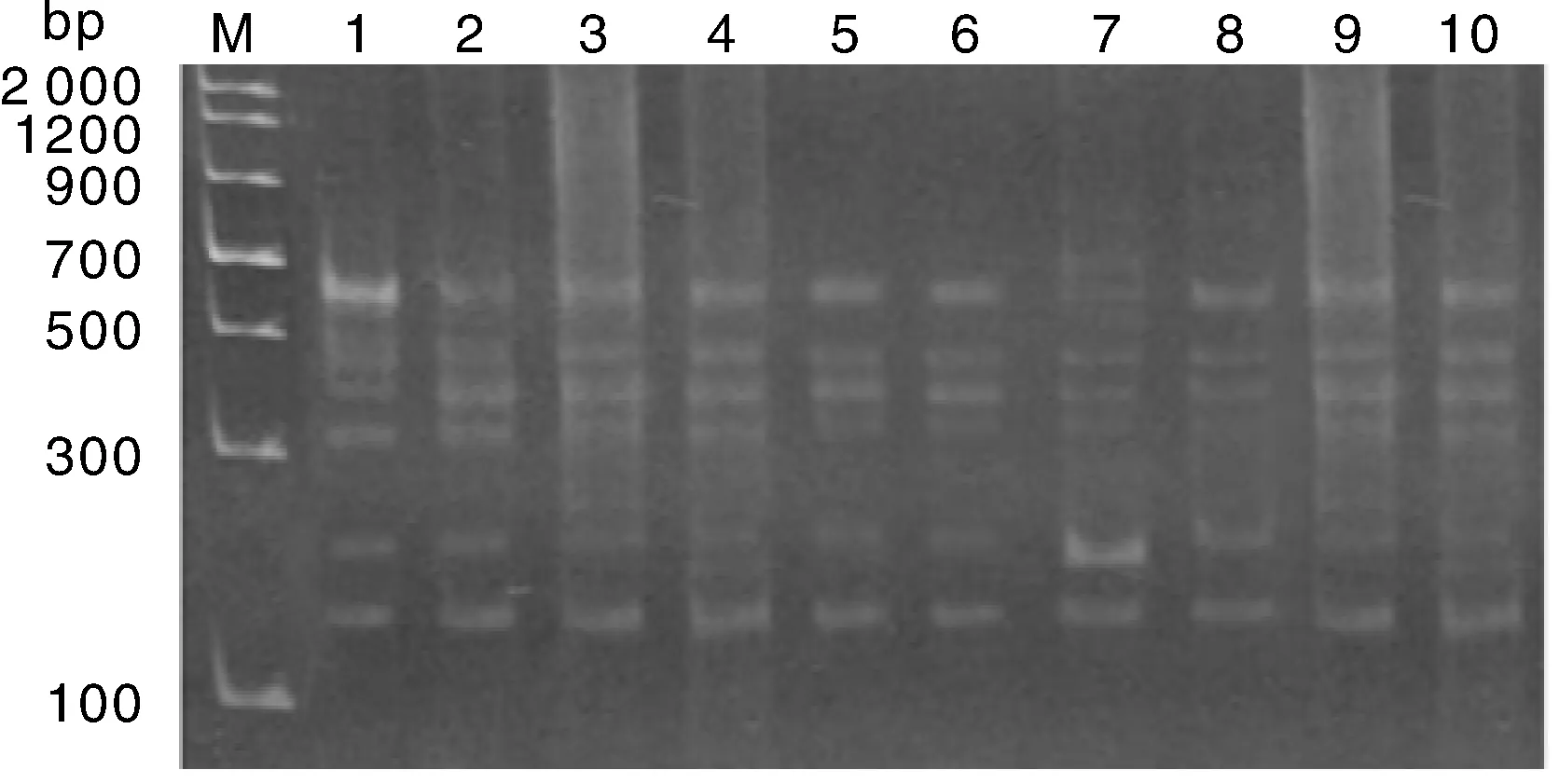
1~2, 常規(guī)繼代植株;3~10, 莖尖凍后再生植株圖7 廣豐藥薯基因組的AFLP擴(kuò)增結(jié)果Fig.7 Profiles of AFLP of the Guangfeng medicinal yam
2.7 江西地方山藥基因型對(duì)包埋玻璃化法超低溫保存的影響
江西地方山藥不同基因型包埋玻璃化法超低溫保存的效果見圖8。從圖8可知,廣豐藥薯建立的莖尖包埋玻璃化法超低溫保存可以應(yīng)用到廣豐千金薯、瑞昌山藥、南城淮山和泰和竹篙薯的種質(zhì)資源保存中,成活率為46.8%~69.5%。其中,廣豐千金薯和泰和竹篙薯的成活率最高,南城淮山的成活率最低,與廣豐藥薯的成活率相比無(wú)顯著差異(P>0.05)。
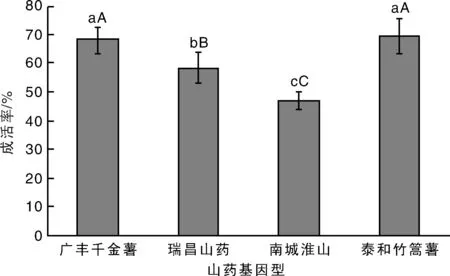
圖8 江西地方山藥不同基因型包埋玻璃化法超低溫保存效果比較Fig.8 Comparison of cryopreservation by encapsulation-vitrification effects of different genotypes of Jiangxi local yam
3 討論
包埋玻璃化法超低溫保存的第1步是對(duì)材料進(jìn)行預(yù)培養(yǎng)。研究表明,用較高濃度的蔗糖預(yù)培養(yǎng)數(shù)天,可以使材料預(yù)先脫除部分水分,提高莖尖胞質(zhì)濃度,從而增加抗凍力,促進(jìn)莖尖凍后的成活[22]。扁桃莖尖包埋珠在含有 0.3 mol·L-1蔗糖和50 g·L-1DMSO(二甲基亞砜)的MS培養(yǎng)基預(yù)培養(yǎng)1 d,成活率高達(dá)52%[23],但扁桃休眠莖尖包埋珠在含有0.5 mol·L-1蔗糖的MS培養(yǎng)基預(yù)培養(yǎng)3 d,成活率可達(dá)到45%,究其原因是休眠莖尖經(jīng)過(guò)冬天的低溫,已形成了保護(hù)性物質(zhì)(如可溶性糖和脯氨酸等),導(dǎo)致其耐凍能力增加,所以利用0.5 mol·L-1蔗糖進(jìn)行預(yù)培養(yǎng)可以代替0.3 mol·L-1蔗糖和50 g·L-1DMSO的預(yù)培養(yǎng)效果;而預(yù)培養(yǎng)時(shí)間從1 d增加到3 d,可能是增加預(yù)培養(yǎng)時(shí)間也可代替DMSO的預(yù)培養(yǎng)效果。本試驗(yàn)也表明,MS+2 mg·L-1KT+0.5 mg·L-1NAA培養(yǎng)基中添加0.75 mol·L-1蔗糖,預(yù)培養(yǎng)5 d有助于廣豐藥薯莖尖的凍后成活。
包埋玻璃化法超低溫保存的第2步是裝載液的裝載。裝載液一般為2 mol·L-1甘油和0.4 mol·L-1蔗糖[24]。前人的研究表明,使用甘油和蔗糖的裝載處理可以減輕后續(xù)步驟的玻璃化液即PVS2的毒害作用。本試驗(yàn)表明,用含有2 mol·L-1甘油和0.4 mol·L-1蔗糖的MS裝載液對(duì)廣豐藥薯莖尖裝載30 min,對(duì)其凍后成活有極顯著的促進(jìn)作用。這與扁桃[23]和李[25]莖尖包埋珠的裝載條件較為一致。用改良的裝載液對(duì)樹莓莖尖包埋珠進(jìn)行裝載,即用含2 mol·L-1甘油和0.9 mol·L-1蔗糖的裝載液處理90 min,成活率也可顯著提高,達(dá)85%[26]。因此,裝載液成分的濃度以及裝載液的裝載時(shí)間因植物種類而異[27]。
包埋玻璃化法超低溫保存的第3步是冰凍保護(hù)劑的脫水。冰凍保護(hù)劑一般用PVS2[18],由于PVS2的毒害效應(yīng),因此,要適度把握好PVS2的脫水時(shí)間和脫水溫度,PVS2脫水溫度一般設(shè)置為0 ℃或25 ℃[28]。研究表明,25 ℃時(shí),脫水時(shí)間可適量減少;0 ℃時(shí),時(shí)間可適當(dāng)延長(zhǎng)[29]。在0 ℃條件下,扁桃[23]、青島百合[30]、李[25]和樹莓[26]莖尖包埋珠PVS2的處理時(shí)間分別延長(zhǎng)到50,60,120和180 min,成活率均可顯著提高。本試驗(yàn)結(jié)果也表明,廣豐藥薯莖尖25 ℃處理40 min,成活率為37.2%,而0 ℃處理時(shí),脫水時(shí)間需要延長(zhǎng)到60 min,但成活率可顯著提高到43.9%。
包埋玻璃化法超低溫保存的最后一步是凍后材料的再生。研究表明,超低溫保存后適當(dāng)?shù)暮诎蹬囵B(yǎng)可以修復(fù)材料在超低溫保存期間造成的傷害。扁桃[23]和青島百合[30]莖尖超低溫保存后轉(zhuǎn)接至恢復(fù)培養(yǎng)基上暗培養(yǎng)一段時(shí)間,均可促進(jìn)成活率的增加。本試驗(yàn)結(jié)果也表明,凍后一定時(shí)間的黑暗培養(yǎng)可以顯著減輕廣豐藥薯莖尖的褐化,提高其凍后成活率。還有研究表明,超低溫保存后材料的成活率也會(huì)因山藥基因型而異[16]。本試驗(yàn)結(jié)果也支持了這種觀點(diǎn)。究其原因,這種差異可能與不同品種的遺傳差異有關(guān)。
植物材料DNA和染色體倍性的穩(wěn)定性是植物種質(zhì)資源保存的一個(gè)重要的問(wèn)題。因此,建立和優(yōu)化超低溫保存程序后,一般還需要對(duì)其再生植株的遺傳穩(wěn)定性進(jìn)行檢測(cè)和分析。目前,遺傳穩(wěn)定性的檢測(cè)和分析多采用分子標(biāo)記(如RAPD,AFLP,ISSR等)和細(xì)胞流式術(shù)(FCM)。朱文濤等[31]和陳冠群等[32]分別運(yùn)用AFLP技術(shù)分析了五葉草莓和百子蓮超低溫后的DNA帶型,均未發(fā)現(xiàn)明顯差異片段,表明超低溫保存前后的五葉草莓和百子蓮沒(méi)有發(fā)生遺傳變異。本研究中AFLP分析未檢測(cè)到位點(diǎn)變異,表明超低溫凍存的廣豐藥薯再生植株無(wú)DNA變異。Zarghami等[33]用AFLP檢測(cè)了3個(gè)馬鈴薯品種凍后再生植株的DNA變異,發(fā)現(xiàn)2個(gè)品種即Agria和Marphona未檢測(cè)到變異,而Agria的3% AFLP位點(diǎn)為多態(tài)性位點(diǎn),但作者也指出,這種變異可能并非來(lái)自冷凍傷害,也許是不同品種對(duì)離體培養(yǎng)的反應(yīng)不同造成的。本研究利用流式細(xì)胞儀對(duì)廣豐藥薯凍后再生植株的染色體倍性進(jìn)行鑒定,未見染色體倍性變異。這與Sch?fer-Menuhr等[34]和Galdiano等[35]在馬鈴薯和文心蘭上的染色體倍性鑒定結(jié)果是一致的。
[1] 蔡金輝, 嚴(yán)漸子, 黃曉輝,等. 山藥品種資源的分類研究[J]. 江西農(nóng)業(yè)大學(xué)學(xué)報(bào), 1999,21 (1): 53-57.
[2] 涂偉鳳, 湯潔, 涂玉琴, 等. 江西山藥生產(chǎn)現(xiàn)狀及發(fā)展優(yōu)勢(shì)[J]. 江西農(nóng)業(yè)學(xué)報(bào), 2012, 24(11): 21-24.
[3] 舒長(zhǎng)興, 彭琴, 付玉, 等. 瑞昌野生山藥種質(zhì)資源調(diào)查及組織培養(yǎng)試驗(yàn)[J]. 湖北農(nóng)業(yè)科學(xué), 2011, 50(10): 2060-2063.
[4] 熊國(guó)華, 嚴(yán)國(guó)勤, 熊兵才. 南城山藥本地種及其栽培技術(shù)[J]. 現(xiàn)代園藝, 2011 (2): 14.
[5] 方利平, 尤玉英, 邱翠金, 等. 廣豐千金薯高產(chǎn)栽培技術(shù)[J]. 安徽農(nóng)學(xué)通報(bào), 2009, 15(6): 147-148.
[6] 尹明華, 徐志堅(jiān), 章省琴, 等. 廣豐藥薯試管苗離體快繁技術(shù)的研究[J].中藥材, 2015, 38(11): 2245-2249.
[7] 王滿生. 泰和竹篙薯栽培技術(shù)[J]. 長(zhǎng)江蔬菜, 2000 (5): 14-15.
[8] 鐘齊剛, 陳章俊, 康來(lái)照, 等. 泰和竹篙薯優(yōu)質(zhì)高產(chǎn)栽培技術(shù)[J]. 安徽農(nóng)學(xué)通報(bào), 2012, 18(12): 240-241.
[9] 湯潔, 戴興臨, 涂玉琴, 等. 淮山藥新品種及栽培新技術(shù)研究[J]. 江西農(nóng)業(yè)學(xué)報(bào), 2011, 23(9): 57-59.
[10] 劉可越, 舒長(zhǎng)興, 石向群, 等. 不同產(chǎn)地山藥主要生化成分對(duì)比分析[J]. 湖北農(nóng)業(yè)科學(xué), 2011, 50(10): 2108-2110.
[11] 石印, 賈夢(mèng)雪, 狄瑋, 等. 芍藥花粉超低溫保存前后氧自由基變化研究[J]. 西北林學(xué)院學(xué)報(bào), 2015, 30(5): 86-90.
[12] 徐瑾, 石印, 劉芊, 等. 超低溫保存后玉蘭花粉的氧化應(yīng)激和細(xì)胞凋亡研究[J]. 植物生理學(xué)報(bào), 2015,51 (6): 916-920.
[13] 李明軍, 洪森榮, 徐鑫, 等. 懷山藥種質(zhì)資源的玻璃化法超低溫保存[J]. 作物學(xué)報(bào), 2006, 32(2): 288-292.
[14] 洪森榮, 李明軍. 玻璃化法超低溫保存懷山藥種質(zhì)的技術(shù)研究[J]. 中草藥, 2006, 37(11): 1715-1718.
[15] 李海兵, 周娜, 趙姣, 等. 懷山藥種質(zhì)資源的包埋玻璃化超低溫保存與植株再生[J]. 植物學(xué)報(bào), 2010, 45(3): 379-383.
[16] AGBIDINOUKOUN A, AHANHANZO C, ADOUKONOU-SAGBADJA H, et al. Impact of osmotic dehydration on the encapsulated apices survival of two yams(Dioscoreaspp.) genotypes from Benin [J].AmericanJournalofPathology, 2013, 65(2): 563-570.
[17] BARRACO G, SYLVESTRE I, COLLIN M, et al. Histocytological analysis of yam (Dioscoreaalata) shoot tips cryopreserved by encapsulation-dehydration.[J].Protoplasma, 2014, 251(1): 177-189.
[18] SAKAI A, KOBAYASHI S, OIYAMA I. Cryopreservation of nucellar cells of navel orange (CitrussinensisOsb. var.brasiliensisTanaka) by vitrification [J].PlantCellReports, 1990, 9(1): 30-33.
[19] 陳佰鴻, 李新生, 曹孜義, 等. 一種用透明膠帶粘取葉片表皮觀察氣孔的方法[J]. 植物生理學(xué)報(bào), 2004, 40(2): 215-218.
[20] 尹明華, 洪森榮. 黃獨(dú)微型塊莖胚性愈傷組織超低溫保存及遺傳穩(wěn)定性研究[J]. 中草藥, 2015, 46(17): 2623-2631.
[21] 郭文, 李婉琳, 肖繼坪, 等. 利用AFLP標(biāo)記構(gòu)建4種薯蕷屬植物的指紋圖譜[J]. 分子植物育種, 2015 ,13(3): 547-555.
[22] SAKAI A, ENGELMANN F. Vitrification, encapsulation-vitrification and droplet-vitrification: a review [J].CryoLetters, 2007, 28(3): 151-172.
[23] 陳加利, 姜喜, 肖巍, 等. 扁桃莖尖包埋玻璃化超低溫保存條件研究[J]. 塔里木大學(xué)學(xué)報(bào), 2014, 26(2): 110-114.
[24] HIRAI D, SAKAI A. Cryopreservation ofinvitro-grown axillary shoot-tip meristems of mint (MenthaspicataL.) by encapsulation vitrification [J].PlantCellReports, 1999, 19(2): 150-155.
[25] 陳紅, 韓曉瑩. 李莖尖包埋玻璃化法超低溫保存的條件探索[J]. 山地農(nóng)業(yè)生物學(xué)報(bào), 2013, 32(3): 276-279.
[26] 婁漢平, 郭修武, 代漢萍. 樹莓試管苗莖尖包埋玻璃化法超低溫保存[J]. 中國(guó)農(nóng)學(xué)通報(bào), 2010, 26(18): 59-62.
[27] SHARAF S A, SHIBLI R A, KASRAWI M A, et al. Cryopreservation of wild Shih (Artemisiaherba-albaAsso.) shoot-tips by encapsulation-dehydration and encapsulation-vitrification [J].PlantCell,TissueandOrganCulture, 2012, 108(3): 437-444.
[28] JEON S M, ARUN M, LEE S Y, et al. Application of encapsulation-vitrification in combination with air dehydration enhances cryotolerance ofChrysanthemummorifoliumshoots tips [J].ScientiaHorticulturae, 2015, 194: 91-99.
[29] HIRAI D, SHIRAI K, SHIRAI S, et al. Cryopreservation ofinvitro-grown meristems of strawberry (Fragaria×ananassaDuch.) by encapsulation-vitrification [J].Euphytica, 1998, 101(1): 109-115.
[30] 王金錄, 鈐泰琳, 趙美愛(ài). 青島百合包埋-玻璃化法超低溫保存技術(shù)研究[J]. 北方園藝, 2012 (18): 135-139.
[31] 朱文濤, 周厚成, 王子成. 五葉草莓超低溫保存及遺傳穩(wěn)定性分析[J]. 果樹學(xué)報(bào), 2013, 30(1): 55-61.
[32] 陳冠群, 李曉丹, 申曉輝. 百子蓮胚性愈傷組織玻璃化法超低溫保存體系建立及遺傳穩(wěn)定性分析[J]. 上海交通大學(xué)學(xué)報(bào)(農(nóng)業(yè)科學(xué)版), 2014, 32(5): 76-83.
[33] ZARGHAMI R, PIRSEYEDI M, HASRAK S, et al. Evaluation of genetic stability in cryopreservedSolanumtuberosum[J].AfricanJournalofBiotechnology, 2008, 7(16): 2798-2802.
[35] GALDIANO R F, LEMOS E G D M, VENDRAME W A. Cryopreservation, early seedling development, and genetic stability ofOncidiumflexuosumSims [J].PlantCell,TissueandOrganCulture, 2013, 114(1): 139-148.
(責(zé)任編輯 侯春曉)
Cryopreservation of Jiangxi yam shoot-tip by encapsulation-vitrification and its detection of genetic stability
YIN Ming-hua, XU Zhi-jian, ZHANG Sheng-qin, LYU Si-jie, ZENG Yan-hong, FU You-zhang, XIA Jin-hua, HONG Sen-rong*
(CollegeofLifeSciences,ShangraoNormalUniversity,Shangrao334001,China)
In this paper, cryopreservation procedure of Guangfeng medicinal yam shoot-tip by encapsulation-vitrification would be optimized by the method of plant tissue culture, and the genetic stability of regenerated plantlets after cryopreservation would be tested with AFLP molecular markers and flow cytometry methods. At the same time, this cryopreservation procedure by encapsulation-vitrification would be applied to other local varieties of Jiangxi yam, in order to lay the theoretical and technical foundation for long-term conservation of genusDioscoreagermplasm. The results showed that: The better pre-culture time of Guangfeng medicinal yam shoot-tips was 5 d, the better sucrose concentration in pre-culture medium was 0.75 mol·L-1; The better loading time was 40 min; The better dehydration temperature was 0 ℃, the better dehydration time was 60 min; 7-day dark culture could significantly improve the survival rate of shoot-tips after cryopreservation. At last, the genetic stability of regenerated plants was detected by AFLP molecular marker and flow cytometry (FCM), and no abnormal bands and chromosome ploidy changes were found. At the same time, no significant variation of stomatal parameters of leaves was observed. The cryopreservation technique was used in other genotypes of Jiangxi yam, the survival rate was about 40%-85%. Therefore, cryopreservation procedure by encapsulation-vitrification could guarantee the genetic stability of Jiangxi yam germplasm resources. The results of this study provided a certain technical support for the establishment of cryopreservation bank of Jiangxi yam germplasm resources.
Jiangxi yam; shoot-tip; encapsulation-vitrification; cryopreservation; genetic stability
10.3969/j.issn.1004-1524.2016.06.13
2015-11-22
國(guó)家自然科學(xué)基金(31360072);江西省教育廳2014年度科學(xué)技術(shù)研究一般項(xiàng)目(GJJ14712);國(guó)家級(jí)大學(xué)生創(chuàng)新創(chuàng)業(yè)訓(xùn)練計(jì)劃項(xiàng)目(201410416003)
尹明華(1973—),女,江西永新人,碩士,副教授,從事植物生物技術(shù)方面的研究。E-mail:yinminghua04@163.com
*通信作者,洪森榮,E-mail:hongsenrong@163.com
S632.1
A
1004-1524(2016)06-0984-08
尹明華, 徐志堅(jiān), 章省琴, 等. 江西山藥莖尖包埋玻璃化法超低溫保存及其遺傳穩(wěn)定性檢測(cè)[J]. 浙江農(nóng)業(yè)學(xué)報(bào), 2016, 28(6): 984-991.
