IL-11調控gp130/JAK2/STAT3信號通路促進Wistar大鼠雪旺細胞中PMP22的表達
2016-12-15 07:17:07郭巍李巖段會全孫超馮世慶徐云強
天津醫科大學學報 2016年6期
關鍵詞:信號
郭巍,李巖,段會全,孫超,馮世慶,徐云強
(天津醫科大學總醫院骨科,天津300052)
論著
IL-11調控gp130/JAK2/STAT3信號通路促進Wistar大鼠雪旺細胞中PMP22的表達
郭巍,李巖,段會全,孫超,馮世慶,徐云強
(天津醫科大學總醫院骨科,天津300052)
目的:探索白介素-11(IL-11)對大鼠雪旺細胞分泌髓鞘蛋白的調節作用及相關機制。方法:取成年Wistar大鼠坐骨神經,分離純化培養雪旺細胞。加入重組大鼠IL-11,分別采用Realtime RT-qPCR和Western boltting檢測雪旺細胞髓鞘蛋白的表達情況,進一步檢測IL-11調控髓鞘蛋白表達的相關通路的激活情況。結果:Realtime RT-qPCR和Western boltting結果均提示IL-11可以顯著提高雪旺細胞中周圍神經髓鞘蛋白22(PMP22)的表達水平,而對雪旺細胞中髓鞘堿性蛋白和髓鞘蛋白零的表達水平則沒無明顯影響,該生理作用可被JAK特異性抑制劑AG490所抑制。在經過IL-11處理的雪旺細胞中可以觀察到pJAK2(Y1007+Y1008)和pSTAT3(Y705)含量明顯高于對照組且伴隨著gp130表達的上調。結論:IL-11可通過調劑gp/30/ JAK2/STAT3信號通路以促進雪旺細胞中PMP22的表達。
白介素-11;雪旺細胞;周圍神經髓鞘蛋白22;JAK2/STAT3信號通路;大鼠
髓鞘為周圍神經系統中雪旺細胞(SCs)膜多次纏繞包裹軸突構成的鞘狀結構,在神經纖維的跳躍式傳導過程中提供絕緣保護作用及結構支持[1]。在脊椎動物中,髓鞘的厚度和所包裹的神經纖維是成比例的,當g比例(軸突直徑/有髓神經纖維直徑)為0.71時神經的傳導速率最高。周圍神經脫髓鞘病變或髓鞘形成不良會降低神經纖維的傳導速率,常常表現為肌肉無力和感覺缺失等典型臨床癥狀并最終導致進行性的軸突變性[2]。因此SCs形成髓鞘的能力對于周圍神經的發育和損傷后的再生過程至關重要[1,3]。SCs形成髓鞘的過程受到多種內源性因素的調節,如細胞外基質蛋白、神經營養因子、細胞因子等,其中以白介素-11(IL-11)和IL-6最為重要[4-5]。研究顯示IL-11在周圍神經元的發育、分化、再生及變性過程中發揮重要作用[6],并且在周圍神經病變患者體內可檢測到IL-11 mRNA表達水平的顯著提高[7]。大鼠坐骨神經損傷后可以觀察到其背跟神經節及損傷遠端華勒氏變性部位IL-11含量及其受體表達上調[8]。近期研究表明,IL-11可通過結合gp130參與周圍神經的再生過程,并且促進神經元中GAP-43的表達[6],但IL-11在髓鞘形成過程中的作用及相關機制仍有待進一步研究。相關研究證實SCs膜上存在IL-11受體gp130[9],敲除編碼白介素家族受體亞單位的gp130基因的小鼠表現出髓鞘退化的現象[10]。這表明IL-11可能通過gp130傳遞髓鞘形成的相關信號,但其下游效應分子仍有待進一步確認。為進一步闡明IL-11對外周神經髓鞘化的影響,本實驗在體外培養的SCs中加入IL-11,檢測其對SCs中周圍神經髓鞘蛋白22(PMP22)等表達的影響并進一步探索相關機制。
1 材料與方法
1.1 材料及試劑 雌性Wistar大鼠(150~200 g)、重組大鼠IL-11、胎牛血清、DMEM/F12細胞培養基、II型膠原酶、AG490、PD98059兔抗大鼠PMP22、兔抗大鼠MBP、兔抗大鼠MPZ、兔抗大鼠pJAK2、兔抗大鼠pSTAT3、兔抗大鼠GAPDH、羊抗兔HPR、羊抗兔 IgG(Dylight488)、GoScriptTM逆轉錄試劑盒(Promega)、GoTaq?qPCR試劑盒(Promega)、羅氏LightCycler 480熒光定量PCR儀等。
1.2 方法
1.2.1 SCs培養和純化 大鼠5只,頸椎脫臼處死后取雙側坐骨神經約1.5 cm,采用酶消化法培養SCs[11]。大致步驟為:解剖顯微鏡下仔細剝除神經外膜。將神經剪碎,體積約1 mm3。加入II型膠原酶,濃度0.1%,體積約為神經組織體積20倍。在二氧化碳培養箱中充分混合消化35~45 min至無明顯組織塊。1 000 r/min離心5 min,棄上清。加入含10%胎牛血清的DMEM/F12培養基重懸細胞,輕輕吹打使細胞充分混勻,接種于層黏連蛋白包被的25 cm2細胞培養瓶中,密度為(3.1±0.17)×105個/mL。采用低濃度膠原酶差速消化法提純SCs[12]。大致步驟為:體外培養48 h后,移除未貼壁的細胞及組織碎片,加入0.05%Ⅱ型膠原酶(0.1 mL/cm2)37℃消化35~40 min后水平輕輕晃動培養瓶使SCs與瓶底分離,收集細胞懸液于15 mL離心管中,1 000 r/min離心5 min,棄上清。加入含有10%胎牛血清的DMEM/F12培養基重懸細胞,接種于培養瓶中,密度為(1.8±0.12)×105個/ mL。48h后,重復上述過程,進行SCs第二次純化。
1.2.2 S100β染色鑒定SCs 4%多聚甲醛固定細胞30 min,磷酸鹽緩沖液(pH=7.4)漂洗3次,每次5 min。加入0.3%Triton X-100透膜。加入兔抗大鼠S100β(1∶200),4℃過夜。磷酸鹽緩沖液漂洗3次,每次5 min;加入熒光二抗(羊抗兔IgG,1∶300,Dylight488),室溫下避光孵育2 h,磷酸鹽緩沖液漂洗3次,每次5 min。DAPI甘油封片。共聚焦顯微鏡觀察及拍照。
1.2.3 Real time RT-qPCR 取純化后的SCs,將其培養基換為含0.1%FBS的DMEM/F12培養12 h。隨后加入相應濃度的IL-11(0、0.5、1.0、1.5 μg/mL)繼續培養12 h。Trizol法提取細胞總mRNA,利用GoScriptTM逆轉錄試劑盒得到cDNA。采用GoTaq? qPCR Master Mix試劑盒和羅氏LightCycler 480熒光定量PCR儀定量分析目的基因的表達量。相應引物序列為:MBP,F:5′-AGAGTCCGACGAGCTTCAG A-3′,R:5′-CAGGTACTTGGATCGCTGTG-3′;MPZ, F:5′-TCTCAGGTCACGCTCTATGTC-3′,R:5′-GCCA GCAGTACCGAATCAG-3′;PMP22,F:5′-CCCAACT CCCAGCCACCA-TG-3′,R:5′-TCATTCGCGTTTCC GCAGGATC-3′;gp130 F:GCCCTTGGGAATGTCTCC TCAGAG R:TCTTCCATATG-AGCCGTGCAGACC GAPDH,F:5′-GTATGTCGTGGAGTCTACTGGCGT-3′, R:5′-TACTCCTTGGAGGCCATGTA-GGCC-3′。各目的基因的表達量由GAPDH基因表達量校準,采用2(-DeltaDeltaC(T))法計算各目的基因的相對含量[13]。
1.2.4 Western blotting 取純化后的SCs,將其培養基換為含0.1%FBS的DMEM/F12培養12 h進行饑餓處理。隨后加入相應濃度的IL-11于細胞培養基中培養12 h。在應用信號通路抑制劑的實驗中,分別在加入 IL-11之前 6 h和 2 h用 AG490和PD98059預處理SCs。用含有蛋白酶抑制劑和磷酸酶抑制劑的裂解液裂解細胞。4℃,12 000 r/min離心20 min后取上清液,與上樣緩沖液按5∶1混合后100℃變性10 min。10%SDS-PAGE凝膠電泳,每空上樣15 μg。電泳完成后,將凝膠上的蛋白電轉至PVDF膜上,加入5%的脫脂牛奶室溫封閉30 min。隨后加入相應的一抗(兔抗,工作濃度1∶3000,4℃過夜)及二抗(HRP標記的羊抗兔IgG,1∶6000,室溫1 h)。設GAPDH內參照,洗膜后用ECL化學發光法檢測結果,采用Image J凝膠分析軟件分析結果。
1.3 統計學處理 SPSS22.0統計軟件進行數據處理。Real time RT-qPCR結果和Western blotting灰度值均由GAPDH校正,分析結果均以平均值±標準誤表示。各組間數據采用獨立樣本t檢驗比較,差異有統計學意義為*P<0.05,**P<0.01。NS,無明顯差別。
2 結果
2.1 SCs的觀察和鑒定 體外培養的SCs包體呈梭形,向兩端發出雙極或三級突起。SCs特異性標記物S100β免疫熒光染色顯示,經過上述培養和純化過程,最終SCs的純度在95%以上。見圖1。
2.2 IL-11上調SCs中PMP22 mRNA含量并促進PMP22蛋白表達 在經純化后得到的SCs的培養基中加入不同濃度的IL-11(0、0.5、1.0、1.5μg/mL),12h后檢測SCs中髓鞘堿性蛋白(MBP)、髓鞘蛋白零(MPZ)、PMP22的mRNA和蛋白含量變化。Real time RT-qPCR結果顯示與對照組相比經IL-11處理的SCs中MBP和MPZ mRNA的含量并沒有明顯變化;但當IL-11濃度為1.0 μg/mL和1.5 μg/mL時,可以觀察到SCs中PMP22 mRNA含量明顯提高(n=3,P=0.002;n=3,P=0.011)(圖 2A)。Western blotting結果同樣顯示當IL-11濃度為1.0 μg/mL和1.5μg/mL時,SCs中PMP22蛋白含量顯著升高(n=3, P=0.001;n=3,P=0.001)(圖2B)。
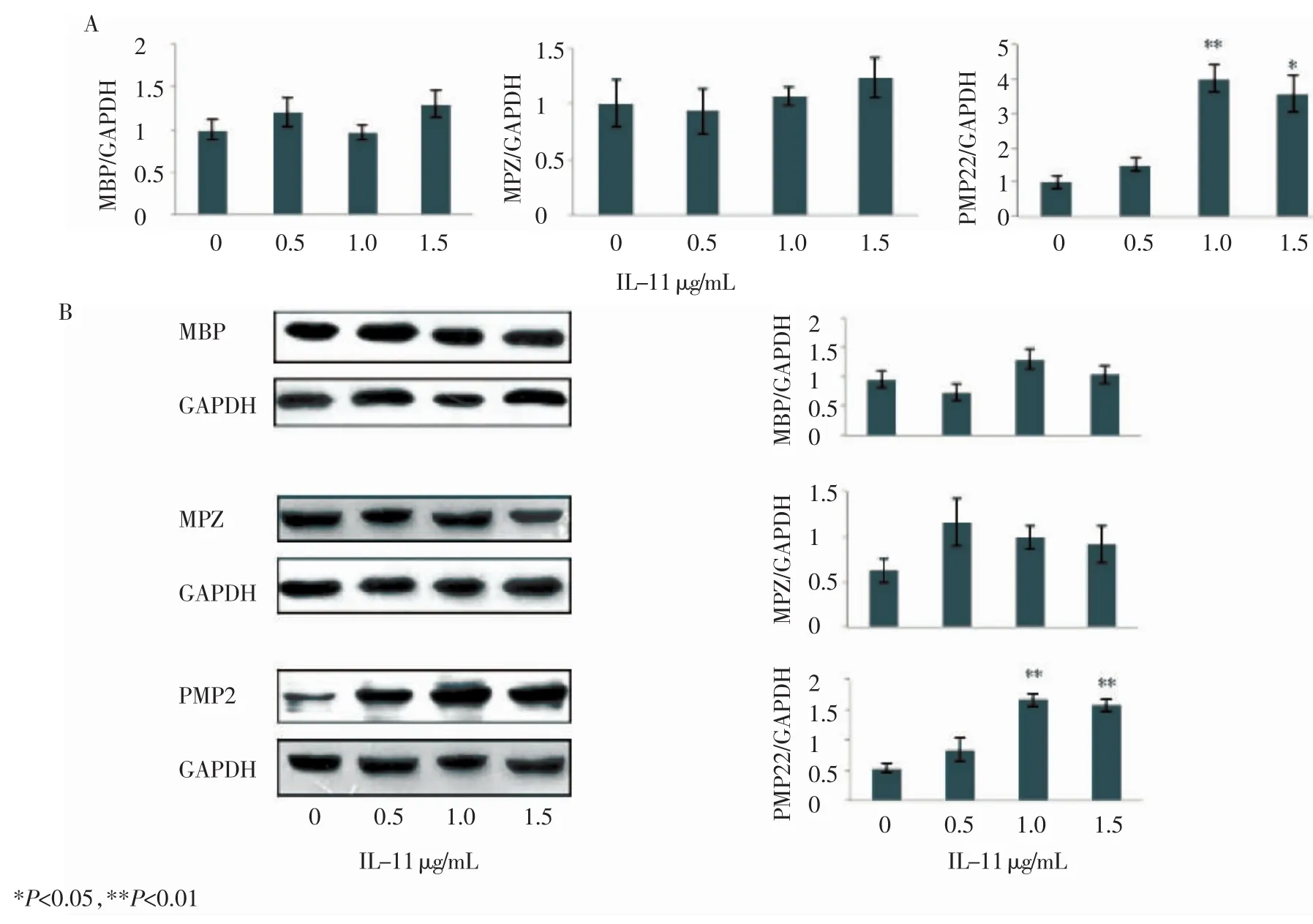
圖2 IL-11上調SCs中PMP22的表達水平Fig 2 IL-11 up-regulated the expression level of PMP22 in SCs
2.3 JAK2/STAT3通路在 IL-11促進 SCs合成PMP22過程中的重要作用 本研究中分別用JAK抑制劑AG490(50 μmol,6 h)和MAPKK抑制劑PD98059(25 μmol,2 h)預處理SCs,隨后再向培養基中加入IL-11(1.0 μg/mL,12 h)。觀察SCs中PMP22的表達情況。結果見圖3A,Real time RT-qPCR結果提示:PD98059并沒有抑制IL-11對SCs中PMP22表達的促進作用,而在經AG490預處理的SCs中,IL-11對PMP22表達的促進作用被顯著抑制(n=3,P=0.005);Western blotting(圖3B)結果進一步證實AG490可有效抑制IL-11對PMP22表達的促進作用(n=3,P=0.007)。
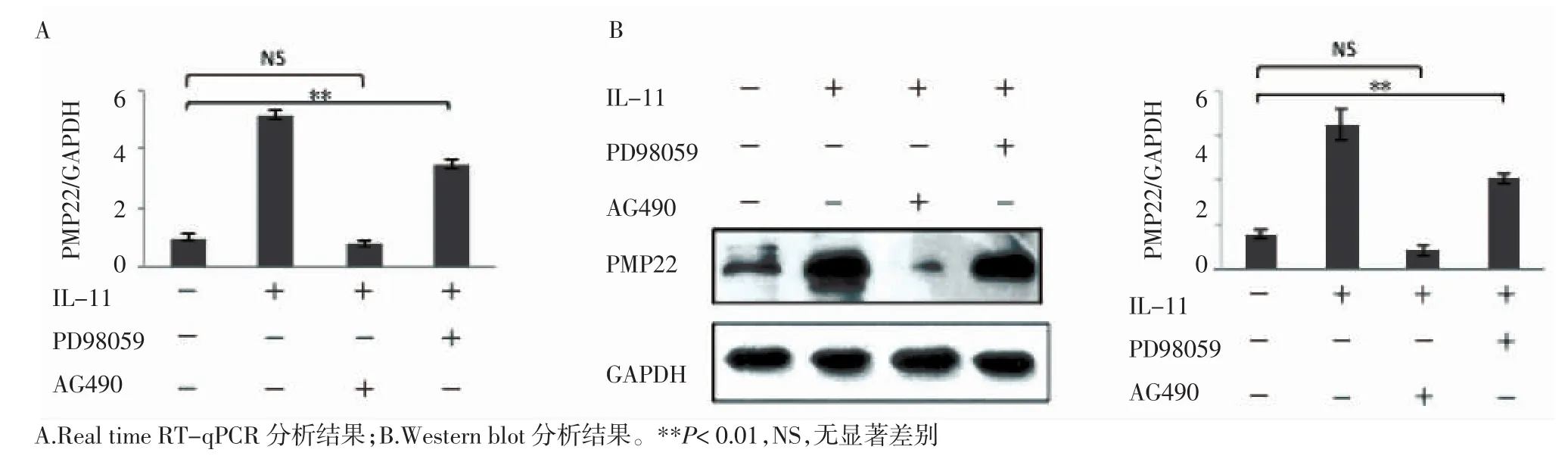
圖3 JAK2特異性抑制劑(AG490)和MAPKK特異性抑制劑(PD98059)對IL-11促進SCs中PMP22表達的影響Fig 3 Effects of JAK2 and MAPK inhibitors on IL-11 induced PMP22 expression
為進一步證實上述結果,本研究檢測了IL-11處理過的SCs中pJAK2(Y1007+Y1008)和pSTAT3(Y705)含量的變化。Western blotting結果分析顯示與對照組相比經過IL-11(1.0 μg/mL,12 h)處理的SCs中pJAK2和pSTAT3含量明顯提高(n=3,P=0.011;n=3,P=0.002),而總JAK2和STAT3含量則無明顯變化(圖4)。提示IL-11可以通過激活 SCs胞中 JAK2/STAT3通路促進PMP22的合成。
2.4 gp130在IL-11促進SCs中PMP22表達的作用 為了進一步驗證IL-11是否也是通過結合gp130來激活JAK2/STAT3信號通路以增強SCs中PMP22的合成,本研究通過Real time RT-qPCR和Western blotting的方法檢測經過IL-11處理的SCs中gp130的表達變化。Real time RT-qPCR結果(A)和Western blotting分析結果(B)均顯示在經過IL-11處理的SCs中gp130的表達量呈現明顯提高(n=3,P=0.001;n=3,P=0.020)(圖5)。結果提示,IL-11可通過結合gp130以激活JAK2/STAT3信號通路促進SCs中PMP22的表達。
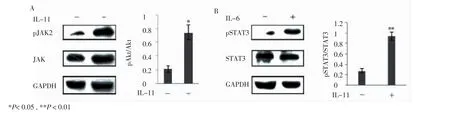
圖4 經IL-11處理后,SCs中JAK2和STAT3磷酸化情況Fig 4 Levels of JAK2 and STAT3 phosphorylation examined by Western blotting analysis
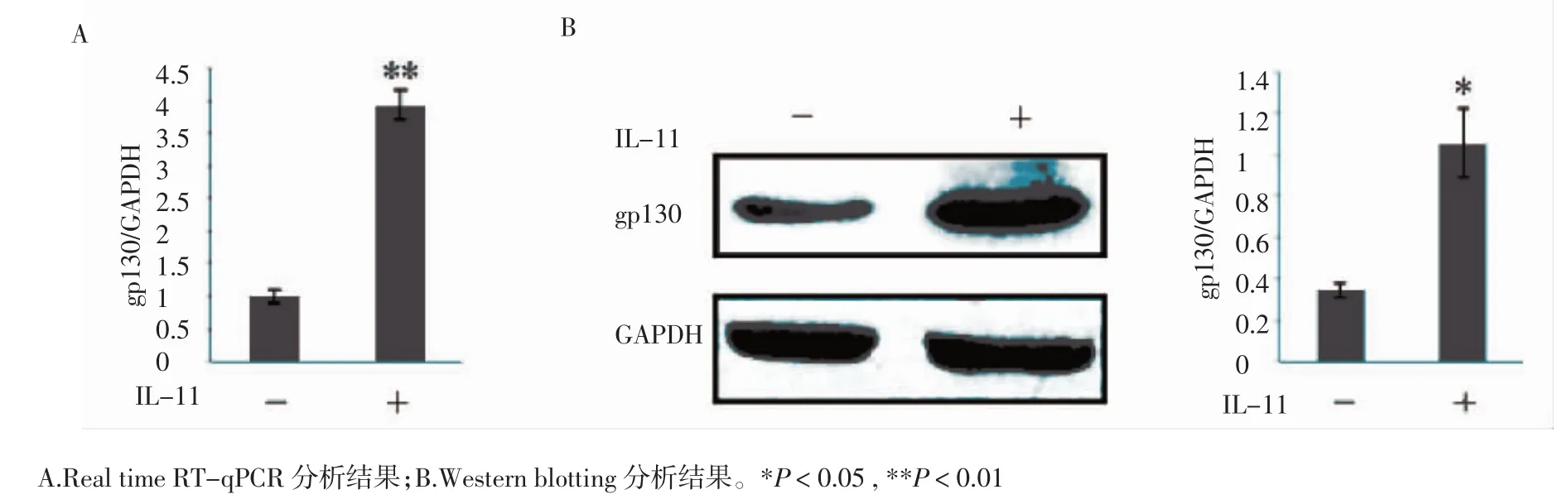
圖5 經IL-11處理后SCs中gp130的表達情況Fig 5 The expressional level of gp130 in SCs treated with IL-11
3 討論
本研究證實,IL-11可以顯著提高 SCs中PMP22的表達而對MBP和MPZ的表達無明顯作用。在非神經組織中,IL-11主要通過JAK2/STAT3通路和RAS/MAPK通路發揮作用[14-15]。本研究發現IL-11對SCs中PMP22表達的促進作用可被JAK特異性抑制劑AG490所抑制,并且在經過IL-11處理的SCs細胞中pJAK2(Y1007+Y1008)和pSTAT3(Y705)含量明顯高于對照組。上述實驗結果表明IL-11可通過激活JAK2/STAT3通路上調SCs中PMP22的表達,但IL-11是如何激活JAK2/STAT3信號的具體機制仍不明確。近來有研究表明IL-6家族(包括白血病抑制因子、IL-6、IL-11、抑瘤素M、睫狀神經生長因子等)可通過結合其共同受體gp130激活JAK/STAT通路以促進視網膜神經節細胞軸突生長[16]。本實驗結果顯示經過IL-11處理的SCs中gp130表達明顯上調。因此我們認為IL-11可通過結合gp130激活JAK2/STAT3信號通路以促進SCs中PMP22的表達。
PMP22是一種由SCs分泌的糖蛋白四聚體,是構成外周神經髓鞘的重要組成部分。盡管PMP22在髓鞘中含量豐富,但其生理作用尚未完全闡明。作為髓鞘形成過程中的重要部分,小鼠出生后和神經再生的過程中PMP22含量會明顯上調[17]。研究顯示PMP22基因缺陷與許多遺傳性神經系統病變有關,如腓骨肌萎縮癥1A型、遺傳性壓力敏感性周圍神經病變等[18]。Amici等[19]研究發現PMP22可與整合α6β4、層粘連蛋白形成復合體參與SCs與細胞外基質的相互作用,敲除IL-11受體gp130基因也將導致小鼠外周神經髓鞘退化[10]。結合以上實驗結果可得出IL-11很有可能通過gp130/JAK2/STAT3信號通路調節SCs中PMP22的表達進而形成和穩定外周神經髓鞘。
早期研究證實cAMP和microRNA-29a分別可以在轉錄層面和轉錄后層面調節PMP22的表達[20]。但這兩種調節過程與本實驗結果之間的關系仍有待進一步研究。坐骨神經橫斷后,背跟神經節中IL-11 mRNA和IL-1的含量在1 d之內快速升高,在隨后的1~2周內逐漸降低[8,21];Sheu等[22]發現在神經斷端的近端和遠端STAT3的磷酸化作用也在1 d之內達到峰值,并且持續時間超過2周;而PMP22 mRNA含量在神經損傷后則迅速下降,1周后逐漸上升[23]。因此IL-11可能是通過gp130/JAK2/STAT3信號通路參與PMP22在外周神經損傷后期表達的上調,但關于外周神經損傷后PMP22表達迅速下降的機制尚不明確。關于IL-11和gp130/JAK2/STAT3信號通路在外周神經再生中的具體作用仍有待進一步研究。
綜上所述,本研究證實IL-11可通過gp130/ JAK2/STAT3信號通路調節SCs中PMP22的表達。如果該通路在生物體內的髓鞘形成過程中也具有重要作用,則可將gp130作為藥物靶點[24],研制新型藥物調控gp130的信號以提高治療外周神經髓鞘病變的效果。同樣,在SCs移植修復神經損傷的治療中,也可將gp130/JAK2/STAT3作為關鍵通路,以調控髓鞘的形成。
[1] Nave K A.Myelination and support of axonal integrity by glia[J]. Nature,2010,468(7321):244
[2] Nave K A,Sereda M W,Ehrenreich H.Mechanisms of disease: inherited demyelinating neuropathies-from basic to clinical research[J].Nat ClinPract Neurol,2007,3(8):453
[3] Stassart R M,Fledrich R,Velanac V,et al.A role for Schwann cellderived neuregulin-1 in remyelination[J].Nat Neurosci,2013,16 (1):48
[4] Chen Z L,Yu W M,Strickland S.Peripheral regeneration[J].Annu Rev Neurosci,2007,30:209
[5] Richner M,Ulrichsen M,Elmegaard S L,et al.Peripheral nerve injury modulates neurotrophin signaling in the peripheral and central nervous system[J].Mol Neurobiol,2014,50(3):945
[6] Elsaeidi F,Bemben M A,Zhao X F,et al.Jak/Stat signaling stimulates zebrafish optic nerve regeneration and overcomes the inhibitory actions of Socs3 and Sfpq[J].J Neurosci,2014,34(7):2632
[7] Uceyler N,Riediger N,Kafke W,et al.Differential gene expression of cytokines and neurotrophic factors in nerve and skin of patients with peripheral neuropathies[J].J Neurol,2015,262(1):203
[8] Ito Y,Yamamoto M,Li M,et al.Temporal expression of mRNAs for neuropoietic cytokines,interleukin-11(IL-11),oncostatin M(OSM), cardiotrophin-1(CT-1)and their receptors(IL-11Ralpha and OSMRbeta)in peripheral nerve injury[J].Neurochem Res,2000,25 (8):1113
[9] Ozaki A,Nagai A,Lee Y B,et al.Expression of cytokines and cytokine receptors in human Schwann cells[J].Neuroreport,2008,19 (1):31
[10]Steelman A J,Zhou Y,Koito H,et al.Activation of oligodendroglial Stat3 is required for efficient remyelination[J].Neurobiol Dis, 2016,91:336
[11]Tao Y.Isolation and culture of Schwann cells[J].Methods Mol Biol, 2013,1018:93
[12]Jin Y Q,Liu W,Hong T H,et al.Efficient schwann cell purification by differential cell detachment using multiplex collagenase treatment[J].J Neurosci Methods,2008,170(1):140
[13]Livak K J,Schmittgen T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2(-DeltaDeltaC(T))Method[J]. Methods,2001,25(4):402
[14]Ernst M,Thiem S,Nguyen P M,et al.Epithelial gp130/Stat3 functions:an intestinal signaling node in health and disease[J]. SeminImmunol,2014,26(1):29
[15]Winship A L,Van Sinderen M,Donoghue J,et al.Targeting interleukin-11 receptor-α impairs human endometrial Cancer cell proliferation and invasion in vitro and reduces tumor growth and metastasis in vivo[J].Mol Cancer Ther,2016,15(4):720
[16]Pernet V,Joly S,Dalkara D,et al.Long-distance axonal regeneration induced by CNTF gene transfer is impaired by axonal misguidance in the injured adult optic nerve[J].NeurobiolDis,2013,51:202
[17]Notterpek L,Snipes G J,Shooter E M.Temporal expression pattern of peripheral myelin protein 22 during in vivo and in vitro myelination[J].Glia,1999,25(4):358
[18]vanPaassen B W,van der Kooi A J,van Spaendonck-Zwarts K Y,et al.PMP22 related neuropathies:Charcot-Marie-Tooth disease type 1A and Hereditary Neuropathy with liability to Pressure Palsies[J]. Orphanet J Rare Dis,2014,9:38
[19]Amici S A,Dunn W A,Murphy A J,et al.Peripheral myelin protein 22 is in complex with alpha6beta4 integrin,and its absence alters the Schwann cell basal lamina[J].J Neurosci,2006,26(4):1179
[20]Verrier J D,Lau P,Hudson L,et al.Peripheral myelin protein 22 is regulated post-transcriptionally by miRNA-29a[J].Glia,2009,57 (12):1265
[21]Saab C Y,Shamaa F,El Sabban M E,et al.Transient increase in cytokines and nerve growth factor in the rat dorsal root ganglia after nerve lesion and peripheral inflammation[J].J Neuroimmunol,2009, 208(1/2):94
[22]Sheu J Y,Kulhanek D J,Eckenstein F P.Differential patterns of ERK and STAT3 phosphorylation after sciatic nerve transection in the rat[J].Exp Neurol,2000,166(2):392
[23]Jung J,Coe H,Michalak M.Specialization of endoplasmic reticulum chaperones for the folding and function of myelin glycoproteins P0 and PMP22[J].FASEB J,2011,25(11):3929
[24]Fischer P,Hilfiker-Kleiner D.Role of gp130-mediated signalling pathways in the heart and its impact on potential therapeutic aspects [J].Br J Pharmacol,2008,153(Suppl 1):S414
(2016-05-11收稿)
IL-11 enhances expression of PMP22 in cultured Schwann cells via gp130/JAK2/STAT3 signaling pathway
GUO Wei,LI Yan,DUAN Hui-quan,SUN Chao,FENG Shi-qing,XU Yun-qiang
(Department of Orthopaedics,General Hospital,Tianjin Medical University,Tianjin 300052,China)
Objective:To examine the effect of IL-11 on the expression of myelin proteins in cultured Schwann cells.Methods:Schwann cells were cultured and purified from sciatic nerves of adult Wistarrats.IL-11 was used to treat purified Schwann cells.The expression level of myelin proteins was examined with Real time RT-qPCR and Western boltting.Results:IL-11 significantly increased the expression level of PMP22 in Schwann cells,but not those of MPZ and MBP.And this effect was significantly inhibited by AG490,anspecificinhibitor of JAK.And the levels of JAK2 and STAT3 phosphorylation were significantly increased in the presence of IL-11 stimulated Schwann cells compared with the absence of IL-11,which was accompanied with the up-regulation of gp130.Conclusion:IL-11 can enhance the production of PMP22 in Schwann cells via gp130/JAK2/STAT3 signaling pathway and thus may contribute to myelination.
interleukin-11;Schwann cells;PMP22;JAK2/STAT3 signaling pathway;rat
R68
A
1006-8147(2016)06-0478-05
郭巍(1989-),男,碩士在讀,研究方向:髓鞘再生;通信作者:徐云強,E-mail:docxu@sina.com。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
