雜交石竹授粉受精及其胚胎發育過程的觀察
2016-12-17 02:11:56張賽陽郝鵬博楊秋生
西北植物學報 2016年10期
關鍵詞:生長
張賽陽,栗 燕,郝鵬博,楊秋生
(河南農業大學 林學院,鄭州 450002)
?
雜交石竹授粉受精及其胚胎發育過程的觀察
張賽陽,栗 燕,郝鵬博,楊秋生*
(河南農業大學 林學院,鄭州 450002)
以‘雜交石竹’為試驗材料,利用熒光顯微鏡觀察其授粉后花粉萌發、花粉管生長情況,采用石蠟切片法對其受精及胚胎發育過程進行觀察研究。結果表明:(1)授粉后1 h母本柱頭上少量花粉開始萌發;授粉后4 h大量花粉萌發,花粉管生長至柱頭中部有胼胝質出現;授粉后6 h花粉管生長至子房組織并有少量與胚珠結合;授粉后15 h柱頭中出現大量胼胝質,花粉管與胚珠結合數增多;授粉后24 h胚珠周圍出現多條花粉管,其中1條花粉管進入胚珠,部分進入胚囊的花粉管卷曲盤繞生長并產生胼胝質;精細胞與極核的融合主要發生在授粉后17~48 h,與卵細胞融合主要于授粉后1~3 d。(2)雜交石竹胚發育經過原胚、球形胚、棒狀形胚、心形胚、魚雷形胚和子葉形胚階段。(3)雜交障礙表現為:只有游離的胚乳核而無胚發育的胚囊、合子未分裂、兩極核未融合、球形胚敗育。研究表明,雜交石竹存在受精前和受精后障礙,這是導致其結實率低的主要原因。
雜交石竹;花粉萌發;花粉管生長;受精;胚胎發育;
石竹(Dianthuschinensis)原產中國,是優良的庭院花卉,可用于花壇、花境或盆栽觀賞,也是制作花籃花束的切花材料。雜交石竹(Dianthushybridus)為石竹科石竹屬多年生草本植物,由中國石竹(Dianthuschinensis)與美國石竹(Dianthusbarbatus)雜交而成,以種子繁殖為主,生長勢強、一致性好、花色豐富、開花量大,在園林上應用廣泛[1]。目前,國內外對香石竹雜交育種方面研究較多,而對雜交石竹少見報道,中國在雜交石竹品種選育方面研究相對落后,園林綠化應用的雜交石竹品種多數為國外育種公司培育。在雜交石竹制種過程中,經常遇到母本結實率低,多數形成空種莢或種莢中種子較少的問題。因此,探究雜交石竹授粉受精特性及胚胎發育對提高種子產量有重要意義。
本試驗利用熒光顯微鏡對雜交石竹花粉萌發、花粉管生長過程進行觀察,采用石蠟切片法對雜交受精及胚胎發育進程進行觀察研究,試圖探索其授粉受精特性及雜交不親和原因,提高雜交制種產量,為雜交石竹育種及相關研究提供理論和技術依據。
1 材料和方法
1.1 材料
供試材料為雜交石竹(Dianthushybridus),田間試驗于2015年4~6月在河南省濟源市雜交石竹制種基地進行,室內試驗在河南農業大學觀賞植物實驗室進行。
1.2 方法
1.2.1 花粉萌發及花粉管生長過程觀察 于2015年5月盛花期,上午10:00采集當天散粉的父本花粉,授在母本柱頭上,于授粉后1、4、6、15和24 h分別采集4~6個雌蕊,用FAA固定液(38%甲醛5 mL∶冰醋酸5 mL∶70%酒精90 mL)固定。取固定好的待觀察材料,首先依次用70%酒精、50%酒精、30%酒精、蒸餾水分別浸泡2 h,然后用8 mol/L NaOH水浴(56 ℃)3~12 h,蒸餾水沖洗2~3次,每次10~20 min,之后用0.1%苯胺藍溶液染色。Nikon熒光顯微鏡觀察花粉萌發、花粉管生長情況。
1.2.2 受精過程及胚胎發育觀察 于2015年5月盛花期,上午10:00采集當天散粉的父本花粉,授在母本柱頭上,于授粉后17 h、1 d、2 d、3 d、5 d、12 d、15 d分別采集4~6個雌蕊,用FAA固定液固定。采用常規石蠟切片法,切片厚度8~10 μm,番紅固綠染色,中性樹膠封片,在Motic數碼顯微鏡下觀察受精和胚胎發育情況。
2 結果與分析
2.1 花粉萌發及花粉管生長過程觀察
利用熒光顯微鏡對雜交石竹花粉萌發及花粉管生長過程進行觀察研究,結果顯示:授粉后1 h柱頭附著大量花粉并有少量花粉開始萌發(圖版Ⅰ,1);授粉后4 h大量花粉開始萌發,花粉管穿過柱頭乳突細胞進入柱頭(圖版Ⅰ,2),授粉后4 h花粉管生長至柱頭1/2處并出現胼胝質塞,同時觀察到異常花粉粒出現較大胼胝質塊,花粉管停止生長(圖版Ⅰ,3);授粉后6 h花粉管已生長至子房組織并有少量與胚珠結合(圖版Ⅰ,4);授粉后15 h柱頭中出現大量胼胝質(圖版Ⅰ,5),授粉后15 h花粉管與胚珠結合數增多(圖版Ⅰ,6);授粉后24 h胚珠周圍出現多條花粉管,其中一條花粉管進入胚珠(圖版Ⅰ,7),授粉后24 h部分進入胚珠的花粉管卷曲盤繞生長并產生胼胝質反應(圖版Ⅰ,8、9)。觀察結果表明雜交石竹花粉萌發和花粉管生長均無明顯阻礙,受精前障礙主要在于進入胚囊內的花粉管卷曲盤繞生長并產生胼胝質而無法完成受精。
2.2 受精過程觀察
利用石蠟切片對雜交石竹受精過程進行觀察,結果顯示:授粉后17 h花粉管穿過珠心組織,經珠孔進入胚囊并與助細胞結合釋放內容物,助細胞中內含物較多,染色呈現較深的紅色(圖版Ⅰ,10);授粉后1 d其中1個精細胞貼附在1個極核核膜上(圖版Ⅰ,11),并與二極核細胞融合形成3核的初生胚乳核(圖版Ⅰ,12);授粉后1 d在珠孔端觀察到1個精細胞向卵核移動,進入卵細胞的細胞質(圖版Ⅰ,13);精核貼附在卵核核膜上,進行核的融合(圖版Ⅰ,14);授粉后2 d在胚囊中觀察到合子和初生胚乳細胞,合子靠近珠孔端,初生胚乳核在胚囊的中心位置,受精卵的核比初生胚乳核小(圖版Ⅰ,15)。通過大量石蠟切片觀察發現,雜交石竹配子融合速度快,融合時間很短,但配子融合時間段相對較長,雜交石竹精細胞與極核的融合主要發生在授粉后17~48 h,與卵細胞融合過程主要于授粉后1~3 d。
2.3 胚胎發育觀察
利用石蠟切片對雜交石竹胚胎發育進行觀察,結果顯示:授粉后2 d卵核受精形成合子,初生胚乳核開始分裂形成多個游離胚乳自由核(圖版Ⅱ,1);授粉后3 d可以觀察到合子分裂形成原胚,以及正在分裂的原胚和大量的游離胚乳核(圖版Ⅱ,2、3);授粉后5 d原胚細胞繼續分裂形成球形胚,球形胚周圍存在游離胚乳自由核(圖版Ⅱ,4);授粉后12 d,球形胚體積增大,細胞緊密排列,周圍存在大量胚乳自由核(圖版Ⅱ,5);授粉后12 d,在部分胚囊中觀察到棒狀形胚和心形胚(圖版Ⅱ,6、7);授粉后15 d,觀察到大量魚雷形胚和子葉形胚(圖版Ⅱ,8、9)。觀察結果表明雜交石竹胚胎發育經歷原胚、球形胚、棒狀形胚、心形胚、魚雷形胚和子葉形胚階段。
同時對異常胚胎發育進行觀察,結果顯示:授粉后2 d觀察到只有游離的胚乳核而無胚發育的胚囊(圖版Ⅱ,10);授粉后3 d,觀察到部分胚囊中有合子未分裂的現象,而在這個時期正常的胚胎已發育到原胚階段(圖版Ⅱ,11),同時觀察到有兩極核未融合的現象(圖版Ⅱ,12),和只有游離的胚乳發育而無胚發育的胚囊(圖版Ⅱ,13);授粉后5 d,敗育的球形胚(圖版Ⅱ,14);授粉后12 d,開始解體的球形胚,核膜界限變模糊(圖版Ⅱ,15)。觀察結果表明雜交石竹受精后障礙發生在受精過程和胚胎發育時期,表現為:合子未分裂、兩極核未融合、只有游離的胚乳發育而無胚發育的胚囊、球形胚敗育。
3 討 論
雜交親和性的障礙發生在生殖過程的不同階段,主要包括受精前障礙和受精后障礙。被子植物中,授粉后花粉在柱頭上的附著量、萌發及花粉管生長情況和受精后胚胎發育情況對雜交結實率影響很大,其中任何一個環節出現異常都有可能導致較低的結實率[2]。
王沖等[3]通過對君子蘭種間雜交及自交親和性研究,熒光顯微觀察發現在君子蘭種間雜交花粉萌發與花粉管伸長均存在胼胝質現象,母本柱頭表面出現大量胼胝質,而這些胼胝質阻礙花粉萌發和花粉管向下伸長。周旭紅等[4]通過對不同倍性香石竹雜交花粉管生長熒光顯微觀察,發現花粉管先端沉積胼胝質而停止生長。櫟屬的2個種進行雜交時,柱頭中出現的大量胼胝質使花粉管不能到達胚囊,導致雜交失敗[5]。本試驗觀察到花粉能夠在柱頭上大量萌發,少量花粉粒中出現較大胼胝質塊花粉管停止生長,小部分花粉管頂端胼胝質異常積累、膨大,母本柱頭中觀察到花粉管壁內出現大量胼胝質塞,花粉管能夠順利生長至柱頭基部,并未觀察到胼胝質阻礙花粉管生長的現象。王文鵬等[6]在夏蠟梅與光葉臘梅屬間雜交未觀察到胼胝質阻礙花粉管生長的現象,花粉萌發和花粉管生長階段均無明顯阻礙。花粉管中胼胝質栓塞的存在將花粉管后部內腔堵死,使內腔成為不連續系統,同時防止管內原生質倒流,從而保證花粉管的頂端生長[7]。當花粉管生長到達一定長度后,在后端形成胼胝質,當花粉管延長時,在一定的間隔位置多次形成胼胝質塞,因此胼胝質塞區隨著花粉管生長而延長[8]。
胚珠周圍出現多條花粉管,其中1條與胚珠結合,在雙受精作用中,通常有阻止多條花粉管進入胚囊的機理,因此僅有1條花粉管攜帶的1對精細胞到達胚囊[8]。花粉管在它進入的助細胞中停止生長和釋放精子,是由于當花粉管進入助細胞后,它周圍的環境發生變化而觸發的[8]。周旭紅等[9]在不同倍性香石竹雜交受精過程及胚胎發育研究中,認為受精的子房頻率低,是不同倍性香石竹雜交結實率低的重要原因。楊鳳君等[10]在歐洲櫻桃與草原櫻桃雜交時,認為花粉管不能進入胚囊,雌雄配子不能相遇而導致不能結實。玉米等作物遠緣雜交中發現,花粉管雖然能夠進入胚囊,但并不一定能完成受精作用,出現胚囊不親和現象[11]。Williams等[12]在杜鵑花種間雜交研究中發現,花粉管經珠孔進入胚囊后,因卷曲盤繞等過度生長情況,而無法完成受精。本試驗觀察到經珠孔進入胚囊的一條花粉管在珠孔附近卷曲盤繞生長并出現胼胝質,這一現象表明,雖然花粉管能夠到達子房并進入胚囊,但不能完成雙受精,存在受精前障礙,這可能是導致雜交石竹結實率低的重要原因之一。
Daston等[13]在玄參科龍面花屬種間雜交時,觀察到父本花粉管進入子房和胚珠,但授粉后子房并沒有膨大,最終沒有得到雜交種子,存在受精后障礙,受精后障礙也是導致不同倍性香石竹雜交結實率低的原因之一[9]。胚胎發育不良是引起種子敗育的主要原因[14],較高比例的胚敗育是影響結實率的另一個重要因素[15],Deng等[16]在菊花和細裂亞菊的屬間雜交中發現較高比率的胚胎敗育是結實率低的主要原因,而花粉活力和花粉管萌發對結實率的影響不大。本試驗通過石蠟切片法對授粉后不同時間的雜交石竹子房進行觀察,發現雜交障礙發生在受精過程和胚胎發育時期,表現為:兩極核未融合、胚囊中合子未分裂、只有游離的胚乳核無胚發育的胚囊、球形胚敗育。因此,受精后障礙也是導致雜交石竹結實率低的重要原因之一。
通過對雜交石竹授粉后花粉管生長及胚胎發育情況進行觀察研究,發現雜交石竹存在受精前和受精后障礙,花粉管進入胚珠后卷曲盤繞生長并產生胼胝質、受精后合子未分裂、兩極核未融合、胚敗育,這幾個因素共同作用,是導致結實率低的主要原因,但引起雜交石竹雜交生殖障礙的機理還不清楚。今后在雜交石竹制種過程中,應選擇在花粉活力和柱頭可授性較強時期內進行授粉,增加受精胚珠的概率,以期獲得更多、更飽滿的雜交種子,提高雜交結實率和制種生產效率。
[1] 宋利娜,許 超,辛海波,等.雜交石竹雄性不育株離體快繁技術研究[J].北京農學院學報,2015,30(3):92-95.
SONG L N,XU C,XIN H B,etal. A study on in vitro rapid propagation technique for male sterile lines ofDianthushybridus[J].JournalofBeijingUniversioyofAgriculture,2015,30(3):92-95.
[2] 孫春青,陳發棣,房偉民,等.野菊與菊花雜交中花粉活力和柱頭可授性及胚胎發育研究[J].西北植物學報,2009, 29(7):1 335-1 341.
SUN C Q,CHEN F D,FANG W M,etal. Pollen viability, pistil receptivity and embryogenesis in the cross betweenDendranthemaindicumandD.grandiflorum[J].ActaBotanicaBoreali-OccidentaliaSinica,2009,29(7):1 335-1 341.
[3] 王 沖,雷家軍,姜 闖,等.君子蘭種間雜交及自交親和性[J]. 中國農業科學,2011,44(18):3 822-3 829.
WANG C, LEI J J,JIANG C,etal. Study on cross-compatibility of interspecific hybridization and selfing inCliviaLindl[J].ScientiaAgriculturaSinica,2011,44(18):3 822-3 829.
[4] 周旭紅,桂 敏,王繼華,等.不同倍性香石竹雜交花粉管生長熒光顯微觀察及結實研究[J].西北植物學報,2012,32(1):67-74.
ZHOU X H,GUI M,WANG J H,etal. Pollen Tube growth after crossing between different ploidsDianthuscaryophyllusby Fluorescence Microscopy and seed setting[J].ActaBotanicaBoreali-OccidentaliaSinica,2012,32(1):67-74.
[5] BOAVIDA L C,SILVA J P,FEIJO J A. Sexual reproduntion in the cork oak (QuercussuberL).II.Crossing intra-and interspecific barriers[J].SexualPlantReproduction,2001,14:143-152.
[6] 王文鵬,周莉花,劉華江,等. 夏蠟梅與美國蠟梅屬間雜交障礙的組織學機理[J].園藝學報,2013,40(10):1 943-1 950.
WANG W P ,ZHOU L H,LIU H J,etal. Histological reproductive barriers for intergeneric cross betweenSinocalycanthuschinensisandCalycanthusfloridusvar.oblongifolius[J].ActaHorticulturaeSinica,2013,40(10):1 943-1 950.
[7] ALEXANDER KRICHEVSKY, STANISLAV V. KOZLOVSKY, GUO W T,etal. How pollen tubes grow[J].DevelopmentBiology, 2007,(303):405-420.
[8] 胡適宜,朱 澂.,被子植物有性生殖圖譜[M].北京:科學出版社,2000.
[9] 周旭紅,桂 敏,陳 敏,等.不同倍性香石竹雜交受精過程及胚胎發育研究[J].西北植物學報, 2013, 33(1):1-6.
ZHOU X H,GUI M,CHEN M,etal. Fertilization and development of embryo in different interploidal crosses ofDianthuscaryophyllus[J].ActaBotanicaBoreali-OccidentaliaSinica,2013, 33(1):1-6.
[10] 楊鳳軍,臧忠婧,吳 瑕. 草原櫻桃種內和種間授粉親和性的熒光顯微觀察[J]. 果樹學報, 2015,(5):909-913.
YANG F J,ZANG Z J,WU X. Fluorescent microscope observation on pollination compatibility of Ground cherry[Cerasusfruticosa(Pall.)]crossing with different cherry cultivars[J].JournalofFruitScience,2015,(5):909-913.
[11] 段桃利.玉米與其近緣種屬雜交花粉管行為研究[D].四川雅安:四川農業大學,2008.
[12] WILLIAMS E G, KAUL V, ROUSE J L,etal. Overgrowth of pollen tubes in embryo sacs of Rhododendron following interspecific pollinations[J].AmerJBot,1986,(34):413-423.
[13] DASTON P M, MURRAY B G, HAMMETT K R W. Pollination systems hybridization barriers and meiotic chromosome behavior in Namesia hybrids[J].Euphytia,2006,151:173-185.
[14] 陳曉月,王金鑫,裴艷梅,等. 無核小棗與金絲小棗授粉受精過程的觀察[J]. 河北農業大學學報. 2014, 37(6):28-32.
CHEN X Y, WANG J X, PEI Y M,etal. Observation of pollination and fertilization ofZiziphusjujubaMill.‘Jinsixiaozao’ andZ.jujubaMill.‘Wuhexiaozao’[J].JournalofAgriculturalUniversityofHebei, 2014, 37(6):28-32.
[15] 胡適宜.被子植物生殖生物學[M].北京:高等教育出版社,2005.
[16] DENG Y M, TENG N J, CHEN S M,etal. Reproductive barriers in theintergeneric hybridization betweenChrysanthemumgrandiflorum(Ramat)Kitam.andAjaniaprzewalskiiPoljak.Euphytica,2010,174(1):41-50.
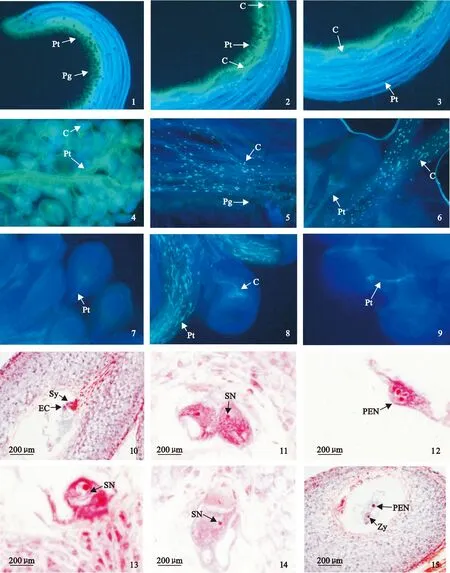
Pt.花粉管;Pg.花粉粒;C.胼胝質;Sy.助細胞;EC.卵細胞;SN.精核;PEN.初生胚乳核;Zy.合子1.柱頭附著大量花粉、授粉后1 h少量花粉開始萌發,×100;2. 授粉后4 h大量花粉開始萌發,少量花粉粒出現較大胼胝質,花粉管穿過柱頭乳突細胞進入柱頭,×100;3.授粉后4 h花粉管生長至柱頭1/2處并出現胼胝質塞,×100;4. 授粉后6 h花粉管已生長至子房組織并有少量與胚珠結合,×200; 5. 授粉后15 h柱頭中出現大量間斷胼胝質反應,×200;6. 授粉后15 h大量花粉管進入子房組織,花粉管與胚珠結合數增多,×200; 7. 授粉后24 h胚珠周圍出現多條花粉管,其中1條花粉管進入胚珠,×200;8、9. 授粉后24 h部分進入胚囊的花粉管卷曲盤繞生長并產生胼胝質,×400;10.授粉后17 h花粉管經珠孔進入胚囊并進入助細胞釋放內容物;11. 授粉后1 d1個精細胞貼附在1個極核核膜上;12. 授粉后1 d1個精細胞與二極核細胞融合形成3核的初生胚乳核;13. 授粉后1 d1個精細胞向卵核移動,進入卵細胞的細胞質;14. 授粉后1 d精核貼附在卵核核膜上;15. 授粉后2 d合子和初生胚乳細胞圖版Ⅰ ‘雜交石竹’花粉萌發、花粉管生長及受精過程Pt. pollen tube; Pg. Pollen grain; C. Callose response; Sy. Synergid; EC. Egg cell; SN. Sperm nuclei; PEN. Primary endosperm nucleus; Zy. ZygoteFig.1. Stigma attached to a large number of pollens a small amount of pollens began to germinationed in 1 h after pollination, ×100; Fig.2. Many of pollen grains on the stigma began to germinationed in 4 h after pollination, pollen grains appear larger callose, pollen tube through the stigma papilla cells into stigma, ×100; Fig.3. The pollen tubes grew to stigma 1/2 and callose plug in 4 h after pollination, ×100; Fig.4. Pollen tubes grew to the ovary tissue and a small amount of them entered the ovule in 6 h after pollination, ×200; Fig.5. Stigma callose reactions occurred in 15 h after pollination, ×200;Fig.6. the large number of pollen tubes into the ovary tissue, pollen tubes increased within ovule, in 15 h after pollination, ×200; Fig.7.Many of pollen tubes had arisen ovule around, one of the pollen tube entered the ovule, ×200; Fig.8,9. Some pollen tubes had formed coiled overgrowth and generated callose reaction in the embryo sac, ×400; Fig.10. The pollen tube entered into one synergid and released contents 17 h after pollination; Fig.11. Sperm nucleus adhered to the nuclear membrane of the polar nuclei 1 d after pollination; Fig.12. Sperm nucleus fused with two polar nuclei and the primary endosperm nucleus 1 d after pollination; Fig.13. Sperm nucleus moved towards egg nucleus and entered the cytoplasm of egg nucleus 1 d after pollination; Fig.14. Sperm nucleus adhered to the nuclear membrane of the egg cell 1 d after pollination; Fig.15. Zygote and primary endosperm nucleus 2 d after pollinationPlate Ⅰ ‘Dianthus hybridus’ pollen germination, pollen-tube growth and fertilization
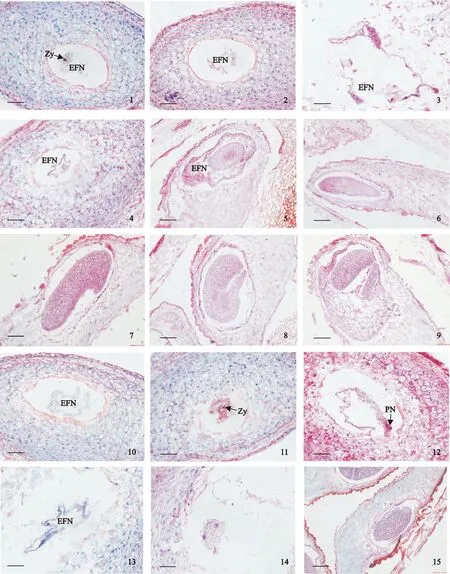
Zy.合子;EFN.胚乳自由核;PN.極核1.授粉后2 d,合子和游離胚乳核;2.授粉后3 d合子分裂形成原胚;3.正在分裂的原胚細胞和胚乳核;4.授粉后5 d,原胚細胞分裂形成球形胚;5.授粉后12 d,球形胚體積增大;6.授粉后12 d,棒狀形胚;7. 授粉后12 d,心形胚;8.授粉后15 d,魚雷形胚;9. 授粉后15 d,子葉形胚;10.授粉后2 d,無合子只有游離胚乳核;11.授粉后3 d,合子未分裂;12.授粉后3 d,兩極核未融合;13.授粉后3 d,無胚發育的胚囊;14.授粉后5 d,敗育的球形胚;15.授粉后12 d,正在解體的球形胚。圖版Ⅱ ‘雜交石竹’胚胎發育情況(標尺=200 μm)Zy. Zygote; EFN. Endosperm free nuclei; PN. Polar nucleiFig.1. Formation of a zygote and endosperm free nuclei 2 days after pollination; Fig.2. Zygote division formed proembryon 2 days after pollination; Fig.3. Division of proembryo and endosperm free nuclei 2 days after pollination; Fig. 4. Proembryo division formed globular embryo 5 days after pollination; Fig.5. Globular embryo volume increase 12 days after pollination; Fig.6. Club-shaped embryo 12 days after pollination; Fig. 7. Heart embryo 12 days after pollination; Fig.8. Torpedo embryo 15 days after pollination; Fig.9. Cotyledonary embryo 15 days after pollination; Fig.10. Only the endosperm without embryo sac 2 days after pollination; Fig.11. Undivision of zygote 3 days after pollination; Fig.12. Degenerated endosperm free nuclei 3 days after pollination; Fig.13. No embryos of embryo sac 3 days after pollination; Fig.14. Degenerated globular embryo 5 days after pollination; Fig.15. The dissolution of globular embryo 12 days after pollination.Plate Ⅱ Embryogenesis after pollination in ‘Dianthus hybridus’(Bars=200 μm)
(編輯:潘新社)
Pollination,Fertilization and Embryonic Development of Dianthus hybridus
ZHANG Saiyang, LI Yan, HAO Pengbo, YANG Qiusheng*
(College of Forestry, Henan Agricultural University, Zhengzhou 450002,China)
With ‘Dianthushybridus’ as experiment material, we used fluorescence microscope to observe the pollen germination and pollen tube growth after pollination, to study the process of fertilization and embryo development with paraffin section method. The results showed that: (1) at 1 h after pollination, a small amount of pollens germinationed; At 4 h after pollination, a large number of pollens germinationed, pollen tube tips were found in the style middle, and callose response was found in pollen tubes; 6 h after pollination pollen tube grow to capital base and with a small amount of ovules; After 15 h stylar cannal has callose response, the pollen tube with the ovule number increased; At 24 h, many of pollen tubes have arisen around, one of the pollen tube enters the ovule, some pollen tubes have formed coiled overgrowth and generate callose reaction in the embryo sac. Sperm nucleus fused with polar nuclei mainly in 17-48 h after pollination, and the egg cell fusion process is mainly in 1-3 d after pollination. (2) The development of embryo through the proembryo, globular embryo, rod shaped embryo, heart-shaped embryo, torpedo embryo and cotyledon embryo stages. (3) Cross obstacle characterized by: only free endosperm nuclear and no embryo developmental embryo sac, zygote did not divided, sperm did not fused with polar nucleus, only free endosperm development without embryos of embryo sac, globular embryo aborted. The pre-fertilization and post-fertilization, obstacle are the main cause of low seed setting rate.
Dianthushybridus; pollen germination; pollen tube growth; fertilization; embryo abortion
1000-4025(2016)10-1984-06
10.7606/j.issn.1000-4025.2016.10.1984
2016-07-14;修改稿收到日期:2016-08-16
張賽陽(1990-),男,在讀研究生,主要從事園林植物種質資源創新及育種。E-mail:zhangsaiyang1990@126.com
*通信作者:楊秋生,博士生導師,主要從事園林植物栽培教學和研究。E-mail:qsyang@henau.edu.cn
Q321+.8; Q944.58
A
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
